
内生真菌对雷公藤组培苗生长、生理及药用成分含量的影响
2024-01-30 04:28胡清霞林照授吴承祯1c
植物资源与环境学报 2024年1期
吴 慧, 胡清霞, 宋 萍, 林照授, 封 磊, 吴承祯, 洪 伟,1c
(1. 福建农林大学: a. 林学院, b. 菌草与生态学院, c. 森林生态系统过程与经营福建省高校重点实验室, d. 资源与环境学院, 福建 福州 350002; 2. 福建省大田桃源国有林场, 福建 三明 366031; 3. 武夷学院, 福建 武夷山 354300)
雷公藤(TripterygiumwilfordiiHook. f.)为多年生木质藤本植物,是中国传统药用植物。从雷公藤植株中提取的雷公藤红素和雷公藤甲素是当前研究较多的药用活性成分,具有较高的临床应用价值,其中,雷公藤红素可用于治疗肥胖、慢性炎症和免疫性疾病[1-3];雷公藤甲素是一种高活性环氧化二萜内酯化合物,具有抗肿瘤、抗炎和免疫抑制等作用[4]。然而,这2种药用活性成分在雷公藤植株体内含量极低[5],加上雷公藤野生资源蕴藏量少,人工培育期长,限制了雷公藤及其药用活性成分在医药上的应用和发展。因此,采取有效措施改善雷公藤生长及促进雷公藤药用活性成分积累具有重要意义。
植物内生真菌是指生活史的部分或全部阶段在宿主植物内部,并且不会引起宿主植物明显病症或造成明显伤害的一类真菌。内生真菌能够促进植物生长。例如:Obledo等[6]从笹之雪(Agavevictoriae-reginaeT. Moore)体内分离出的非致病真菌能够促进笹之雪叶片和根部生长,提高其叶绿素含量,增强其光合能力;Mucciarelli等[7]认为,辣薄荷(Mentha×piperitaLinn.)内生真菌能够促进宿主植物生长发育,使植株高度和叶片大小明显提高。内生真菌能利用自身的代谢产物促进植物生长,增加植物产量。例如:印度梨形孢(Piriformosporaindica)可以分泌吲哚乙酸(IAA)等生长素改善植物生长[8];内生真菌Sebacinavermifera能增强渐狭叶烟草(NicotianaattenuataTorr. ex S. Watson)的生长活力,抑制渐狭叶烟草体内乙烯的产生[9]。内生真菌还能够增强植株对逆境的抗性。例如:印度梨形孢能提高作物的抗病性和耐盐性[10];Boutelouaeriopoda(Torr.) Torr.和四翼滨藜〔Atriplexcanescens(Pursh) Nutt.〕的内生真菌通过向根表面分泌多糖类黏液物质并形成稳定的菌膜,与植物协同抗旱[11];Paraphaeosphaeriaquadriseptata能分泌一种抑制植物热激蛋白HSP90活力的活性物质,从而增强植物的耐热能力[12];禾本科(Poaceae)植物与Epichlo⊇属内生真菌的共生体具有更强的生态适应性及竞争力[13];内生真菌能够改变干旱条件下水稻(OryzasativaLinn.)自身激素的积累,减弱因为缺水导致的光合作用中色素的分解和损失,从而加强水稻在干旱环境中的抵抗力[14]。内生真菌可以通过分泌抗生素、生态位竞争、重寄生作用及诱导植物抗性等提高植物的抗病能力[15]。如球孢白僵菌(Beauveriabassiana)分泌的白僵菌黄色素、白僵菌素、类白僵菌素和白僵内酯等代谢产物会对农林植物的病虫产生毒害作用[16]。植物内生真菌与宿主植物协同进化,其不仅能够产生与宿主植物相同或相似的生物活性物质,还能够调节植物体内某些生物活性物质的合成和积累。例如:内生真菌E4(Fusariiumsp.)和E5(Fusariiumsp.)不仅能提高大戟(EuphorbiapekinensisRupr.)单株根的鲜质量和折干率,而且能促进大戟中萜类活性物质异大戟素和大戟醇的积累[17];Ye等[18]发现,内生真菌Ilyonectrialiriodendra可促进蕺菜(HouttuyniacordataThunb.)根茎生长以及多数酚类物质和挥发物的积累,Penicilliumcitrinum可增加植株鲜质量、总叶面积和高度,促进根茎生长。由此可见,内生真菌在植物生长发育、胁迫抗性、病原体防御以及次生代谢产物积累中发挥着重要作用。然而,雷公藤内生真菌对宿主植物生长及药用成分积累的作用尚不明确。
针对雷公藤生长缓慢、药用活性成分含量低的问题,本研究在分析雷公藤内生真菌促生潜力的基础上,将内生真菌与雷公藤组培苗共培养,研究内生真菌对宿主植物生长、光合作用及营养元素吸收的影响,探究内生真菌对宿主植物积累雷公藤红素和雷公藤甲素的作用,以期为改善雷公藤生长及提高药用活性成分含量提供研究依据。
1 材料和方法
1.1 材料
供试材料为种植于福建农林大学森林生态系统过程与经营福建省高校重点实验室田间试验基地(东经119°14′08.65″、北纬26°05′06.55″)的人工栽培的3年生雷公藤植株,参照宋萍等[19]的方法,从雷公藤根、茎和叶片分离内生真菌。经与雷公藤组培苗共培养测试,将其中可与组培苗共生的11株菌株作为供试菌株,分别为菌株NS1、NS4、NS6、NS7、NS13、NS14、NS18、NS25、NS31、NS32和NS33。各菌株保藏于作者所在实验室。
1.2 方法
1.2.1 菌株促生潜力测定 参考Amprayn等[20]的方法测定菌株培养液中吲哚乙酸分泌量。每种菌株3次重复,每重复测定1次。
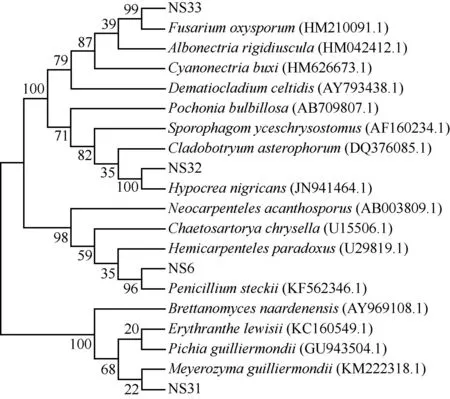
采用CAS比色法[21]测定菌株铁载体相对产量(SP),计算公式为SP=(1-A/Ar)×100%,式中,A为供试液波长630 nm处的吸光度,Ar为参比值,即空白检测液的吸光度。根据文献[22],当SP值分别在80% 将供试菌株接种于马铃薯葡萄糖琼脂(PDA)平板培养基活化培养,取活化的菌落接种于改进的Pikovskaya无机磷培养基〔含10.0 g·L-1葡萄糖、0.5 g·L-1(NH4)2SO4、0.3 g·L-1NaCl、0.3 g·L-1KCl、0.3 g·L-1MgSO4·7H2O、0.03 g·L-1FeSO4·7H2O、0.03 g·L-1MnSO4·4H2O、5.0 g·L-1Ca3(PO4)2和20.0 g·L-1琼脂,pH 7.0至pH 7.5〕中,以添加相同大小的无菌PDA培养基作为空白对照。每个处理3次重复,每重复测定1次。摇床震荡培养7 d后于4 ℃、8 000g离心10 min,采用钼锑抗比色法[23]测定上清液中的磷含量,上清液中的磷含量减去空白对照中的磷含量即为菌株的溶磷量。 1.2.2 内生真菌与组培苗共生体系建立 将内生真菌接种于PDA平板培养基上活化培养。在活化的菌落边缘用打孔器取直径5 mm的菌块,选取株高和叶片数等长势相近的雷公藤第3代无菌组培苗〔种植于添加质量浓度1.0 mg·L-12,4-二氯苯氧乙酸(2,4-D)、0.1 mg·L-1萘乙酸(NAA)和0.1 mg·L-1激动素(KT)的MS培养基,pH 5.8至pH 6.0〕,将菌块置于距组培苗根部2 cm处对峙培养,对照(CK)放置同样大小的PDA培养基。每个处理10瓶,每瓶1株组培苗。置于温度25 ℃、光照时间10 h·d-1、光照强度40 mol·m-2·s-1的光照培养箱中培养30 d,待测。 1.2.3 生长指标测量 随机取待测组培苗3株,测定组培苗植株的单株鲜质量、单株干质量、株高(茎基部至株顶的距离)、最长根长、单株叶片数。其中,株高和最长根长使用直尺(精度1 mm)测量;用超纯水洗掉组培苗根部粘附的培养基,表面水分用滤纸吸干,使用千分之一电子天秤称量单株鲜质量,然后在60 ℃烘箱中烘至恒质量,称量单株干质量。每个植株的各指标均测量1次。 1.2.4 叶绿素含量和叶绿素荧光参数测定 每株称取新鲜的雷公藤组培苗中部叶片0.2 g,每个处理重复3次,分别加入丙酮-乙醇(体积比1∶1)混合浸提液,定容至25 mL。待叶片变白后,以混合浸提液作为空白对照,使用UV-2000紫外可见分光光度计(上海尤尼柯仪器有限公司)分别测定波长663和645 nm处的吸光度A663和A645[24]。叶绿素含量的计算公式为叶绿素含量=(20.3A645+8.04A663)V/(1 000m),式中,V为浸提液体积,m为叶片鲜质量。 选择组培苗中部3枚充分伸展的叶片,暗适应30 min后使用OS-5P便携式脉冲调制叶绿素荧光仪(美国OPTI-Sciences公司)测定初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)和PSⅡ最大光化学效率(Fv/Fm)。 1.2.5 C、N、P和K含量的测定 将雷公藤组培苗整株在60 ℃烘箱中烘至恒质量,粉碎后过孔径0.22 mm筛。称取0.400 0 g,加入30 mL浓硝酸-浓高氯酸(体积比5∶1)混合液消煮。使用EA3000元素分析仪(意大利Euro Vector公司)测定C和N含量;采用钼锑抗比色法[23]测定P含量;使用FP-640型火焰光度计(上海精密科学仪器公司)测定K含量。重复测定3次。 1.2.6 雷公藤红素和雷公藤甲素含量的测定 参考文献[1],使用L-2000高效液相色谱仪(日本日立公司)测定雷公藤组培苗根、茎和叶片中的雷公藤红素和雷公藤甲素含量。重复测定3次。 1.2.7 菌种的分子鉴定 通过26SrDNA D1/D2区序列分析对所选菌株进行分子鉴定。首先,将少量活化菌体加入到50 μL TaKaRa Lysis Buffer for Microorganism to Direct PCR细胞裂解液(Code No. 9164)(日本TaKaRa公司)中,80 ℃变性15 min,于4 ℃、10 000g离心10 min分离出DNA。取1 μL上清液作为PCR反应模板,使用真菌鉴定试剂盒TaKaRa Fungi Identification PCR Kit(Code No. RR178)扩增目的片段。采用上述真菌鉴定试剂盒提供的测序引物,正向引物序列为5′-CGCCAGGGTTTT CCCAGTCACGAC-3′,反向引物序列为5′-GAGCGG ATAACAATTTCACACAGG-3′。PCR扩增体系总体积50.0 μL,包括1.0 μL DNA模板、25.0 μL PCR Premix、0.5 μL正向引物、0.5 μL反向引物和23.0 μL dH2O。PCR扩增程序:94 ℃预变性5 min;94 ℃变性0.5 min、55 ℃退火0.5 min、72 ℃延伸1 min,共30个循环;最后72 ℃延伸5 min。利用TaKaRa MiniBEST Agarose Gel DNA Extraction Kit Ver. 4.0试剂盒(Code No. 9762)(日本TaKaRa公司)对目的片段切胶回收,交由宝生物工程(大连)有限公司测序。将测序结果提交至NCBI数据库,获取GenBank登录号,通过BLASTn进行同源序列比对,采用邻接法构建系统发育树。 利用EXCEL 2010软件统计数据;利用SPSS 18.0软件进行单因素方差分析(one-way ANOVA),采用Duncan’s test进行多重比较。 雷公藤不同内生真菌菌株促生指标的比较结果见表1。由表1可以看出:雷公藤11株内生真菌菌株中,菌株NS32的吲哚乙酸分泌量最大,吲哚乙酸分泌量为33.10 mg·L-1,显著(P<0.05)高于其他菌株;菌株NS6、NS7和NS14的吲哚乙酸分泌量也较高,分别为17.24、30.76和21.78 mg·L-1,显著高于剩余7株菌株(吲哚乙酸分泌量小于6.00 mg·L-1)。产铁载体能力很高〔80%<铁载体相对产量(SP)≤100%〕的菌株为NS1、NS4、NS6、NS25和NS33,显著高于其他菌株;产铁载体能力较高(60% 表1 雷公藤不同内生真菌菌株促生指标的比较 2.2.1 对生长指标的影响 不同内生真菌菌株对雷公藤组培苗生长指标的影响见表2。由表2可以看出:菌株NS33、NS32和NS1处理的雷公藤组培苗的单株鲜质量和干质量均显著(P<0.05)高于对照(CK),其中,菌株NS33的促进作用最大,组培苗的单株鲜质量和干质量分别较对照增加了71.9%和76.4%。菌株NS6处理的组培苗单株鲜质量和菌株NS31处理的组培苗单株干质量也显著高于对照。菌株NS4、NS7、NS13和NS14处理的组培苗单株鲜质量以及菌株NS4、NS7、NS13和NS25处理的组培苗单株干质量较对照显著降低。菌株NS4、NS6、NS18、NS25、NS32和NS33处理的组培苗株高显著高于对照,其他5株菌株处理与对照无显著差异。菌株NS1、NS6、NS18和NS25处理的组培苗最长根长显著高于对照,其他7株菌株处理与对照无显著差异。仅菌株NS6处理的组培苗单株叶片数显著高于对照,其他10株菌株处理与对照无显著差异。 表2 不同内生真菌菌株对雷公藤组培苗生长指标的影响 2.2.2 对叶片叶绿素含量和叶绿素荧光参数的影响 不同内生真菌菌株对雷公藤组培苗叶片叶绿素含量和叶绿素荧光参数的影响见表3。由表3可以看出:除菌株NS13处理的雷公藤组培苗叶片叶绿素含量显著低于对照外,其他10株菌株处理的组培苗叶片叶绿素含量均显著高于对照,其中,菌株NS18处理的组培苗叶片叶绿素含量最高(2.738 mg·g-1),菌株NS6和NS25处理的组培苗叶片叶绿素含量也较高(分别为2.512和2.456 mg·g-1)。菌株NS7和NS13处理的组培苗叶片初始荧光(Fo)显著高于对照,其他9株菌株处理与对照无显著差异。菌株NS1、NS7、NS18、NS31和NS32处理的组培苗叶片最大荧光(Fm)显著高于对照,其他6株菌株处理与对照无显著差异。菌株NS1、NS18、NS31、NS32和NS33处理的组培苗叶片可变荧光(Fv)显著高于对照,其他6株菌株处理与对照无显著差异。菌株NS14处理的组培苗叶片PSⅡ最大光化学效率(Fv/Fm)显著高于对照,而菌株NS7和NS13处理的组培苗叶片Fv/Fm值则显著低于对照,其他8株菌株处理与对照无显著差异。 表3 不同内生真菌菌株对雷公藤组培苗叶片叶绿素含量和叶绿素荧光参数的影响 2.2.3 对C、N、P和K含量的影响 不同内生真菌菌株对雷公藤组培苗中C、N、P和K含量的影响见表4。由表4可以看出:除菌株NS13、NS18和NS25处理的雷公藤组培苗C含量显著低于对照外,其他8株菌株处理的组培苗C含量显著高于对照,其中,菌株NS14处理的组培苗C含量最高,菌株NS1、NS7和NS31处理的组培苗C含量也较高,较对照升高了15.5%~21.5%。菌株NS1、NS4、NS32和NS33处理的组培苗N含量显著高于对照,较对照升高了29.0%~137.2%,其中,菌株NS33处理对组培苗N含量的促进作用最大,其次为菌株NS1处理;其他7株菌株处理的组培苗N含量接近或显著低于对照。与对照相比,11株菌株处理的组培苗P含量均有所升高,其中,NS1、NS6、NS7、NS13、NS14、NS25、NS31、NS32和NS33菌株处理的组培苗P含量显著高于对照。菌株NS1、NS7、NS14、NS18、NS31、NS32和NS33处理的组培苗K含量显著高于对照,其他4株菌株处理的组培苗K含量则接近或显著低于对照。 表4 不同内生真菌菌株对雷公藤组培苗中C、N、P和K含量的影响 不同内生真菌菌株对雷公藤组培苗中雷公藤红素和雷公藤甲素含量的影响见表5。由表5可以看出:11株菌株处理的雷公藤组培苗根中雷公藤红素含量显著(P<0.05)高于对照,较对照升高了3.3~10.3倍,其中,NS6、NS14和NS33处理的组培苗根中雷公藤红素含量显著高于其他菌株处理;菌株NS4、NS7和NS31处理的组培苗根中雷公藤红素含量也较高,显著高于剩余5株菌株处理。11株菌株处理的雷公藤组培苗茎中雷公藤红素含量显著高于对照,较对照升高了48.7%~104.2%,其中,菌株NS7、NS18和NS32处理的组培苗茎中雷公藤红素含量显著高于其他菌株处理,菌株NS1和NS13处理的组培苗茎中雷公藤红素含量也较高。内生真菌对组培苗叶片中雷公藤红素含量的影响总体较小,仅菌株NS33处理的组培苗叶片中雷公藤红素含量显著高于对照,其他10株菌株处理与对照无显著差异。 表5 不同内生真菌菌株对雷公藤组培苗中雷公藤红素和雷公藤甲素含量的影响 雷公藤组培苗茎和叶片中均未检出雷公藤甲素。菌株NS6、NS7、NS13、NS25、NS32和NS33处理的组培苗根中雷公藤甲素含量显著高于对照,其中,菌株NS7处理的组培苗根中雷公藤甲素含量最高,显著高于其他菌株处理;菌株NS32和NS33处理的组培苗根中雷公藤甲素含量也较高;菌株NS1、NS14、NS18和NS31处理的组培苗根中雷公藤甲素含量显著低于对照。 综合上述研究结果,菌株NS1、NS6、NS31、NS32和NS33具有促生潜力且能够明显促进雷公藤组培苗生长、营养元素积累,提高雷公藤红素和雷公藤甲素含量,对这5株内生真菌进行分子鉴定。菌株NS1为Fusariumnisikadoi[25],将其余4株菌株的26SrDNA D1/D2区基因序列在NCBI中进行BLASTn比对,并构建系统发育树,结果见图1。由图1可以看出:菌株NS6、NS31、NS32和NS33分别与歧皱青霉(Penicilliumsteckii)、季也蒙迈耶氏酵母(Meyerozymaguilliermondii)、黑肉座菌(Hypocreanigricans)和尖孢镰刀菌(Fusariumoxysporum)聚在同一分支上,BLASTn比对序列相似性均达99%。推测菌株NS6、NS31、NS32和NS33分别属于青霉菌属(Penicillium)、迈耶氏酵母属(Meyerozyma)、肉座菌属(Hypocrea)和镰刀菌属(Fusarium)。将菌株NS6、NS31、NS32和NS33的序列提交GenBank,登录号分别为KP715295、KP715296、KP715297和KP715298。 分支上的数值代表各分支的频率参数The values on the branches indicate the frequency parameters of each branch. 括号中编号为GenBank登录号Nos. in brackets are GenBank accession numbers. 内生真菌对宿主植物的生长发育具有重要的促进作用。植物促生真菌被广泛应用于农林生产中,在提高产量、防治病害和增强胁迫耐性方面有重要作用[26]。植物内生真菌对植物促生效应存在表型差异,例如:内生真菌Gilmaniellasp. AL12能够将苍术〔Atractylodeslancea(Thunb.) DC.〕苗的茎和根质量提高1.13倍以上[27];内生真菌AlternariaA7显著增加了烟草(NicotianatabacumLinn.)的叶面积和干质量[28];而深色有隔内生真菌链格孢菌Alternariasp. CGMCC 17463既增加了紫花苜蓿(MedicagosativaLinn.)的株高和地上部干质量,也提高了其根部干质量和根冠比[29]。与雷公藤组培苗共生培养后,菌株NS1、NS6、NS31、NS32和NS33总体上显著(P<0.05)促进了组培苗的单株鲜质量和干质量,菌株NS6、NS32和NS33对株高有显著促进作用,菌株NS1和NS6对最长根长有显著促进作用,菌株NS6能显著增加单株叶片数,说明雷公藤植株内存在能促进其生长的真菌,但这些促生内生真菌对其最长根长和株高等表型的影响存在差异。内生真菌促生效应的植物表型差异可能源于不同内生真菌促生机制的差异。 内生真菌具有生物固氮、磷溶解、产生植物激素和拮抗病原菌等特性,对宿主植物的生长发育有促进作用[30]。如球状茎点霉菌(Phomaglomerata)和青霉菌(Penicilliumsp.)能够分泌赤霉素和吲哚乙酸,从而显著促进水稻生长[31];具有溶磷能力的内生百岁兰曲霉(Aspergilluswelwitschiae)能够增加大豆〔Glycinemax(Linn.) Merr.〕的根芽长、鲜质量和干质量[32]。雷公藤促生内生真菌中,菌株NS1具有较好的产铁载体和溶磷能力,菌株NS6的产铁载体、溶磷和吲哚乙酸分泌能力均较强,菌株NS31、NS32和NS33分别具有较高的溶磷、吲哚乙酸分泌和产铁载体能力。铁载体的生产有利于微生物和植物捕获环境中的铁离子。本研究中,推测属于镰力菌属的菌株NS1和NS33均具有很高的产铁载体能力,其他研究也报道了能产铁载体的镰刀菌,如纹瓣兰〔Cymbidiumaloifolium(Linn.) Sw〕的内生尖孢镰刀菌CAF1[33-34],且非致病镰刀菌比致病镰刀菌能生产更多的铁载体[35]。磷酸盐溶解微生物有助于提高植物根系对磷的吸收。具有溶磷能力的季也蒙迈耶氏酵母CC1能提高玉米(ZeamaysLinn.)产量,降低化学肥料的使用量[36]。吲哚乙酸是根和茎生长的重要植物激素[37]。相关研究结果显示:许多青霉菌能够分泌吲哚乙酸,如内生青霉菌LWL3能够促进水稻茎长、茎鲜质量、茎干质量和叶绿素含量,具有较高的吲哚乙酸分泌能力[31];定植于丹参(SalviamiltiorrhizaBunge)根部细胞间隙的歧皱青霉DF33能产吲哚乙酸[38]。因此,不同雷公藤促生内生真菌具有不同的促生潜力,对宿主组培苗生长的促进效应存在差异。 植物叶绿素含量越高,光合反应速率越快。叶绿素荧光参数是反映叶绿体状态和表征植物光合能量转换效率的重要参数。本研究中,除菌株NS13外,其他10株内生真菌均能显著提高雷公藤组培苗叶片叶绿素含量;菌株NS1、NS18、NS31和NS32能显著提高组培苗叶片的最大荧光和可变荧光,菌株NS14能显著提高组培苗叶片PSⅡ最大光化学效率,这些雷公藤内生真菌对宿主植物的PSⅡ电子传递有积极作用,能提高雷公藤光合能力。木麻黄(CasuarinaequisetifoliaLinn.)内生真菌可以提高其水培苗叶绿素相对含量及主要荧光参数[39];与深色有隔内生真菌相关的植物的叶绿素含量和PSⅡ光化学效率较高[40]。睡茄〔Withaniasomnifera(Linn.) Dunal〕根和叶片中部分内生真菌可以提高睡茄的光合效率以及茎和根的干质量[41]。上述研究结果说明内生真菌能够提高植物的叶绿素含量和叶绿素荧光参数值,提高植物的光合能力,光合作用同化物和糖类的产量也随之增加,植物的生长和生物量得到改善。因而,雷公藤内生真菌对组培苗叶绿素含量和叶绿素荧光参数值的提高作用可能是其促进组培苗生长的原因之一。 内生真菌可能增加植物养分吸收,进而促进植物生长[42]。在缺氮和正常供氮条件下接种内生真菌Suillusgranulatus、Pisolithustinctorius、Pleotrichocladiumopacum或Pseudopyrenochaetasp.能显著提高油松(PinustabuliformisCarriere)苗的生长性能,增加地上部和根部养分含量[42]。来自冬青(IlexchinensisSims)的深色有隔内生真菌Acrocalymmavagum和Scytalidiumlignicola可以明显提高镉胁迫下紫花苜蓿和沙冬青〔Ammopiptanthusmongolicus(Maxim. ex Kom.) S. H. Cheng〕的干质量,而二者分别对沙冬青的总氮含量和紫花苜蓿的有机碳含量有显著促进作用[43]。此外,深色有隔内生真菌还可以提高植物根和茎的质量、总质量以及N和P含量[44]。本研究进一步证实了促生内生真菌在植物养分获取中的积极作用,雷公藤促生内生真菌菌株NS1、NS32和NS33能显著提高组培苗中C、N、P和K含量,菌株NS6能显著提高组培苗中C和P含量,菌株NS31能显著提高组培苗中C、P和K含量。 内生真菌可以调节宿主植物基因表达和生理反应,诱导宿主植物的次生代谢。雷公藤红素和雷公藤甲素是雷公藤植株中的重要药用活性成分,11株雷公藤内生真菌对组培苗根和茎中雷公藤红素含量均有显著促进作用,分别较对照升高了3.3~10.3倍和48.7%~104.2%;对根中雷公藤甲素含量的影响则存在差异,其中,菌株NS6、NS7、NS13、NS25、NS32和NS33有显著促进作用。内生真菌与宿主植物相互作用的类型决定了宿主植物中次生代谢产物的变化。镰刀菌因其具有许多独特的基因簇参与次级代谢产物产生而被广泛关注[45]。勇应辉等[17]认为内生镰刀菌在促进大戟生长和萜类化合物含量中有重要作用。雷公藤内生真菌中,推测属于镰刀菌属的菌株NS1提高了根和茎中雷公藤红素含量,菌株NS33提高了根、茎和叶片中雷公藤红素含量以及根中雷公藤甲素含量。青霉菌属真菌因其具有高度通用的细胞毒性次生代谢产物合成潜力而得到广泛开发[46]。从丹参中分离的歧皱青霉DF33通过上调关键基因显著促进丹参根部丹参酮ⅡA和隐丹参酮的合成[38]。肉座菌属包含多种能够生产工业酶、次生代谢产物或生物防治剂的菌株[47]。来自野生丹参的黑肉座菌菌株S79显著促进丹参酮合成[48]。雷公藤内生真菌中,推测属于青霉菌属的菌株NS6和属于肉座菌属的菌株NS32均能显著提高雷公藤组培苗根和茎中雷公藤红素含量以及根中雷公藤甲素含量,进一步表明内生青霉菌和肉座菌对宿主植物次生代谢产物积累有促进作用,是促进植物中生物活性成分积累的重要功能真菌。内生真菌可能通过产生与宿主植物相同或相似的次生代谢产物而提高其在植物体内的积累,而某些内生真菌可以产生多种类型的酶或代谢前体物,这些酶或代谢前体物可以转化或诱导植物合成某些生物活性物质[49]。雷公藤内生真菌菌株NS1具有产雷公藤甲素的能力[25],然而与对照相比,与菌株NS1共培养的组培苗根中雷公藤甲素含量却显著降低,可能是内生真菌产物、共培养条件或植物的生长阶段等影响了植物的次生代谢[50]。 综上所述,雷公藤内生真菌能够分泌吲哚乙酸、产铁载体和溶解无机磷,总体能够促进雷公藤组培苗生长,增加叶绿素含量,改善养分吸收。部分内生真菌还参与雷公藤次生代谢产物的调控,促进植株体内雷公藤红素和雷公藤甲素的积累。本研究结果可为使用雷公藤内生真菌改善宿主植物生长和调控次生代谢提供科学依据。1.3 数据处理
2 结果和分析
2.1 雷公藤内生真菌的促生潜力

2.2 内生真菌对雷公藤组培苗生长和生理的影响
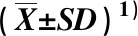
2.3 内生真菌对雷公藤组培苗中雷公藤红素和雷公藤甲素含量的影响
2.4 菌株的鉴定
3 讨论和结论
猜你喜欢
世界科学技术-中医药现代化(2021年7期)2021-11-04诗潮(2018年5期)2018-08-20现代园艺(2018年3期)2018-02-10江苏农业科学(2016年8期)2017-02-15现代园艺(2016年9期)2016-07-16西南农业学报(2016年6期)2016-04-16中国病理生理杂志(2015年8期)2015-12-21南方农业·下旬(2015年10期)2015-11-20江苏农业科学(2015年8期)2015-09-10食品工业科技(2014年23期)2014-03-11
