
甜菊糖苷的生物学功能及其在畜禽生产中的应用研究进展
2024-03-05 12:13何振涛熊云霞
动物营养学报 2024年2期
何振涛 熊云霞 王 丽 朱 翠
(1.佛山科学技术学院,生命科学与工程学院,佛山 528225;2.广东省农业科学院动物科学研究所,猪禽种业全国重点实验室,农业农村部华南动物营养与饲料重点实验室,岭南现代农业科学与技术广东省实验室茂名分中心,广东省畜禽育种与营养研究重点实验室,广州 510640)
甜叶菊为菊科、甜叶菊属多年生草本植物,被认为是世界第三大甜味来源。目前,我国的甜叶菊种植、甜菊糖苷生产和甜菊糖苷出口均居世界首位[1]。甜菊糖苷是一种从甜叶菊叶中提取的天然甜味剂。欧洲食品安全局(EFSA)和食品法典委员会(CODEX)均于2011年批准将甜菊糖苷用作食品添加剂。甜菊糖苷是一种天然、安全、低热量、高甜度的甜味剂,在全球食品、医药、化妆品和家用化工产品中具有广泛的应用[2]。根据国际市场分析研究和咨询集团(IMARC)的数据,甜叶菊的全球市场价值预计到2024年将达到近8.18亿美元[3]。研究发现,甜菊糖苷除了作为甜味剂使用外,还具有抗氧化、抗炎、改善血糖血压、抗菌、抗肿瘤、抗腹泻、利尿和免疫调节等多种生理功能[4]。近年来研究表明,在畜禽饲粮中添加适宜的甜菊糖苷有助于改善畜禽的生产性能[5-6]、提高饲料利用率[7-8]和调节肠道微生物群[9],表明甜菊糖苷在畜禽生产应用上具有巨大潜能。因此,本文主要综述了甜菊糖苷的消化、代谢、生物学功能,旨在为甜菊糖苷在畜禽生产中的合理开发和应用提供参考。
1 甜菊糖苷的化学结构、提取方法以及味觉评价
甜菊糖苷(C38H60O18)是甜叶菊的主要甜味成分,呈白色粉末状,是一类四环二萜类糖苷物质[10],主要分布于甜叶菊的根、茎、叶,其中以叶片含量最高(可高达14%)。研究表明,甜菊糖苷在各种加工和储存条件下均表现出良好的稳定性,不与水溶性维生素、有机酸、甜味剂和咖啡酸发生反应;热稳定性强,不参与美拉德反应;且长时间储存不发生发酵[11]。甜菊糖苷具有吸湿性强、低热、高甜、稳定性好且无毒副作用等优点,其甜度是蔗糖的50~300倍,可作为蔗糖的替代品[1,12]。现已从甜叶菊叶片中鉴定出64种甜菊糖苷,其中甜菊苷、莱鲍迪苷A、莱鲍迪苷B、莱鲍迪苷C、莱鲍迪苷D、莱鲍迪苷E、莱鲍迪苷F、甜菊双糖苷、杜克苷A、甜茶苷和莱鲍迪苷M等11种含量相对较高[13]。不同类型的甜菊糖苷化学结构主要差异在于R1和R2基团的区别(表1),其R1和R2基团分别连接在C13-羟基和C19-羧基[14]。
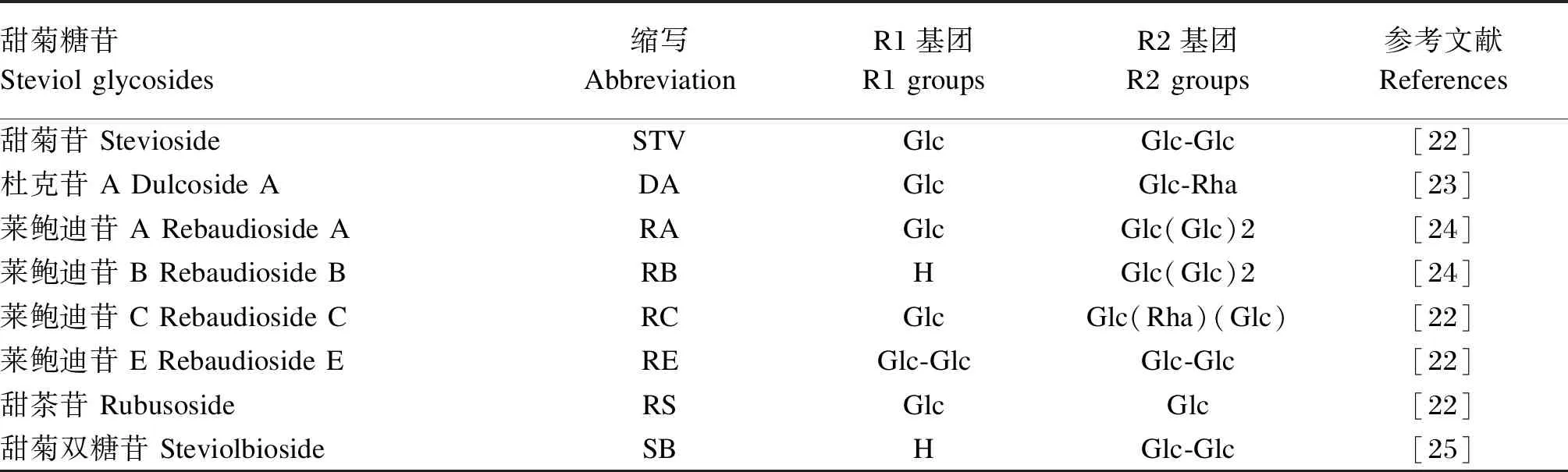
表1 甜菊糖苷的化学结构
甜菊糖苷作为新型代糖,可通过味觉感受器感知甜味和些许苦味等杂味。研究发现,甜菊糖苷可激活人体1型味觉受体成员2(T1R2)和1型味觉受体成员3(T1R3)感受甜味,也可激活2型味觉受体成员4(T2R4)和2型味觉受体成员14(T2R14)感受苦味[26]。甜菊糖苷的味觉感受与糖元链长度、吡喃糖取代和C16双键等结构特征有关,碳骨架中C-19上的葡萄糖基越少,苦味回味时间越长,而C-13上的葡萄糖基越多,起甜越快,甜度强度越强。例如,相较于莱鲍迪甙A和莱鲍迪甙C的甜味较弱和苦味较强,莱鲍迪苷M和莱鲍迪苷D虽然在甜叶菊中的天然含量很低,但具有很强的甜味和比较弱的苦味;而甜茶苷和甜菊苷则苦味后味强,甜味较弱[27]。研究表明,消除或减少甜菊糖苷苦味的最直接方法是在产品中添加风味增强剂或味觉修饰剂以掩盖其苦味,如赤藓糖醇、麦芽糖醇和木糖醇等[28],也可采用以麦芽糊精和菊粉为胶囊剂的喷雾干燥技术,可以最大限度地减少甜菊糖苷的苦味[29],或者通过酶促反应加强其亲水性从而掩盖其苦味[30]。研究发现,将Kamiya-8基团直接修饰甜菊糖苷骨架结构,也可提高甜菊糖苷对甜味受体T1R2及T1R3的亲和性,降低对苦味受体T2R4及T2R14的亲和性,从而改善甜菊糖苷的适口性[3]。
2 甜菊糖苷在机体中的水解代谢过程
目前研究表明,人和鼠对甜菊糖苷的水解代谢部位和代谢过程不尽相同。无论是人体还是大鼠体内(图1),甜菊糖苷通过口服后进入消化道,由于胃液及消化酶无法将其降解,导致其在肠道内无法被正常水解吸收[31-32]。研究表明,结肠菌群参与了甜菊糖苷的水解、吸收过程,拟杆菌属可以将甜菊糖苷水解为甜菊醇[32-34]。甜菊糖苷进入机体后首先被肠道微生物分解为甜菊醇,甜菊醇则经肝-肠循环在胆汁和肾脏中进一步代谢,进入肾脏后甜菊醇会在细胞色素P450酶的作用下代谢生成一羟基和二羟基代谢物[32]。研究证实,大鼠口服甜菊醇15 min后其血液中的甜菊醇浓度出现峰值,进一步用Caco-2细胞模型研究发现,肠道上皮细胞对甜菊醇的吸收转运很快,这与大鼠体模型结果[34]一致。此外,连续口服3 d甜菊糖苷的人粪便中未检测出甜菊糖苷,仅可检测出甜菊醇[35-36]。
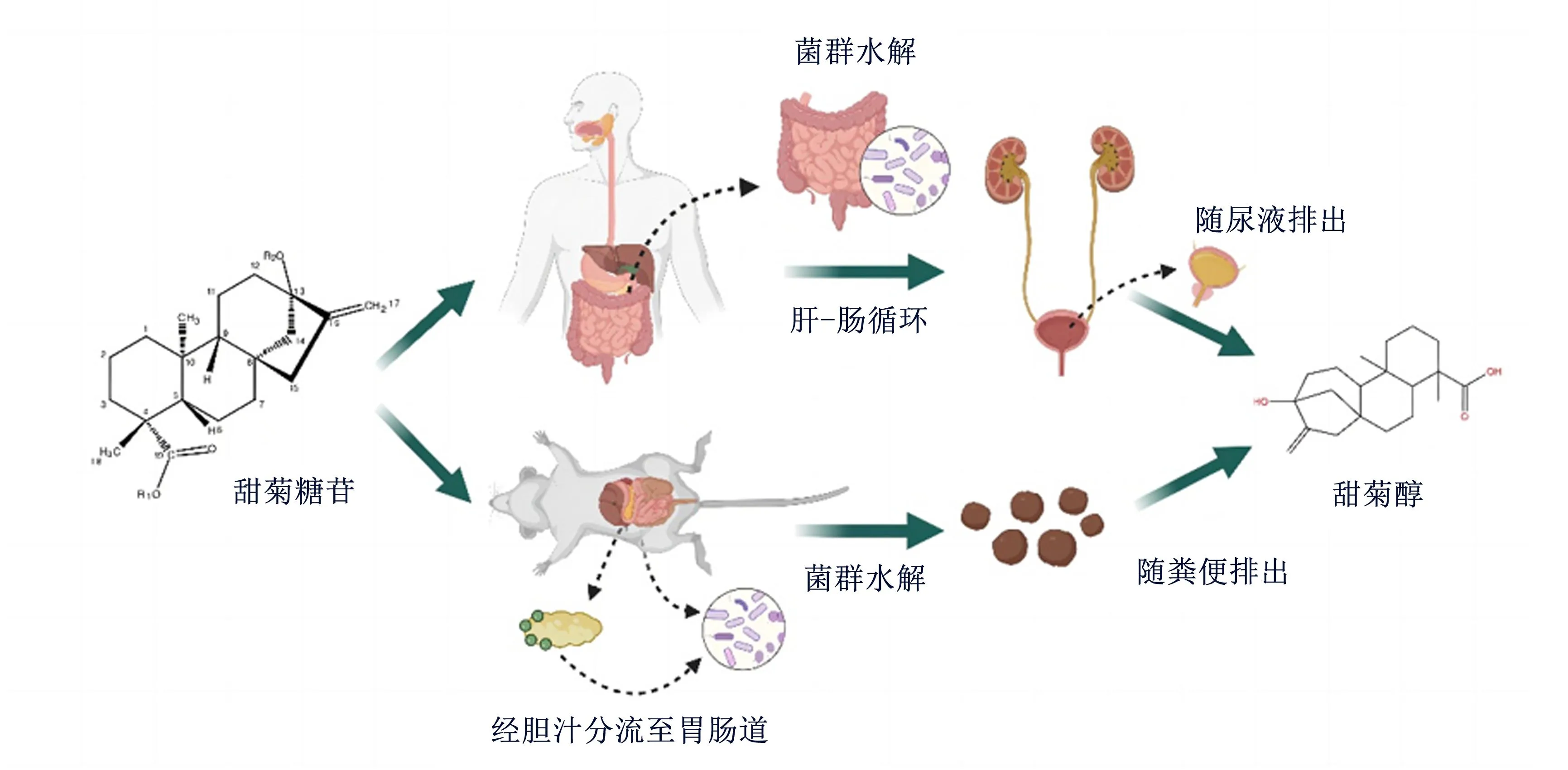
图1 甜菊糖苷的代谢过程
Cardoso等[37]将同位素131I标记的甜菊糖苷静脉注射Wistar雄性大鼠,研究其在体内的分布和代谢情况,发现注射10和120 min后,大鼠肝脏和小肠中的放射性物质浓度最高。由于消化系统的不同,甜菊糖苷在不同动物机体内代谢的过程也有所不同,可能涉及到肾脏的有机阴离子转移[38]。研究指出,在大鼠体内甜菊糖苷的主要代谢产物会通过胆汁随粪便排泄[39]。甜菊糖苷主要通过人体的肾脏代谢,然后随尿液排出,72 h后,尿液中会发现62%的甜菊糖苷以甜菊醇葡萄糖醛酸苷的形式存在[35-36,40],有机阴离子排泄量的差异与此有关。甜菊醇葡萄糖醛酸的分子质量为494 u,分子质量在600 u或以下的有机阴离子在人体中主要通过肾脏排泄,而分子质量在600 u或以上的有机阴离子在人体则主要通过胆汁排泄[41];相反,分子质量为325 u或更大的化合物在大鼠体内主要通过胆汁排出体外[41]。然而,目前关于甜菊糖苷在畜禽体内的代谢规律研究相对较少。
3 甜菊糖苷的生物学功能
3.1 抗氧化作用
研究证实,甜菊糖苷具有较强的抗氧化活性。甜菊糖苷对糖尿病大鼠肝脏中的超氧化物歧化酶(SOD)和过氧化氢酶(CAT)的活性都有明显提高,这对氧化损伤具有显著的保护作用[42]。甜菊糖苷可通过抑制细胞外调节蛋白激酶1/2(ERK1/2)、信号传导及转录激活因子3(STAT3)和核因子-κB(NF-κB)从而抑制小鼠肾脏的氧化应激,减少各种次级反应中产生的自由基的毒性作用[43]。体外研究也证实了甜叶菊及其提取物具有良好的抗氧化活性。Jahan等[44]测得甜叶菊80%乙醇提取物的最大吸光度为1.071 nm,而抗坏血酸作为标准物质的最大吸光度为1.374 nm,这表明甜叶菊乙醇提取物比抗坏血酸(维生素C)具有更高的还原能力。甜叶菊叶的水提取物对超氧阴离子、羟基自由基和2,2-二苯基-1-苦味酸(DPPH)具有有效的清除能力[45]。研究还发现,用甜叶菊叶处理四氧嘧啶诱导的糖尿病小鼠,能显著提升谷胱甘肽过氧化物酶(GSH-Px)活性和还原型谷胱甘肽(GSH)含量[46]。此外,添加200 mg/kg甜菊糖残渣提取物可显著改善高脂肪高果糖(HFFD)诱导的小鼠的抗氧化状态和肝脏脂肪积累情况[47],而添加200和400 mg/kg甜菊糖残渣提取物还能改善高尿酸血症小鼠的肠道结构和肠道抗氧化状态[48]。研究指出,甜叶菊提取物良好的抗氧化活性使其用作鲑鱼酱和其他海鲜产品的防腐剂成为可能[49]。
3.2 抗炎作用
研究表明,甜叶菊有机溶剂提取物具有较好的抗炎作用。例如,甜叶菊叶乙醇提取物可有效缓解花生四烯酸(AA)和佛波醇12-肉豆蔻酸13醋酸盐(TPA)诱导的炎症模型CF-1小鼠的耳水肿炎症[50]。对卡拉胶诱导的大鼠脚部肿胀,甜叶菊的氯仿提取物和甲醇提取物均能产生显著的抑制作用[51]。甜菊糖苷能够通过下调Toll样受体2(TLR2)、NF-κB和丝裂原活化蛋白激酶(MAPK)途径,来降低乳腺炎模型小鼠中肿瘤坏死因子-α(TNF-α)、白细胞介素-1β(IL-1β)和白细胞介素-6(IL-6)等促炎因子的水平[52]。甜菊糖苷还可以显著抑制结肠炎模型的小鼠的TNF-α和IL-6的水平和抑制环氧合酶-2(COX-2)和诱导型一氧化氮合酶(iNOS)蛋白的表达[53]。同样地,有研究发现,甜菊糖苷可以下调脂多糖诱导的巨噬细胞中IL-6、TNF-α、IL-1β、COX-2、高迁移率族蛋白B1(HMGB1)和诱导型一氧化氮合酶/一氧化氮(iNOS/NO)等促炎因子的表达,并上调抗炎因子转化生长因子-β1(TGF-β1)和白细胞介素-10(IL-10)的表达[54]。还有研究发现,甜菊糖苷和甜菊醇对Caco-2细胞无毒,并能阻断核因子抑制蛋白α/核因子-κB(IκBα/NF-κB)信号通路,该通路在脂多糖作用下会导致TNF-α、IL-1β和IL-6的产生[55]。
3.3 降糖、降压作用
甜菊糖苷作为低卡、高甜度代糖,可改善机体血糖状况。Shivanna等[42]的研究表明,甜菊叶粉可增强Ⅰ型糖尿病大鼠胰岛β细胞的胰岛素分泌,Ⅱ型糖尿病大鼠的细胞胰岛素敏感性和葡萄糖耐受性也可提高。此外,研究还发现,甜菊糖苷能显著改善糖尿病大鼠的胰岛素和肝糖原水平,同时显著降低其空腹血糖和糖化血红蛋白的水平[56-57]。当机体血糖水平较高时,胰岛素分泌和莱鲍迪苷A之间存在直接的剂量-反应关系[58]。甜菊糖苷可以通过激活钙离子(Ca2+)从而激活味觉受体与胰腺β细胞上的阳离子通道(TRPM5)来增强葡萄糖诱导的胰岛素分泌,进而改善机体血糖情况[59-60]。有研究发现,甜菊醇能够显著减少肠道中的葡萄糖积累,这是由于甜菊醇会降低肠道黏膜中三磷酸腺苷(ATP)含量和改变肠道形态结构,从而抑制肠道对葡萄糖的吸收[61]。
有研究称,女性高血压患者服用500 mg/d甜菊糖苷2年后,明显降低了收缩压和舒张压[62]。Chan等[63]也发现28~75岁的女性高血压患者服用0.25 g甜菊糖苷3次/d,仅7 d后,收缩压和舒张压均显著降低。研究认为,甜菊糖苷良好的降血压作用可能与其强烈的抑制血管紧张素转换酶活性有关[64]。
3.4 抑菌、抗癌作用
Puri等[65]发现,通过乙醇、甲醇和氯仿等有机溶剂进行提取纯化的甜菊糖苷,可抑制肺炎克雷伯菌、蜡样芽孢杆菌、铜绿假单胞菌、枯草芽孢杆菌、脱氮产碱杆菌和鼠伤寒沙门氏菌等的生长,特别是对枯草芽孢杆菌、肺炎克雷格菌和铜绿假单胞菌的抗菌活性呈现明显正向的剂量依赖性变化。有意思的是,成年人用莱鲍迪苷漱口后的牙菌斑累积量比用蔗糖漱口后的牙菌斑累积量降低82%[66],而甜菊糖苷因其具有抑菌能力被广泛应用于口腔溃疡和牙龈疾病的传统治疗[67]。
通过研究发现,甜菊糖苷还可通过选择性抑制膀胱癌细胞的增殖能力,并通过诱导细胞凋亡从而抑制其生长[68]。甜菊糖苷还能降低乳腺癌细胞MDA-MB-231细胞的迁移和转移潜能[69],降低其对机体的侵袭性,且甜菊糖苷的主要成分甜菊醇对MCF-7乳腺癌细胞具有较强的细胞毒性作用,可以通过G2/M周期阻滞抑制MCF-7细胞的增殖[70]。Chen等[71]研究表明,甜菊醇对人胃MKN-45、MGC-803和HCG-27以及结直肠HCT116、HCT-8和Caco-2等6种癌细胞系均有抗增殖作用,对细胞活力的抑制呈剂量依赖性,且胃癌细胞比结直肠癌细胞更敏感;进一步分析细胞周期证实甜菊醇可使MKN-45、HGC-27、HCT-116和Caco-2细胞在G1期阻滞,而使MGC-803和HCT-8细胞在G2期阻滞。
4 甜菊糖苷在动物生产中的应用
4.1 甜菊糖苷在家畜生产中的应用
研究发现,对猪、牛、羊等味觉敏感家畜而言,甜菊糖苷可影响其生产性能,提高日增重,并有助于降低幼龄家畜的腹泻[5,72]。研究表明,在断奶后第2周,饲粮中添加167 mg/kg甜菊糖苷可显著提高断奶仔猪的日增重,但是添加不同比例(0.083 3%、0.167%或0.334%)的甜叶菊苷对断奶仔猪采食量和料重比无显著影响[72]。在饲粮中添加300 mg/kg甜菊糖苷时,28日龄断奶仔猪的平均日增重最高,平均日采食量呈线性增加,料重比则呈线性下降,且当添加251 mg/kg甜菊糖苷时断奶仔猪腹泻率最低[5]。此外,在饲粮中添加0.3%甜菊糖苷,还能显著提高杜长大生长猪的日增重与饲料利用率,增强机体免疫力,改善其背膘厚度等胴体性状[7]。总的来说,在饲粮中添加甜菊糖苷可能通过提高饲料利用率和增强机体免疫力,从而有助于改善猪的生长性能。目前,关于甜菊糖苷如何影响家畜动物的肠道菌群变化研究还比较少,且甜菊糖苷对猪只生长性能的改善是否与肠道菌群的调控有关还有待进一步研究揭示。但有研究证实,在妊娠母猪饲粮中添加甜叶菊残渣(含有甜菊糖苷、绿原酸类、黄酮类等物质)能调控肠道菌群的组成,显著促进肠道有益菌(如毛螺菌科和瘤胃球菌科)的相对丰度,并减少密螺旋体等有害菌的相对丰度,并且可以显著提高哺乳母猪平均日采食量[73]。
目前,甜菊糖苷作为饲料添加剂应用于牛、羊饲粮的研究相对较少。Han等[74]发现,在山东黑山羊饲粮中添加400~800 mg/kg甜菊糖苷可以显著提高采食干饲草的量和饲粮总采食量。在饲粮中添加0.3%甜菊糖可以提高韩牛的末体重、增重和胴体的粗蛋白质含量,并显著降低了滴水损失和剪切力,增加了背最长肌的肉色红度值,从而改善胴体品质[75]。然而,具体关于饲粮中添加甜菊糖苷调控牛、羊生产性能的作用机制还需要进一步进行研究。
4.2 甜菊糖苷在家禽生产中的应用
研究发现,甜菊糖苷具有改善家禽生产性能和免疫功能的作用。然而,禽类由于缺失T1R2甜味受体,对甜味感受并不敏感,因此禽类主要通过非T1R2依赖途径影响其生产性能[6]。例如,添加0.3%甜叶菊提取物,能显著提高3周龄的青脚麻鸡的日增重和饲料利用率,增加血清白蛋白含量,提高法氏囊、脾脏和胸腺等器官指数[8,76]。Jiang等[6]对罗斯308肉鸡的研究也发现,在饲粮中添加250 mg/kg甜菊糖苷能显著提高其体重、平均日增重和平均日采食量。最新研究表明,甜菊糖苷可能通过调节下丘脑神经活性配体-受体相互作用途径和改变肠道微生物组成促进肉鸡采食量的增加[9]。由于具有甜菊糖苷的抗氧化和抗炎作用,甜菊糖苷显著改善了肉鸡血清和空肠的抗氧化能力,可显著降低NF-κB在回肠黏膜中的mRNA表达量,且对空肠IL-1β和回肠γ-干扰素(IFN-γ)的mRNA表达量有降低趋势,并显著提高肉鸡核因子E2相关因子2(Nrf2)、CAT和超氧化物歧化酶1(SOD1)在空肠和回肠黏膜中的mRNA表达量[77]。此外,在饲粮中添加80 mg/kg的甜叶菊甜味剂(含有0.5%甜菊糖苷)可以显著增加科宝系肉鸡试验第42天后的体重和日增重[78]。以上相关结果表明,在家禽饲粮中添加甜菊糖苷可以改善家禽饲料利用率、机体抗氧化能力和免疫功能,并且调控肠道微生物的组成从而有助于提高家禽生长性能。但是,也有研究发现,添加667 mg/kg甜菊糖苷对肉仔鸡的采食量、体重增加及饲料转化率均无显著影响[79]。需要注意的是,添加过高剂量的甜菊糖苷(3 200 mg/kg)对爱拔益加肉鸡的平均日增重、平均日采食量、料重比和免疫器官指数均无显著影响,但是显著降低了其血糖水平,抑制葡萄糖的合成,并提高钙和磷的表观消化率[80]。以上不同研究报道结果存在一定差异,这可能跟动物品种、甜菊糖苷的有效活性及添加剂量、处理时间和饲养管理等因素不同有关。
饲粮添加甜菊糖苷还可能存在母子代际效应。Jiang等[81]研究发现,在肉种鸡的饲粮中添加250 mg/kg甜菊糖苷可改善其鸡胚胎发育以及改善后代肉鸡的肠道形态完整性和肠道免疫功能,且还发现甜菊糖苷还可以通过调节肠道微生物群进而减轻脂多糖对后代肉鸡造成的肠黏膜损伤[82]。此外,通过给蛋鸡饲喂添加1%甜叶菊提取物的饲粮不仅能提高日增重和饲料利用率,改善其胃肠功能,增强免疫力,还可以提升鸡蛋中蛋黄色泽和鸡蛋中人体必需氨基酸以及其蛋白质总量[83]。目前,甜菊糖苷作为饲料添加剂在肉鸡和蛋鸡上的应用研究报道较多,而关于甜菊糖苷在其他家禽上应用研究仍相对较少。
5 小结与展望
综上所述,甜菊糖苷作为一种天然甜味剂,具有良好的抗氧化、抗炎、改善血糖血压、抗菌、抗癌和免疫调节等生物学功能。研究表明,在饲粮中添加甜菊糖苷具有提高畜禽采食量和促进生长的作用,还能提高畜禽的抗氧化能力、调节机体和肠道免疫功能,并对畜禽肉品质和蛋品质等方面具有一定的改善作用。因此,甜菊糖苷作为新型饲料添加剂在畜禽生产中具有广阔的应用前景。但是,目前甜菊糖苷在畜禽生产的应用研究报道还相对较少,并且关于其在影响畜禽生产性能方面的研究结果不尽一致。因此,未来仍需要进一步深入开展有关甜菊糖苷的加工工艺和有效活性的含量、在动物饲粮中最适的添加配比以及改善畜禽生产和肠道健康的作用机制等方面研究。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
食品工业(2021年5期)2021-06-10
天然产物研究与开发(2018年6期)2018-07-09
中国甜菜糖业(2018年2期)2018-01-24
动物营养学报(2017年8期)2017-08-16
中国糖料(2016年1期)2016-12-01
天津科技大学学报(2016年1期)2016-02-28
食品界(2016年4期)2016-02-27
动物营养学报(2015年9期)2016-01-07
生物技术通报(2015年9期)2015-10-25
