
铜死亡诱导剂FDX1、硫辛酸与冠状动脉病变严重程度的相关性
2024-03-18 11:31丁洋洋张佳佳李金龙康品方张宁汝
南方医科大学学报 2024年2期
魏 婷,丁洋洋,张佳佳,李金龙,张 恒,康品方,2,张宁汝
蚌埠医科大学1第一附属医院心血管科,2心脑血管基础与临床重点实验室,安徽 蚌埠233000
动脉粥样硬化(AS)是泛指动脉壁增厚、失去弹性、硬化性的疾病,以脂质沉积和内膜纤维化为特征,所导致的冠状动脉粥样硬化性心脏病(CHD)、心肌梗死、脑梗死等心脑血管疾病严重威胁人类生命健康[1,2]。CHD的一级预防、早期发现和治疗一直是全球长期的科学研究目标,但其致病机制及风险因素多种多样且复杂,导致了对它的防治仍然十分困难[3]。血脂异常是AS和CHD的直接相关危险因素,深入研究AS的发病机制、潜在治疗靶点的同时,探究脂代谢异常的相关机制对AS防治同样具有重要意义。
铁氧还蛋白1(FDX1)是一种线粒体代谢相关的基因,属于铁硫(Fe-S)簇蛋白家族,广泛表达于与类固醇生成相关的组织如:肾上腺、性腺,与脂肪酸氧化和氨基酸等代谢密切相关[4,5]。当FDX1活性的丧失会破坏铁硫簇酶和细胞铁稳态的活性,可导致线粒体铁超负荷和胞质铁消耗[6,7]。但目前为止,FDX1是否参与心血管疾病的发生发展尚不完全清楚,有关FDX1是否参与动脉粥样硬化斑块的形成,国内外鲜有研究。
硫辛酸(LA)在临床上是常用的抗氧化剂,在酒精性肝炎、神经病变以及缺血再灌注损伤等疾病中发挥强大的抵御氧化应激方面作用[8]。最新的研究[9]表明,FDX1是诱导铜死亡的关键基因,它通过上游调节蛋白质脂酰化过程来调控细胞对铜离子的敏感性,而LA是蛋白脂酰化反应的主要原料,那么通过上调上游调节因子FDX1的表达来促进线粒体合成内源性LA,是否有可能成为防治AS的新靶点有待研究。
目前FDX1与LA水平与CHD的相关性仍未被充分证实,本研究拟通过检测有无冠脉病变及冠脉不同病变程度下FDX1、LA水平差异,以及高脂血症动物模型的建立,明确FDX1与LA水平与高脂血症所致动脉血管损伤的相关性,为研究CHD的新型治疗、预防策略提供参考依据。
1 资料和方法
1.1 研究对象及分组
选择2021年10月~2022年10月在蚌埠医科大学第一附属医院的可能患有冠心病的226例患者进行临床诊断,其诊断分型依照1999年WHO/WPR标准。所有入院患者接受心电图和冠状动脉造影(CAG)等检查明确病情,并在入院后完善心脏彩超及抽血化验,记录患者既往病史等基本资料。根据CAG中手术记录患者的冠状动脉左主干、左前降支、左旋支、右冠状动脉等病变情况,将179例冠脉有病变的患者按照冠状动脉狭窄最严重的局部血管病变情况分为:轻度狭窄组(a-CHD,狭窄程度≤49%,n=30)、中度狭窄组(b-CHD,狭窄程度50%~74%,n=30)和重度狭窄组(c-CHD,狭窄程度≥75%,n=30),其中有47例虽有胸痛症状但CAG检查无异常患者作为对照组。排除标准:既往有心脏瓣膜性病、严重肝肾及心血管功能不全、恶性肿瘤病史、风湿及免疫系统疾病、心房颤动等[10]。入选患者均签署知情同意书,研究方案获得本院伦理委员会批准(伦科批字[2023]344号)。根据患者CAG结果计算Gensini评分,Gensini评分法[11](表1)。
1.2 一般资料收集及标本采集
收集所有入选患者的临床数据资料,记录患者基本信息、饮酒、吸烟史以及既往疾病史等,比较红细胞计数(RBC)、白细胞计数(WBC)、白蛋白(CRP)、血小板(PLT)、低密度脂蛋白(LDL)、肝(谷丙转氨酶ALT)、肾(肌酐Cr)功能、脑钠肽(BNP)以及心脏彩超结果中射血分数(LVEF)、缩短分数(FS)等指标。入选患者于禁食12 h 后留取肘静脉血5 mL 于肝素抗凝试管中,以3000 r/min离心5 min分离血清,将上清液留取至单独的冻存管,-80 ℃冰箱中备用。
1.3 血清FDX1和LA表达水平检测
将血清从-80 ℃冰箱取出融化,使用人FDX1、LA酶联免疫分析(ELISA)试剂盒(上海优选生物科技有限公司)检测血清FDX1、LA水平,检测过程严格按照试剂盒说明书加入标本、标准品、检测抗体等,在操作完成后得到FDX1、LA的吸光度(A值),通过计算出标准曲线的直线回归方程,将A值代入方程式计算出样品浓度,再乘以稀释倍数,得到实际浓度。
1.4 实验动物及高脂血症模型诱导
10周龄雄性ApoE-/-小鼠15只,品系C57BL/6J,体质量20±2 g,15只相同遗传背景和周龄的雄性C57BL/6J小鼠,体质量20±2 g,购自常州卡文斯实验动物有限公司,均为SPF 级动物,许可证号:SCXK(苏)2021-0013。动物房温度设置为(20~22)℃,明暗周期12∶12,环境的湿度60%。用标准饲料适应性喂养5 d后,高脂高胆固醇饲料喂养12周诱导高脂血症模型,对照组设为标准饲料喂养的同周龄15只C57BL/6J小鼠,研究方案获得本院伦理委员会批准(伦动科批字[2023]第546号),高脂饲料(2.0%胆固醇,2.5%猪胆盐,10.0%猪油加入小鼠标准饲料中混匀)由常州卡文斯实验动物有限公司提供。
1.5 小鼠血清生化指标检测
为确定高脂血症模型是否制备成功,12周后全部处死小鼠,随后对各组别小鼠分别进行眼球取血,静置1 h,3000 r/min离心10 min,经全自动生化分析仪(深圳雷杜生命科技股份有限公司)检测TC、TG、LDL和HDL水平,根据相关公式计算出AS指数:(TC-HDL)/HDL,TG、TC、HDL、LDL测定试剂盒为全自动生化分析仪配套试剂盒由深圳雷杜生命科技股份有限公司提供。
1.6 小鼠主动脉切片HE染色
12周后停止药物干预,将两组小鼠处死,解剖小鼠胸主动脉组织,制备石腊,按常规方法进行脱蜡,水合,分两次使用二甲苯将切片浸泡5 min,再分别在无水乙醇、95%乙醇、85%乙醇、70%乙醇中浸泡5 min,使用PBS将切片浸洗3次,在苏木素-伊红染液试剂盒(江苏凯基生物技术股份有限公司)中试剂一核染液染缸内染色3~5 min,水洗约30~60 s;浸入试剂盒的二分色液I中约20 s,再次水洗;浸入试剂盒中试剂三分色液II中约40 s后水洗;最后置于试剂盒中试剂四浆染液中染色2 min,最后多余染液使用试剂五增色液去除,滤纸吸干封片,普通光学显微镜下观察两组血管形态。
1.7 小鼠主动脉切片天狼猩红染色
按常规方法进行脱蜡,水合,分两次使用二甲苯将切片浸泡5 min,再分别在无水乙醇、95%乙醇、85%乙醇、70%乙醇中浸泡5 min,自来水将切片清洗3次。0.1 g天狼猩红(索莱宝生物科技有限公司)溶解于100 mL苦味酸饱和液(索莱宝生物科技有限公司)中,配成苦味酸天狼猩红工作液。将切片置于入苦味酸天狼猩红染液染色2 h,并用自来水稍洗,晾干,中性树胶封片,使用偏振光显微镜下观察血管胶原蛋白沉积。
1.8 小鼠主动脉切片免疫组化染色
按常规方法进行脱蜡,水合,分2次使用二甲苯将切片浸泡5 min。再分别在无水乙醇、95%乙醇、85%乙醇、70%乙醇中浸泡5 min,使用PBS将切片浸洗3次。把切片放入抗原修复缓冲液的染色盒内,使用微波炉加热10 min,取出清洗冷却后用PBS 浸洗3 次,每张切片滴加2 滴3%H2O2-甲醇溶液,室温(15~25 ℃)封闭10 min。滴加1%BSA50~100 μL,室温孵育20 min。滴加FDX1(英国Abcam)1∶200、LA(英国Abcam)1∶100一抗50~100 μL,4 ℃过夜。加入羊抗兔IgG(英国Abcam)1∶200二抗50 μL,使用DAB试剂盒(福州迈新生物科技有限公司)显色,苏木素染液复染10 min,显微镜下观察棕黄色颗粒沉积作为阳性。以Image J 软件进行半定量分析,计算单位面积平均光密度值(MOD)值。
1.9 RT-qPCR 检测小鼠主动脉FDX1、硫辛酸合成酶(LIAS)和线粒体顺乌头酸酶(ACO2)mRNA表达
称取适量的胸主动脉样本,将剩余组织冻存,加入1 mL的Trizol试剂(赛默飞世尔科技公司),研磨粉碎胸主动脉后,萃取小鼠主动脉组织的总RNA,TAKARA逆转录试剂盒(南京诺维赞生物科技股份有限公司)处理RNA,将总RNA 转成cRNA,4℃保存,冰上解冻cDNA和qPCR试剂盒(南京诺维赞生物科技股份有限公司),配置qPCR反应液,拨动八联管至充分混匀,气泡消失后可放置于荧光定量PCR仪中进行扩增。按下列条件进行qPCR反应:95℃预变性30 s;然后95 ℃5 s,60 ℃30 s,共进行40个循环。溶解曲线:95 ℃5 s;60 ℃1 min。降温:50 ℃30 s,1个循环。内参基因β-actin、FDX1、LIAS、ACO2引物由北京擎科生物科技股份有限公司生产。采用2-△△CT的方法计算目的基因的相对表达量(表2)。

表2 引物序列Tab.3 Primer sequence for RT-PCR
1.10 统计学方法
最终的数据分析,采用SPSS26.0软件,计量资料采用均数±标准差表示,计数资料用例数(n)及百分率(%)表示;组间比较采用t检验、方差分析、卡方检验;对于指标间的相关性检测,使用Pearson 法分析;使用软件Image pro plus对图片进行测量分析;当Ρ<0.05表示差异具有统计学意义,所有的实验都是独立重复3次。
2 结果
2.1 受试者临床资料比较
c-CHD组LVEF、FS显著低于对照组(Ρ<0.01),肝肾功能、血常规等相关指标差异均无统计学意义(Ρ>0.05,表3)。

表3 基线资料比较Tab.3 Baseline data of the patients indifferent groups
2.2 血清FDX1、LA表达水平和Gensini评分在不同冠脉狭窄中的变化
与对照组相比,a-CHD组、b-CHD组和c-CHD组的血清FDX1水平显著减低(Ρ<0.05~Ρ<0.01),随冠状动脉狭窄进程加重血清FDX1、LA水平逐渐降低,差异有统计学意义(Ρ<0.01)。Gensini评分随冠状动脉狭窄程度加重逐渐增高,各组之间评分比较差异均有统计学意义(Ρ<0.05,表4)。

表4 不同冠脉狭窄患者血清FDX1、LA表达水平和Gensini评分比较Tab.3 Comparison of serum FDX1 and LA levels and Gensini score in patients with different severities of coronary stenoses
2.3 血清FDX1、LA表达水平与病变支数的关系
冠状动脉造影结果显示各组患者冠状动脉病变支数不同,将实验组患者分为单支组、双支组及多支病变组(3支及以上),分析得出,单支组、双支组和多支组血清FDX1、LA较对照组相比显著降低(Ρ<0.05,表5)。

表5 血清FDX1、LA表达水平与病变支数的关系Tab.3 Correlation of serum FDX1 and LAlevels with the number of lesion branches
2.4 血清FDX1、LA表达水平与Gensini评分的相关性
血清FDX1 水平与Gensini 评分呈负相关关系(r=-0.241,Ρ<0.01),LA水平与Gensini 评分呈负相关关系(r=-0.273,Ρ<0.01)。
2.5 ApoE-/-小鼠高脂血症模型制备
为确定高脂血症模型是否成功,连续高脂饮食12周后,随机从对照组和ApoE-/-模型组中采集样本。小鼠血清学检测结果显示,ApoE-/-模型组小鼠TG、TC、LDL水平,与对照组相比均明显上升(Ρ<0.01);HDL水平显著下降(Ρ<0.01),AS指数明显增高(Ρ<0.01,表6)。
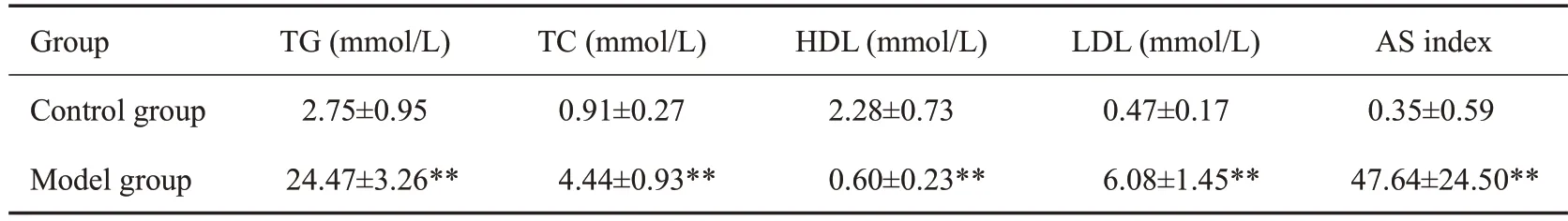
表6 两组小鼠TG、TC、HDL、LDL、AS指数水平Tab.3 Levels of TG,TC,HDL,LDLandAS index in normal andApoE-/-mice
2.6 小鼠主动脉切片HE染色结果
两组小鼠胸主动脉HE染色显示,C57正常对照组,血管层次分明,内中外三层膜结构清晰可见,中膜细胞排列规律整齐,未见有病理性变化;ApoE-/-模型组血管中膜增厚,中膜中弹性纤维间距增大(如黑色箭头)伴有脂质聚集。间隙内出现炎性细胞(棕黄色箭头),弹性纤维被破坏,出现断裂(如蓝色箭头),(图1A)。

图1 两组小鼠主动脉HE染色、天狼猩红染色结果Fig.1 HE staining(A)and Sirius red staining(B)of the aorta of normal and ApoE-/-mice(Scale bar=20µm).
2.7 小鼠主动脉切片天狼猩红染色结果
在偏振光显微镜下观察小鼠主动脉天狼猩红染色显示,C57正常对照组I型和III型纤维紧密排列,I型胶原纤维和III型胶原纤维占比适中;ApoE-/-模型组I型胶原显著增加,III型胶原显著减少,III型胶原纤维紧断断续续排列与I型胶原纤维之中,I型胶原纤维占比显著高于III型胶原纤维。与C57正常对照组相比,ApoE-/-模型组胸主动脉胶原纤维增生明显,血管纤维化程度明显增加(图1B)。
2.8 FDX1、LA的免疫组织化学染色结果
免疫组化中FDX1、LA染色阳性信号为棕色颗粒,表达在细胞质中,结果显示ApoE-/-模型组主动脉的FDX1、LA 表达量明显低于C57 正常对照组;两组间FDX1 表达MOD 值差异有统计学意义(F=0.620,Ρ<0.01),两组间LA表达MOD值差异有统计学意义(F=0.397,Ρ<0.05,图2)。

图2 FDX1、LA免疫组化染色结果Fig.2 Immunohistochemical staining of FDX1(A) and LA (B) in the aorta of normal and ApoE-/-mice and quantitative analysis of their expression levels(Scale bar=100µm).*P<0.05,**P<0.01 vs control group.
2.9 RT-qPCR 检测小鼠主动脉FDX1、LIAS、ACO2 mRNA表达结果
与对照组比较,ApoE-/-模型组主动脉FDX1 mRNA表达明显降低(Ρ<0.01);与C57 正常对照组比较,ApoE-/-模型组LIAS mRNA表达明显降低(Ρ<0.001);与C57正常对照组比较,ApoE-/-模型组ACO2 mRNA表达明显降低(Ρ<0.01,图3)。

图3 FDX1、LIAS、ACO2的mRNA相对表达量Fig.3 FDX1(A),LIAS(B)and ACO2(C)mRNA expression levels in the aorta of normal and ApoE-/-mice.*P<0.05,**P<0.01,***P<0.001 vs control group.
3 讨论
AS的发生是在微血管阻力增加、脂质代谢异常、纤维组织增生等因素共同作用下导致的血管病变[12]。研究表明,高脂血症与CHD的发生、发展密切相关,同时也是导致CHD的一个独立的重要危险因素[13],血浆中增多的胆固醇脂质易沉积于动脉内膜,引起结缔组织纤维化,使动脉增厚和变硬。当单核-巨噬细胞迁移至内皮损伤部位,吸收脂质形成巨噬细胞源性泡沫细胞,从而促进AS进展[14]。在临床实践中血脂水平经常被用于估计AS等心血管疾病的风险,并作为不良心血管事件一级预防的治疗目标。Fang[15]通过荟萃分析证实茶黄素通过抑制胶束形成以减少胆固醇吸收来改善高胆固醇血症,减轻了动脉粥样硬化程度,从而预防冠状动脉疾病的发生。因此在探究CHD发病机制的同时,研究高脂血症相关血管病变的机制对于CHD的防治也有着至关重要的临床意义。
越来越多的研究表明线粒体的代谢与AS的发生息息相关,并参与了CHD中能量代谢、氧化应激、钙调节和细胞死亡的多个病理生理过程[16-18]。当参与线粒体的代谢的信号分子表达发生变化,如线粒体活性氧簇增加、ATP减少等,影响线粒体及细胞功能,进而引发相关生物应激反应,最终导致心血管相关疾病的发生[19]。FDX1是线粒体代谢相关的基因的一种,归属于Fe-S簇蛋白家族[20]。Sheftel等[21]研究显示,FDX1利用类固醇合成急性调节蛋白将胞质内胆固醇转移到线粒体,接着胆固醇侧链裂解酶将胆固醇进一步转化为孕烯醇酮,对于启动类固醇激素的合成至关重要。Cai等[22,23]通过实验发现,对于脂酰合成,FDX1将电子转移到自由基S-腺苷蛋氨酸依赖性脂酰合酶中以启动其自由基链反应,且FDX1在血红素a和脂酰辅因子生物合成中具有附加和特异性作用。当FDX1缺乏时Fe-S簇蛋白生物表达降低,线粒体中细胞铁摄取和铁积累增加,会破坏铁硫簇的组装和维持正常的胞质和线粒体铁稳态,最终导致细胞凋亡和心血管功能紊乱[24]。Yuan等[25]通过研究证实,铜死亡诱导剂FDX1可能参与心力衰竭的病理生理过程,是导致线粒体蛋白高乙酰化的主要原因,有可能成为心力衰竭新型治疗机制的关键靶点。以上研究均提示FDX1可能通过调节线粒体代谢过程参与了心血管疾病的发生发展。
LA是线粒体代谢过程的重要辅助因子,也是目前研究较广泛的抗氧化剂[26]。国内外LA与CHD的相关文献均表明,LA具有清除活性氧、金属鳌合、修复氧化损伤的能力,在动脉粥样硬化过程中的氧化应激、炎症反应等多个病理机制中均发挥着重要调节作用[27,28]。Lian等[29]通过研究证实,内源性LA在线粒体中通过脂肪酸合成通路II(mtFAS II),由LIAS 等关键酶催化合成[30],成为蛋白质脂酰化的重要原料。当阻断线粒体LA合成,造成蛋白脂酸化无法正常进行,线粒体会出现分解代谢障碍,引起细胞能量不足,表现为胚胎发育、神经和心血管功能紊乱[31,32]。WANG等[33]通过实验发现,LA能降低心肌的缺血性损伤,其可能是心肌缺血/再灌注损伤的内源性细胞保护重要介质。因此,通过探究LA的上游调控因子从而促进内源性LA合成具有重要临床意义[34]。Tscetkov等[35]研究表明,FDX1和蛋白质脂酰化是调控铜依赖性细胞死亡—铜死亡的关键因子,FDX1可上游调节线粒体蛋白质脂酰化过程,而LA是蛋白质脂酰化的关键原料,此外Joshi 等[4]也证实了FDX1 对于硫辛酸辅因子的产生至关重要,基于以上研究提示:FDX1可能通过上调内源性LA的表达共同参与了心血管疾病的发生发展,可能与冠状动脉粥样硬化的形成有潜在联系,但具体机制尚不明确。
本研究首次发现,冠脉造影异常的患者血清FDX1、LA水平与正常对照组相比明显降低,且随着冠脉狭窄程度的加重血清FDX1、LA水平逐渐减低,提示血清FDX1、LA水平变化可能与冠脉狭窄程度有关。另外,单支病变的患者血清FDX1、LA水平最高,随着病变支数的增加,血清FDX1、LA 水平逐渐降低,表明FDX1、LA均可以对冠脉病变情况造成影响。临床上,Gensini评分可以反映冠心病患者冠脉病变严重程度[36],Pearson检验分析发现,实验组患者血清FDX1、LA水平与Gensini 积分呈负相关,Gensini 积分越高,血清FDX1、LA 水平越低;而FDX1 与LA 呈正相关,这与Tscetkov等的研究结论一致[35]。Huo等[37]通过铜离子载体Elesclomol在心肌细胞中诱导铜死亡,发现其特征是Fe-S簇蛋白的减少和线粒体酶脂酰化的减少,多种线粒体Fe-S簇蛋白如FDX1、LIAS、ACO2等的表达均明显下降。为了探究FDX1、LA的表达是否与脂代谢异常导致的血管病变有关,本研究中成功建立了高脂血症小鼠模型,利用免疫组织化学染色、RT-PCR等方法,首次证实了在高脂血症模型中主动脉FDX1、LA的表达较正常对照组明显下降,同时LIAS、ACO2等的Fe-S簇蛋白表达也明显降低,提示,FDX1、LA的表达与高脂血症所致的动脉病变有关。
综上所述,冠脉狭窄患者血清FDX1、LA 水平降低,两者水平变化与冠脉病变支数和Gensini积分有关,且两者共同参与了冠状动脉粥样硬化的发生与进程。该结果为判断冠脉病变程度及研究CHD致病机制提供了一定参考依据。血脂异常作为冠状动脉粥样硬化的独立危险因素,通过动物模型进一步证实了FDX1、LA与高脂血症导致的血管病变有关,为CHD的防治提供参考,尽管本实验证实了FDX1、LA在冠状动脉粥样硬化冠脉病变、高脂血症所致血管损伤之间的相关性,但其具体作用机制及作用途径仍需深入研究。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
中国卫生标准管理(2015年24期)2016-01-14
分子影像学杂志(2015年3期)2015-12-04
中国当代医药(2015年21期)2015-03-01
癌变·畸变·突变(2014年1期)2014-03-01
实验动物与比较医学(2014年5期)2014-02-28
哈尔滨医药(2014年4期)2014-02-27
中国医学科学院学报(2013年2期)2013-03-11
