
Platycodin D inhibits angiogenic vascular mimicry in NSCLC by regulating the eIF4E-mediated RNA methylome
2024-03-21 05:51ShuyuZhengYnlinXinJiminLinZejunXieKeyuChengShnshnWngWenliLuHoYngTinmingLuJunLiRuoguQiYunyunGuo
Shuyu Zheng , Ynlin Xin , Jimin Lin , Zejun Xie , Keyu Cheng ,Shnshn Wng , Wenli Lu , Ho Yng , Tinming Lu , Jun Li , Ruogu Qi ,**,Yunyun Guo ,*
a Department of Biochemistry and Molecular Biology,School of Medicine&Holistic Integrative Medicine,Nanjing University of Chinese Medicine,Nanjing,210023, China
b Department of Neurosurgery, The Second Affiliated Hospital, Zhejiang University, Hangzhou, 310009, China
The treatment of non-small cell lung cancer (NSCLC) remains a challenge due to tumor evolution during anti-angiogenesis therapies,in which the mechanism of vascular mimicry(VM)is believed to result in ineffective treatment [1].To conquer this challenge,substantial effort has recently been devoted to seeking out natural compounds on account of their multitarget actions.As a traditional herbal medicine,platycodin D(PD)is the major bioactive monomer derived from Platycodon grandiflorum(P.grandiflorum)and is used as an expectorant for pulmonary disease in Asia[2].We previously found that PD exhibits anticancer effects on NSCLC through noncoding RNA(ncRNA)regulation[3].Although the RNA methylome is a hotspot in ncRNA regulation studies, especially for N7-methylguanosine (m7G) modification, in-depth pharmacological studies related to this topic are limited [4].Herein, we aimed to study the anti-VM function and potential long noncoding RNA (lncRNA)/m7G regulatory mechanism of PD in NSCLC.We found that PD could not only inhibit VM formation in vitro in NSCLC but also block vessel generation in vivo,which was mediated by the m7G gene eukaryotic translation initiation factor 4E (eIF4E).In addition, the combination of PD and the eIF4E inhibitor ribavirin exerted synergistic anti-VM effects on NSCLC via destabilization of epidermal growth factor receptor (EGFR).In summary, we found that PD could inhibit VM in NSCLC via lncRNA/eIF4E/EGFR regulation, revealing the potential of PD for anti-VM treatment in NSCLC patients.
We first investigated the effect of PD on NSCLC angiogenesis and VM through gene set enrichment analysis (GSEA) of PD-regulated DEm7G-lncRNAs and DEm7G-genes.The results indicated that the influenced core differentially expressed genes were mainly related to angiogenesis, vascular mimicry, tumor migration, and RNA modification,including pre-messenger RNA (mRNA)capping,the vascular endothelial-derived growth factor (VEGF)-VEGF receptor (VEGFR)signaling pathway,extracellular matrix organization,focal adhesion,and the integrin 1 pathway(Figs.1A and S1).To validate this,we then assessed the influence of PD on the vascular mimicry of NSCLC cells.According to the results from the capillary-like tube formation assay in Fig.1B,compared to the control group,there was a decrease in the angiogenesis rate of A549 cells.Moreover,conditioned medium from PD-treated A549 cells significantly inhibited HUVEC tube formation compared to control non-PD-treated A549 cell medium(Figs.1B and C).In addition,the in vitro extravasation model demonstrated a dosedependent effect of PD on the inhibition of A549 cell extravasation(Figs.1D and E).Furthermore, in vivo animal model experiments demonstrated that 7.5 mg/kg PD treatment could significantly inhibit the formation of microvessels, VM, and the expression levels of vascular endothelial growth factor A(VEGFA)/VEGFR in both the lung and liver (Table S1 and Figs.1F and H).More importantly, PD treatment resulted in near complete inhibition of angiogenesis and small capillary formation in the in vivo Matrigel plug assay(Figs.1I and J).

Fig.1.In vitro and in vivo evaluation the anti-angiogenesis and anti-vascular mimicry(VM)formation effect of platycodin D(PD)on non-small cell lung cancer(NSCLC).(A)Gene set enrichment analysis (GSEA) displayed the top 5 enriched pathways in PD-regulated N7-methylguanosine (m7G) and long non-coding RNAs (lncRNAs).(B, C) Tube formation and vasculogenic mimicry formation assays showed PD could decreased the number of tube nodes both in A549 cells and human umbilical vein endothelial (HUVEC) cells.(D) The in vitro extravasation model was built based on co-cultivation of A549 (stained with calcein AM) and HUVEC cells.The A549 cells migrated across the barrier of HUVEC and the membrane of Transwell chambers was photographed by inverted fluorescent microscopy.(E) The number of extravasated A549 cells was decreased under the treatment of PD,compared to the control.(F,G)The image(F)and quantitative analysis(G)of micro-vessel and VM in the lung and liver sections of mice with or without PD treatment.Brown arrows indicate typical blood vessels with brown CD31+ staining and red arrows indicate VM channels (CD31-/PAS+).(H) Vascular endothelial-derived growth factor (VEGF) receptor(VEGFR),vascular endothelial growth factor A(VEGFA),and Hoechst co-staining in the lung and liver sections of mice with or without PD treatment.(I,J)Matrigel plug assay showed that PD treatment could inhibit angiogenesis in vivo.The Matrigel group was used as the negative control,and the placebo group was used as the positive control.The hemoglobin concentration was significantly decreased in the PD-treated group, compared to the placebo control (P <0.001).Concentration of PD treatment: 7.5 mg/kg.*P <0.05, **P <0.01,***P <0.001, and ****P <0.0001.VM: vascular mimicry.
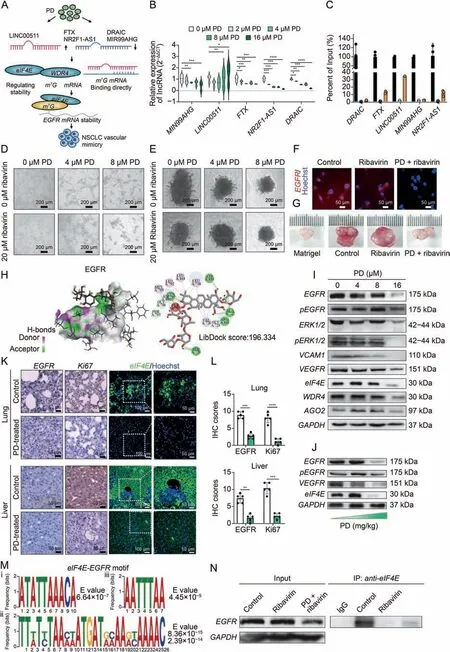
Fig.2.Eukaryotic translation initiation factor 4E (eIF4E) mediates platycodin D (PD) induced anti-angiogenesis and anti-vascular mimicry (VM) formation via destabilization of epidermal growth factor receptor(EGFR).(A)Schematic illustrating potential mechanism by which PD inhibits VM of non-small cell lung cancer(NSCLC).(B)The relative expression of PD-regulated DEm7G-long noncoding RNAs(lncRNAs)was verified by real-time quantitative polymerase chain reaction(qPCR).(C)RNA immunoprecipitation(RIP)for eIF4E and associated lncRNAs followed by qPCR analysis.(D)Vasculogenic mimicry formation assay showed a synergistic inhibitory effect of PD and ribavirin on the number of tube nodes in A549 cells.(E) Spheroid sprouting angiogenesis assay showed that PD combined with ribavirin treatment inhibited the sprouting angiogenesis in a concentration-dependent manner.(F) Confocal fluorescence microscopy indicated the expression of EGFR was synergistically inhibited by PD and ribavirin treatment.(G) Matrigel plug assay showed more than a 1.5-fold decrease in relative hemoglobin content of the plug when added in PD treatment,compared to the ribavirin treatment alone in vivo.Concentration of PD treatment:7.5 mg/kg;concentration of ribavirin treatment:10 mg/kg.(H)Molecular docking exhibited the interaction between PD and EGFR protein.In H-bonds,pink indicates the donor,and green indicates the receptor.The LibDock scores are calculated by Discovery Studio 2019.(I) The relative expression of PD-regulated proteins was validated through Western blotting.(J)The relative expression of EGFR,pEGFR,and VEGFR was decreased in the lung tissue of mice,as the treatment dose of PD increased(0,5,and 7.5 mg/kg).(K,L)EGFR,Ki67,eIF4E, and Hoechst co-staining (K) in the lung and liver sections of mice with or without PD treatment, and quantitative analysis (L).(M) Enriched motifs with sequences in m7G gene eIF4E and EGFR were analyzed through MEME suite.i,ii,and iii represent different motifs estimated by MEME.(N)The direct interaction of eIF4E and EGFR was confirmed by co-immunoprecipitation (co-IP) assay.*P <0.05, **P <0.01, ***P <0.001, and ****P <0.0001.mRNA: messenger RNA; WDR4: WD repeat domain 4; FTX: FTX transcript; DRAIC:downregulated RNA in cancer, inhibitor of cell invasion and migration; pEGFR: phosphorylated EGFR; ERK1/2: extracellular regulated protein kinases 1/2; VCAM1: vascular cell adhesion molecule 1; VEGFR: VEGF receptor; AGO: Argonaute; GAPDH: glyceraldehyde-3-phosphate dehydrogenase; IHC: immunohistochemistry; IP: immunoprecipitation.
To elucidate the molecular mechanism underlying PD-mediated anti-angiogenesis and VM formation, whole transcriptome analysis of PD-regulated ncRNAs was conducted(Fig.2A).The results revealed that MIR99AHG,FTX,NR2F1-AS1,and DRAIC expression were significantly decreased with increasing treatment dose compared to the control (Table S2, P <0.01).Moreover, compared to the control,LINC00511 expressionwas significantly increased(P <0.05,Fig.2B).In addition, the direct interaction between m7G-related lncRNAs and eIF4E was validated through RNA immunoprecipitation(RIP)assays.As expected,a significant enrichment of lncRNA FTX,LINC00511 and NR2F1-AS1 with eIF4E antibody was observed compared with the nonspecific IgG antibody,while there was little enrichment of DRAIC or MIR99AHG (Fig.2C).It is generally accepted that ribavirin could function as a physical mimic of the m7G cap and thereby inhibit eIF4E binding activity,for which ribavirin has been clinically evaluated as a potential anticancer drug.Here,to evaluate the synergistic potency of PD and ribavirin in NSCLC treatment, we first performed a VM formation assay.The results showed that the cotreatment of PD and ribavirin on A549 cells could significantly reduce the number of nodes to an average of 15%of the values in the control(Fig.2D).In the spheroid sprouting assay,a dramatic decrease in total sprout length could be observed in the combinational treatment compared to the single agent treatment(Fig.2E).In addition,confocal results consistently showed that the expression of EGFR was decreased under the combinational treatment of PD and ribavirin(Fig.2F).In vivo,PD and ribavirin also exhibited synergistic inhibitory effects on angiogenesis and small capillary formation in the Matrigel plug assay(Fig.2G).
We further calculated the binding ability of PD and target proteins bycovalent docking-based molecular simulation.As shown in Fig.2H,molecular docking indicated that PD could form a stable covalent bondwiththeactivesiteofEGFR(LibDock score:196.334).Inaddition,functional analysis of PD-regulated DEm7G-lncRNAs and DEm7Ggenes demonstrated that the top enriched Gene Ontology (GO)terms and Kyoto Encyclopedia of Genes and Genomes(KEGG)pathways were associated with EGFR-TKI resistance and RNA m7G cap binding (Fig.S2).The prediction was further validated through Western blotting.PD strongly inhibited the phosphorylation of EGFR,VCAM1,VEGFR,and their downstream signaling proteins and inhibited eIF4E and WDR4 in a dose-dependent manner(Fig.2I).Confocal results consistently showed that the expression of EGFR was decreased under PD treatment (Fig.S2).Moreover, in vivo experiments showed that compared to the control mice,lung tissues from mice treated with different doses of PD(5 and 7.5 mg/kg)exhibited a dose-dependent decrease in the expression levels of pEGFR, EGFR,VEGFR,and eIF4E(Fig.2J).To further validate the effect of PD on EGFR and eIF4E in vivo, we conducted immunohistochemistry (IHC) and immunofluorescence staining of PD-treated mice and control mice.IHC analysis delineated that Ki67-and EGFR-positive cells were substantially decreased in the PD treatment group compared to the control group (Fig.2K).Likewise, immunofluorescence staining demonstrated decreased expression(green)of eIF4E in both the lung and liver of PD-treated mice compared with the control(Figs.2K and L).Since there was a close relationship between PD,eIF4E,and EGFR,we hypothesized that the inhibition of EGFR expressionwas mediated by eIF4E m7G capping of EGFR.To substantiate this,three consensus DNA motifs were found in eIF4E and EGFR binding (Fig.2M).Coimmunoprecipitation (co-IP)experiments validated the direct binding of eIF4E and EGFR.In addition,this binding effect could be disrupted under treatment with PD and ribavirin, indicating the regulation of eIF4E on EGFR mRNA stability,which in turn achieves the purpose of targeted therapy(Fig.2N).
In summary, we found that PD could inhibit angiogenesis and vascular mimicry in NSCLC by regulating the m7G gene eIF4E and related lncRNAs.Moreover,we demonstrated novel mechanisms by which eIF4E suppresses the expression of EGFR, revealing that the combinational treatment of PD and ribavirin is promising for targeted therapy in EGFR + NSCLC patients.
Ethical statement
All methods used in this study were carried out in accordance with national animal care and use guidelines laws, and all experimental protocols were approved by the Institutional Animal Care and Use Committee of Nanjing University of Chinese Medicine,Nanjing,China (Approval No.:202112A068).
CRediT author statement
Shuyu Zheng:Data curation, Investigation, Writing - Original draft preparation;Yanlin Xin:Methodology, Writing - Original draft preparation;Jiamin Lin:Investigation,Visualization,Writing- Original draft preparation;Zejuan Xie:Formal analysis, Data curation;Keyu Cheng:Investigation,Validation;Shanshan Wang:Methodology, Investigation;Wenli Lu:Visualization;Hao Yang:Validation;Tianming Lu:Investigation;Jun Li:Visualization;Ruogu Qi:Writing - Reviewing and Editing, Funding acquisition;Yuanyuan Guo:Funding acquisition, Resources, Supervision,Conceptualization, Project administration.
Declaration of competing interest
The authors declare that there are no conflicts of interest.
Acknowledgments
We thank Prof.Yang Zhao for his kind assistance in HUVEC cultivation and Dr.Xin for her kind assistance in data processing.This research was funded by the National Natural Science Foundation of China (Grant Nos.: 82004081 and 52073145) and the National Natural Science Foundation of Nanjing University of Chinese Medicine, China (Grant No.:NZY82004081).
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jpha.2023.10.003.
 Journal of Pharmaceutical Analysis2024年1期
Journal of Pharmaceutical Analysis2024年1期
- Journal of Pharmaceutical Analysis的其它文章
- Lipid metabolism analysis in esophageal cancer and associated drug discovery
- Push forward LC-MS-based therapeutic drug monitoring and pharmacometabolomics for anti-tuberculosis precision dosing and comprehensive clinical management
- The role of innate immunity in diabetic nephropathy and their therapeutic consequences
- Metformin: A promising clinical therapeutical approach for BPH treatment via inhibiting dysregulated steroid hormones-induced prostatic epithelial cells proliferation
- Epimedin B exhibits pigmentation by increasing tyrosinase family proteins expression, activity, and stability
- Hydralazine represses Fpn ubiquitination to rescue injured neurons via competitive binding to UBA52