
连续间作下的紫花苜蓿/燕麦根系与碳氮代谢特性研究
2024-03-22 01:08汪雪刘晓静王静吴勇童长春
草业学报 2024年3期
汪雪,刘晓静,王静,吴勇,童长春
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
间作是指在同一地块、同一时期以成行或成带(多行)的形式种植2 种及以上相似生长期作物的种植方式,不同作物在生长过程中可通过对资源的利用差异来实现光、肥、水、气、热等有限农业资源的高效利用[1],以提高经济、生态和社会效益。种间根系分布的优化是形成间套作系统优势效应的主要机制之一[2],间作体系内作物会通过调整根系形态的策略来有效吸收和利用土壤养分资源。豆/禾间作时,作物会调控其根系通过空间错位回避邻近根系以减少与邻近根系的重合以降低竞争作用,进而优化间作作物的整体状况。李金婷等[3]在大豆(Glycine max)/玉米(Zea mays)间作的研究中指出,间作系统内玉米根系长度、根体积和根尖数等根系形态的变化是显著促进玉米对氮、磷营养元素吸收量增加的重要原因之一。邵泽强等[4]在紫花苜蓿(Medicago sativa)/玉米间作的研究中发现,间作改善了玉米根长、根表面积、根体积等根系形态指标,提高了其吸氮量,使玉米增产了72.90%。Sekiya 等[5]研究表明,玉米/木豆(Cajanus cajan)间作促进了系统内作物根系空间生态位的优化和分离,改善了系统内作物的种间竞争,提高了作物对水分的吸收利用。间作除了地下部根系的分布状况外,对光能的截获和利用也同样影响着作物对有限农业资源的高效利用,尤其是以光合作用所产生的能量及其中间产物为运行条件的碳、氮代谢,其作为植物体内最重要的两大生理代谢过程,运行的强度和质量直接关乎作物的品质和产量[6],其中,核酮糖-1,5-二磷酸羧化酶(ribulose-1,5-bisphosphate carboxylase,RuBPCase)、硝酸还原酶(nitrate reductase,NR)、谷氨酰胺合成酶(glutamine synthetase,GS)和亚硝酸还原酶(nitrate reductase,NiR)等相关酶活性的高低对碳、氮代谢过程起关键调控作用[7]。邓小燕等[8]在“麦(Triticum aestivum)/玉/豆”套作模式中发现,套作提高了玉米穗位叶硝酸还原酶和谷氨酰胺酶活性,从而提高了作物的氮素利用。本团队前期研究也发现,紫花苜蓿与玉米间作有利于提高玉米的NR 和GS 等氮代谢酶活性及氮积累量[7,9]。在豆/禾间作系统内,不同作物种间既会发生互利促进作用,也会存在养分及空间资源的种间竞争,这种相互作用同时存在于地上部与地下部,进而共同决定间作系统的生产力。然而,目前有关豆/禾间作系统内根系特性、碳氮代谢特性及二者相互协调对其系统生产性能的影响研究多集中在粮食作物/粮食作物间作和粮食作物/牧草间作上,而在豆科牧草/禾本科牧草间作上进行的研究较少。
紫花苜蓿作为多年生豆科牧草,因其特有的固氮方式能增强其对氮素的高效利用[10],故而成为了供家畜转化利用的优质植物性蛋白饲料;燕麦(Avena sativa)属一年生粮饲兼用型禾本科作物,是我国西北干旱地区家畜冬春补饲的重要饲草[11]。大量研究发现[12-15],多年生豆科牧草与一年生禾本科牧草间作不仅会更有利于改变作物根系在土壤中的单一化分布范围,增加土壤氮养分供应和土壤节肢动物,还能直接混收混贮以降低青贮成本、减少污染浪费及解决豆科牧草单一青贮效果不佳等问题。通过紫花苜蓿与燕麦间作不但能提高间作体系的氮吸收及氮互补利用优势[16],而且还能改善群体作物地上部光合性能[17],获得良好的生态效益[18]。此外,也有研究表明[9],单作和间作种植模式下作物的根系形态参数均与碳、氮代谢关键酶呈正相关关系,尤其是根长和根表面积与RuBPCase 和NR 呈极显著正相关,可见,作物地下部的根系活动与其地上部光合作用、碳氮代谢及物质转运等过程是一个不可分割的有机整体,作物的根系会起到承上启下的关键作用,其将吸收的土壤养分及水分运输到地上部,供地上部碳、氮代谢合成有机物质并部分输送到根部,进而形成一个完整的良性循环体系以维持植物体正常的生长与发育[19],目前相关研究主要是针对根系分布特征、根系间相互作用及碳代谢、氮代谢单一特性孤立地解析豆/禾间作的优势效应,而多年生豆科牧草与禾本科牧草间作系统中根系与碳、氮代谢相互耦联机制及其长期效应仍然未知。为此,本研究以框栽土培的方法模拟生产实践中的间作方式,以紫花苜蓿/燕麦间作种植后第2、3 年(高产期)连续两年开展研究,旨在揭示紫花苜蓿/燕麦间作模式下根系与碳、氮代谢的协调机制及其持续效应,为提升牧草间作系统生产潜力提供理论依据与参考。
1 材料与方法
1.1 供试材料
紫花苜蓿品种为LW6010,由北京猛犸种业有限公司提供;燕麦品种为海威,由北京克劳沃公司提供。
1.2 试验设计
试验于2020-2022 年在甘肃农业大学校园内进行。采用框栽土培法,即以紫花苜蓿和燕麦为供试材料种植于高为80 cm,直径为63 cm 的PVC 框内,该框所用的土样均为过2.5 mm 筛的甘肃农业大学牧草试验基地风干土。试验设紫花苜蓿/燕麦间作、紫花苜蓿单作、燕麦单作3 种种植模式,每个处理重复9 次。种植时,先将空PVC 框埋入地下,使框顶端与周围地面齐平,框底端与土壤相通,随后将准备好的风干土装入PVC 框内,最后采用穴播的种植方式将挑选好的颗粒饱满的紫花苜蓿和燕麦种子分别均匀地种植于预先埋好的PVC 框内,间作种植时则将紫花苜蓿均匀地栽植于PVC 框一侧,燕麦栽植于另一侧,播种密度为10 穴·框-1,每穴3~5 粒,出苗前每天浇3 次水,保持土壤正常湿度,待出苗后约3~5 d 浇一次水。播种14 d 后间苗,单作种植下的紫花苜蓿和燕麦每框各留10 株,间作种植下的紫花苜蓿和燕麦每框各留5 株。紫花苜蓿均于初花期[2021 年第1 茬(4 月30 日)、第2 茬(6 月4 日)、第3 茬(7 月10 日);2022 年第1 茬(5 月2 日)、第2 茬(6 月10 日)、第3 茬(7 月14 日)] 进行刈割取样;燕麦于抽穗期进行刈割取样,燕麦取样同时进行紫花苜蓿第3 茬刈割取样,其中,碳、氮代谢关键酶活性和碳水化合物含量的测定位点与光合位点相同,均需在紫花苜蓿和燕麦功能叶片上进行。试验于2020 年3 月7 日播种 紫 花 苜 蓿;于2020 年3 月14 日、2021 年3 月8 日 和2022 年3 月10 日 播 种 燕 麦。
1.3 测定指标
1.3.1 生长特性 地上生物量:紫花苜蓿于初花期(10%开花)刈割取样,燕麦于抽穗期刈割取样,刈割的留茬高度为5 cm;取样时紫花苜蓿和燕麦于每个框里随机选取5 株称其地上鲜重,每个处理重复3 次。
1.3.2 根系特性 根系总长度(root length,RL)、根表面积(root surface area,RSA)、根系平均直径(root average diameter,RAD)和根体积(root volume,RV):挖出种植框后用水洗法将完整的根系从土壤中分离出来,紫花苜蓿和燕麦于每个框里随机选取5 株,迅速用吸水纸吸干,然后将根系放入Epson 根系扫描仪(GXY-A,中国)进行扫描并存入电脑,再用WinRHIZO 根系分析系统软件获得根系总长度、根表面积、根系平均直径和根体积数据,每个处理重复3 次。
1.3.3 碳代谢特性 气体交换参数[净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、胞间CO2浓度(intercellular CO2concentration,Ci)和气孔导度(stomatal conductance,Gs)]:在紫花苜蓿(初花期)、燕麦(抽穗期)取样前选择晴天,采用GFS-3000 便携式光合仪(Walz,德国)于早上9:00 至11:30 快速测定紫花苜蓿和燕麦功能叶片的光合气体交换参数等指标,紫花苜蓿选取顶叶向下第3 片完全展开的健康完整叶片,燕麦选择旗叶,每个处理每种牧草随机测定5 片功能叶,重复3 次。
核酮糖-1,5-二磷酸羧化酶(ribulose-1,5-bisphosphate carboxylase,RuBPCase)活性:用上海杰美科技有限公司生产的RuBP ELISA 试剂盒测定。
碳水化合物:采用Somogyi 法测定[20]。
碳水化合物积累量:为干物质重与碳水化合物含量的乘积。
1.3.4 氮代谢特性 硝酸还原酶(nitrate reductase,NR)、谷氨酰胺合成酶(glutamine synthetase,GS):参照邹琦[21]的方法对植株叶片进行测定。
亚硝酸还原酶(nitrate reductase,NiR):参照Rajasekhar 等[22]的方法对植株叶片进行测定。
谷氨酸合酶(glutamate synthase,GOGAT):参考郑朝峰等[23]的方法对植株叶片进行测定。
氮积累量:为干物质重与氮含量的乘积。
蛋白总量:为干物质重与粗蛋白含量的乘积。
1.4 数据分析
采用SPSS 22 统计软件Duncan 法对数据进行多重比较,利用Excel 2010 软件绘图,P<0.05 为差异显著。
2 结果与分析
2.1 紫花苜蓿/燕麦间作下的地上生物量
由图1 可知,在2021 年,紫花苜蓿地上生物量表现为间作显著低于单作,降低了5.22%,而燕麦地上生物量表现为间作显著大于单作(P<0.05),提高了20.78%。在2022 年,紫花苜蓿地上生物量表现为间作显著低于单作,降低了11.91%,而燕麦地上生物量表现为间作显著大于单作(P<0.05),提高了28.27%。
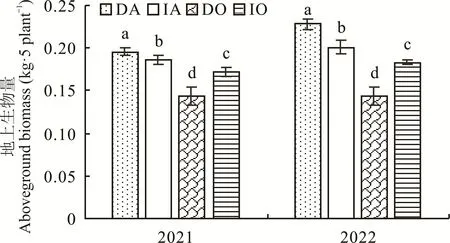
图1 不同种植方式下紫花苜蓿与燕麦的地上生物量Fig. 1 Aboveground biomass of alfalfa and oat under different planting methods
2.2 紫花苜蓿/燕麦间作下的根系特征
2.2.1 总根长、根表面积 由图2 可知,在2021年,紫花苜蓿总根长和根表面积均表现为间作显著低于单作,分别降低了2.58%和6.53%,而燕麦总根长和根表面积均表现为间作显著大于单作(P<0.05),分别提高了8.94%和19.45%。在2022 年,紫花苜蓿和燕麦总根长均表现为间作显著大于单作,分别提高了27.43%和1.19%,而紫花苜蓿根表面积表现为间作显著低于单作(P<0.05),降低了5.60%,燕麦根表面积表现为间作显著大于单作,提高了13.07%。
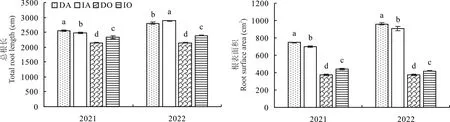
图2 不同种植方式下紫花苜蓿与燕麦的总根长和根表面积Fig.2 Total root length and root surface area of alfalfa and oat under different planting methods
2.2.2 根平均直径、根体积 由图3 可知,在2021 年,紫花苜蓿根平均直径和根体积均表现为间作显著低于单作,分别降低了3.54%和4.44%,而燕麦根平均直径和根体积均表现为间作显著大于单作(P<0.05),分别提高了12.69%和9.47%。在2022 年,紫花苜蓿根平均直径和根体积均表现为间作显著低于单作,分别降低了12.94%和4.38%,而燕麦根平均直径表现为间作显著大于单作(P<0.05),提高了10.30%,燕麦根体积表现为单作和间作之间无显著性差异。

图3 不同种植方式下紫花苜蓿与燕麦的根平均直径和根体积Fig.3 Root average diameter and root volume of alfalfa and oat under different planting methods
2.3 紫花苜蓿/燕麦间作下的碳代谢特性
2.3.1 光合气体交换参数 由表1 可知,在2021 年,紫花苜净光合速率(Pn)和气孔导度(Gs)均表现为间作显著低于单作,分别降低了17.60%和30.15%,燕麦Tr和Gs均表现为间作显著大于单作(P<0.05),分别提高了18.54%和55.07%,而紫花苜蓿胞间CO2浓度(Ci)表现为间作显著大于单作,提高了5.63%。在2022 年,紫花苜蓿Tr、Pn和Gs均表现为间作显著低于单作,分别降低了24.32%、14.85%和11.20%,燕麦Tr和Pn均表现为间作显著大于单作(P<0.05),分别提高了11.10%和44.27%,而紫花苜蓿Ci表现为间作显著大于单作,提高了20.36%。

表1 不同种植方式下紫花苜蓿与燕麦的光合气体交换参数Table 1 Photosynthetic gas exchange parameters of alfalfa and oat under different planting methods
2.3.2 核 酮 糖-1,5-二 磷 酸 羧 化 酶(RuBPCase)由图4 可知,在2021 年,紫花苜蓿核酮糖-1,5-二磷酸羧化酶(RuBPCase)活性表现为间作显著低于单作,降低了6.09%,而燕麦RuBPCase 活性表现为间作显著大于单作(P<0.05),提高了18.56%。在2022 年,紫花苜蓿RuBPCase 活性表现为间作显著低于单作,降低了4.54%,而燕麦RuBPCase 活性表现为间作显著大于单作(P<0.05),提高了20.64%。

图4 不同种植方式下紫花苜蓿与燕麦的核酮糖-1,5-二磷酸羧化酶活性Fig. 4 RuBPCase activities of alfalfa and oat under different planting methods
2.3.3 碳水化合物含量及积累量 由表2 可知,在2021 年,紫花苜蓿碳水化合物含量表现为间作显著低于单作,降低了4.70%,而燕麦碳水化合物含量和碳水化合物积累量均表现为间作显著大于单作(P<0.05),分别提高了8.36%和37.36%。在2022 年,紫花苜蓿碳水化合物积累量表现为间作显著低于单作,降低了17.88%,而燕麦碳水化合物含量和碳水化合物积累量均表现为间作显著大于单作(P<0.05),分别提高了9.41%和37.91%。
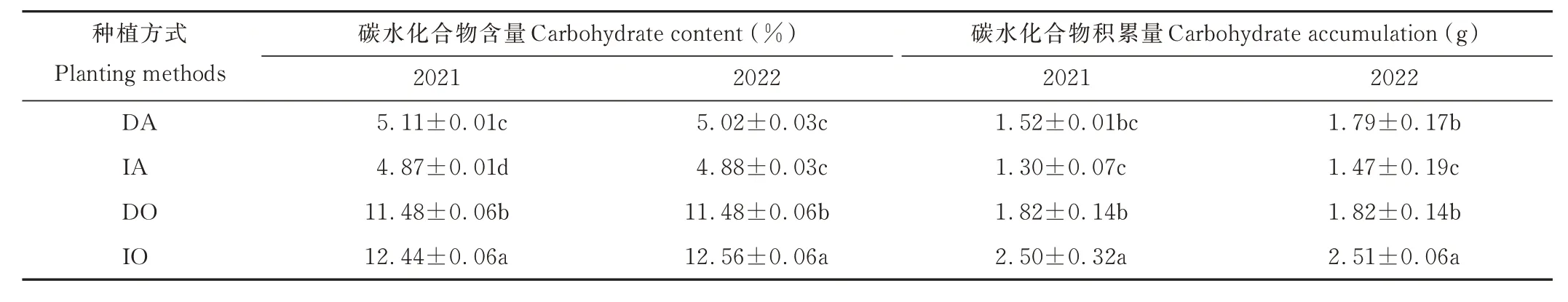
表2 不同种植方式下紫花苜蓿与燕麦的碳水化合物含量及积累量Table 2 Carbohydrate content and accumulation of alfalfa and oat under different planting methods
2.4 紫花苜蓿/燕麦间作下的氮代谢特性
2.4.1 硝酸还原酶(NR)、亚硝酸还原酶(NiR) 由图5 可知,在2021 年,紫花苜蓿硝酸还原酶(NR)活性表现为间作显著低于单作,降低了7.90%,而燕麦NR 和亚硝酸还原酶(NiR)活性均表现为间作显著大于单作(P<0.05),分别提高了13.97%和7.04%。在2022 年,紫花苜蓿NR 活性表现为间作显著低于单作,降低了7.50%,而燕麦NR 和NiR 活性均表现为间作显著大于单作(P<0.05),分别提高了20.65%和12.30%。
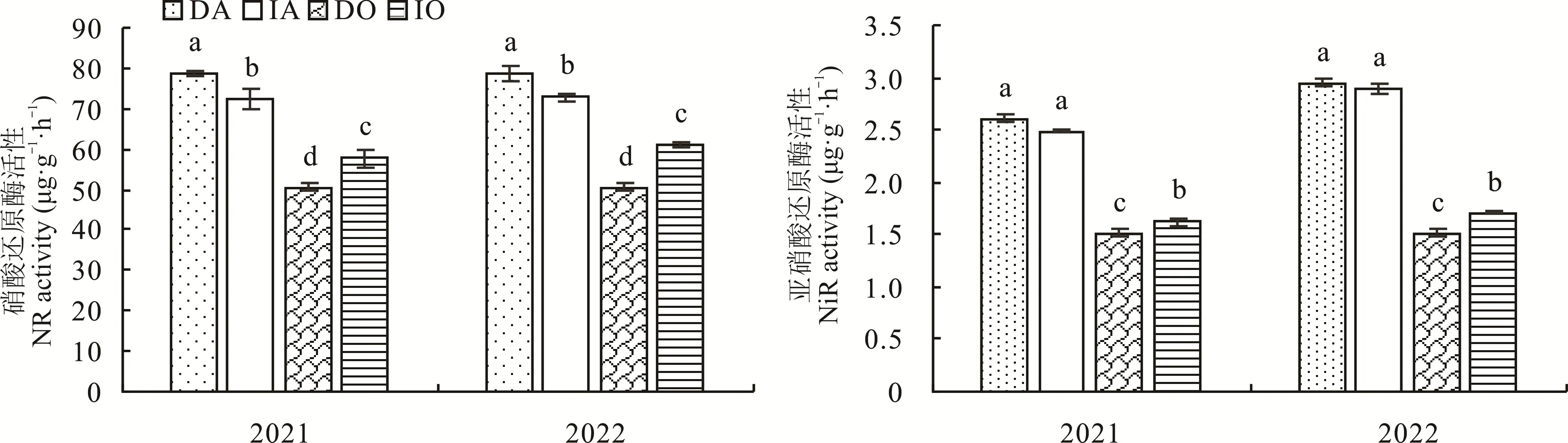
图5 不同种植方式下紫花苜蓿与燕麦的硝酸还原酶和亚硝酸还原酶活性Fig.5 NR and NiR activities of alfalfa and oat under different planting methods
2.4.2 谷氨酰胺合成酶(GS)、谷氨酸合酶(GOGAT) 由图6 可知,在2021 年,紫花苜蓿谷氨酸合酶(GOGAT)活性表现为间作显著低于单作,降低了1.07%,而燕麦谷氨酰胺合成酶(GS)和GOGAT 活性均表现为间作显著大于单作(P<0.05),分别提高了17.81%和4.97%。在2022 年,紫花苜蓿GS 活性表现为间作显著低于单作,降低了6.73%,而燕麦GS 和GOGAT 活性均表现为间作显著大于单作(P<0.05),分别提高了16.88%和3.46%。

图6 不同种植方式下紫花苜蓿与燕麦的谷氨酰胺合成酶和谷氨酸合酶活性Fig.6 GS and GOGAT activities of alfalfa and oat under different planting methods
2.4.3 氮积累量及蛋白总量 由表3 可知,在2021 年,紫花苜蓿氮积累量和蛋白总量均表现为间作显著低于单作(P<0.05),分别降低了18.10%和18.42%,而燕麦氮积累量和蛋白总量均表现为单作和间作差异不显著。在2022 年,紫花苜蓿氮积累量和蛋白总量均表现为间作显著低于单作(P<0.05),分别降低了27.05% 和27.20%,而燕麦氮积累量和蛋白总量均表现为单作和间作差异不显著。
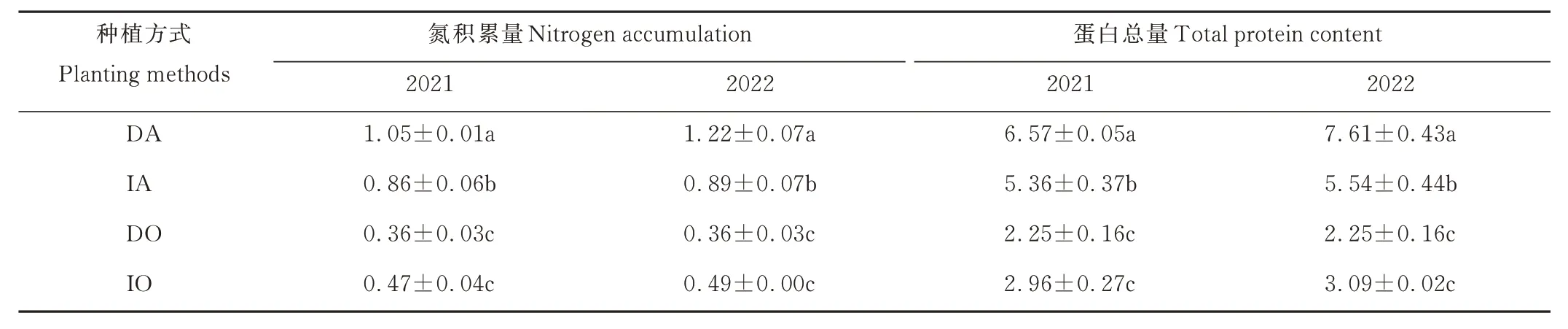
表3 不同种植方式下紫花苜蓿与燕麦的氮积累量和蛋白总量Table 3 N accumulation and total protein content of alfalfa and oat under different planting methods(g)
2.5 根系与碳、氮代谢指标的相关性
由表4 可知,地上生物量与光合气体交换参数、氮代谢特性、根系特性呈正相关。根长(RL)与Pn、NR、NiR、GS、GOGAT 活性呈显著正相关(P<0.05)。根表面积(RSA)与Tr、Pn和Gs、NR 活性、氮积累量、蛋白总量呈显著正相关(P<0.05),与NiR、GS、GOGAT 活性呈极显著正相关(P<0.01)。根平均直径(RAD)与Tr、Pn和Gs、NR 活性、GS 活性、氮积累量、蛋白总量呈显著正相关(P<0.05),与NiR、GOGAT 活性呈极显著正相关(P<0.01)。根体积(RV)与Pn、Gs、NR、GS 活性、氮积累量、蛋白总量呈显著正相关(P<0.05),与NiR、GOGAT 活性呈极显著正相关(P<0.01)。
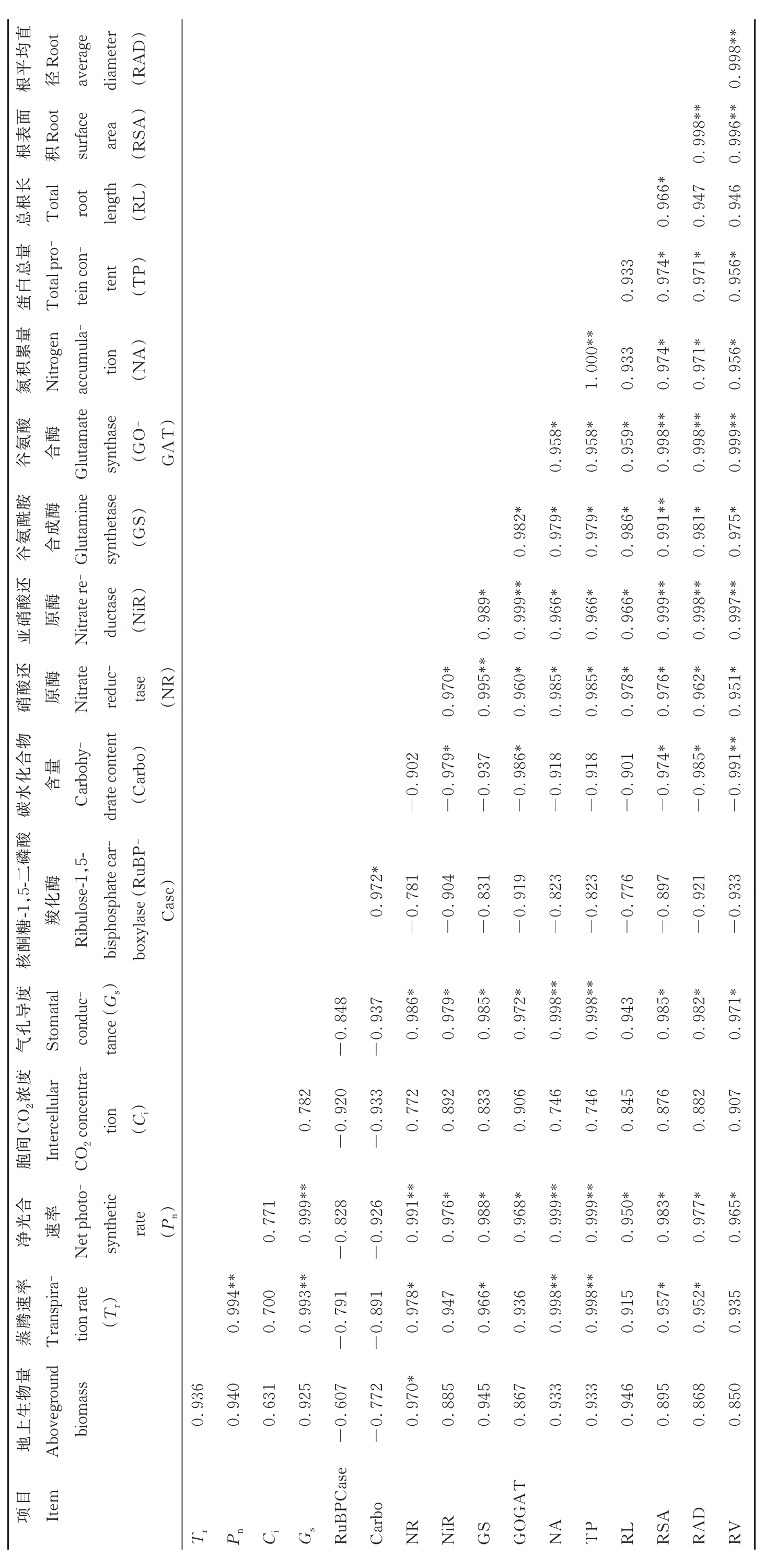
表4 根系与碳、氮代谢指标的相关性Table 4 The correlation between root system and carbon-nitrogen metabolism indicators
3 讨论
间作体系内作物根系构型的调整或重塑是土壤养分及空间资源获取和利用的重要特征[24]。本研究中,与单作相比,间作显著提高了燕麦的总根长、根表面积和根平均直径,而显著降低了紫花苜蓿根表面积、根平均直径和根体积,邵泽强等[4]在紫花苜蓿/玉米间作对根系形态影响的研究中也有此结果,说明紫花苜蓿与燕麦间作中存在种间竞争关系,在此间作体系内禾本科牧草燕麦是优势作物[25],间作能显著改善燕麦根系形态以拓展和延伸其根系侵占邻近作物紫花苜蓿的养分和空间资源,加强其对自身及紫花苜蓿周围环境氮素的竞争和利用[16],而处于空间分布和氮素竞争劣势的紫花苜蓿根系形态参数会减小。但是,紫花苜蓿/燕麦间作体系内存在种间竞争的同时也存在种间促进,尤其是种间氮营养互馈,燕麦对氮素强烈的竞争会刺激紫花苜蓿增强结瘤固氮,进而加强间作系统内种间氮营养的互馈以改善燕麦根系形态[26],促使紫花苜蓿增加根长以获取深层土壤中的养分。也有研究表明[27-28],间作系统内根系特性的优化也取决于根系分泌物的化感促进作用及土壤中难溶性磷的活化等。此外,本研究还发现,随着种植年份的增加,无论单作还是间作下,紫花苜蓿总根长、根表面积和根体积均随之增加,且间作中紫花苜蓿根长提升幅度增大,燕麦对紫花苜蓿根表面积和根体积的抑制逐渐减弱,燕麦在第2 和第3 年间作下的根系形态参数均高于单作,可见,随着年份的增加,根系互作越来越紧密,越有利于间作系统内紫花苜蓿根系在浅层土壤中向水平方向扩展,根系生物量重心下移至深层土壤,紫花苜蓿与燕麦实现根系空间互补,进而形成资源“互利互补”。另外,在紫花苜蓿/燕麦间作体系中,间作燕麦地上生物量和蛋白总量较单作明显提高,其很重要的一方面在于燕麦根系构型的调整和重塑,同时燕麦根系形态的优化也加强了其对碳氮代谢的显著促进作用,进而形成了一个完整的良性循环体系,而紫花苜蓿根系生长及碳、氮代谢在间作模式下受到了抑制,致使其生物量和蛋白总量较单作低,这也从另一个侧面证实了间作可通过重塑根系构型、优化根系空间分布以及协调碳氮代谢以提高牧草生物量和蛋白总量的结论[8,29]。
光合作用是碳、氮代谢的主要能源物质获取渠道,碳代谢为氮代谢提供能量和碳骨架,RuBPCase、NR 等均为碳氮代谢运转的关键酶[26,30]。本研究中,紫花苜蓿与燕麦间作显著提高了燕麦的Pn、Tr、Gs和RuBPCase 活性,显著增加了其体内碳水化合物的合成及积累,而对紫花苜蓿有抑制作用,这在玉米/紫花苜蓿间作[31]、紫花苜蓿/燕麦间作[32]和燕麦/马铃薯(Solanum tuberosum)间作[33]的研究中也得到了相似结论,可能是紫花苜蓿与燕麦间作有利于地上部形成良好的“伞状”冠层结构,促进豆/禾间作系统内优势主导作物禾本科牧草燕麦截获和利用更多有效光能,提高其光合RuBPCase 活性和光合速率,增强燕麦光合作用为碳、氮代谢提供能量保障,而受到燕麦遮阴的紫花苜蓿虽提高了中下层对光能的截获和利用[34-35],但获取的弱光会降低其碳同化酶活性,最终导致其光合能力降低,不利于其碳代谢及物质积累。同时,RuBPCase 活性的提升也会加速燕麦合成碳水化合物,使得燕麦在间作下的碳水化合物含量及积累量显著高于单作。另外,本研究中氮代谢特征表现为,间作能显著提高燕麦的NR、NiR、GS、GOGAT 氮代谢酶活性,加速其氮代谢运转,进而提高其体内氮积累量和蛋白含量,改善燕麦品质,而间作不利于紫花苜蓿氮代谢产物及氮代谢酶活性的增加,在袁洋等[36]玉米/花生(Arachis hypogaea)间作和宋艳霞等[37]玉米套作大豆的研究中也有类似发现,可能是由于间作提升了燕麦旗叶的有效光能利用和光合速率,加速了燕麦功能叶片NR转录翻译及GS表达以加强其对NO3
--N 的还原和NH4+-N 同化[32],并 且GS 与GOGAT,NR 与NiR 之间皆具有显著的耦合关系,燕麦氮代谢水平的提升也会刺激其根系竞争吸收紫花苜蓿根区氮养分,增强紫花苜蓿结瘤固氮以促进系统内氮营养的互补利用,进而显著增加燕麦氮代谢酶活性和氮代谢产物积累。此外,本团队前期研究已证实,豆/禾间作中随着地上及地下完全互作,禾本科牧草的氮代谢酶活性以及氮代谢产物积累会加强[7]。可见,作物地下部的根系活动与其地上部光合作用、碳氮代谢及物质转运等过程是一个不可分割的有机整体,合理搭配的豆/禾间作系统会更有利于发挥其地上、地下互作正效应,加强系统内禾本科牧草光合作用,增强其碳、氮代谢酶活性和加速代谢产物形成,进而提高禾本科牧草的生物量和蛋白总量。
间作系统内作物地上部对光热资源的利用和竞争,地下部根系在空间上的交叉和重叠以及对地下部水分及养分资源的竞争和互补的协调统一是形成间作优势效应的关键[38]。在本研究紫花苜蓿/燕麦间作体系中,生物量与光合气体交换参数、氮代谢酶活性、根系特性呈正相关;蛋白总量与光合气体交换参数呈极显著正相关,与4个氮代谢酶活性、根表面积等呈显著正相关,说明无论紫花苜蓿还是燕麦,地上部与地下部对资源的获取和分配均存在较强相关性,根系及碳、氮代谢的改善是其获得较高生物量和蛋白总量的重要基础。此外,作物的根系活动与碳、氮代谢过程是一个相互协调且不可分割的有机整体[9],本研究的紫花苜蓿/燕麦间作中,根长与净光合速率和4个氮代谢酶活性呈显著正相关;根表面积、根平均直径与NiR、GOGAT 活性呈极显著正相关,充分说明改善间作系统内作物根长和根表面积等根系形态参数会对碳、氮代谢起显著促进作用,进而形成一个完整的良性循环体系,汪顺义等[39]对甘薯(Ipomoea batatas)和王佳等[40]对小麦的研究中也证实了根系与碳、氮代谢变化具有协调一致性。由此可见,在紫花苜蓿/燕麦间作体系内,无论紫花苜蓿还是燕麦,其根长、根表面积和根体积等根系特性直接决定了碳、氮代谢运转的强度和质量,协调好根系与碳、氮代谢连续体之间的相互关系是提高燕麦和紫花苜蓿生物量和品质的重要基础。
4 结论
与单作相比,紫花苜蓿与燕麦间作显著改善了燕麦的根系形态参数,提高了其光合作用及碳、氮代谢酶活性,促进了其体内碳水化合物及蛋白质积累以改善其生物量和品质,而降低了紫花苜蓿根表面积及碳、氮代谢水平,不利于其代谢产物及生物量的积累;随着种植年份的增加,无论单作还是间作下,紫花苜蓿根系特性均随之增加,连续间作促进了系统内紫花苜蓿总根长的拓展,逐渐减弱了燕麦对紫花苜蓿根表面积和根体积的抑制,且燕麦在第2 和第3 年间作下的根系形态参数均高于单作。
在紫花苜蓿/燕麦间作体系内,总根长、根表面积和根体积对碳、氮代谢起显著促进作用,根系特征与碳、氮代谢特性的协调一致可有效地提高间作体系的生物量和蛋白总量。
猜你喜欢
经济林研究(2022年4期)2023-01-12
广西林业科学(2022年2期)2022-05-09
小小说月刊·下半月(2021年8期)2021-09-05
河北果树(2020年4期)2020-11-26
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
动漫界·幼教365(大班)(2018年3期)2018-05-14
干旱地区农业研究(2017年5期)2017-12-18
山东农业科学(2017年4期)2017-05-18
新疆农垦科技(2016年2期)2016-08-21
