
质体ACCase Trp-1999-Leu 突变对小麦田菵草生长适合度的影响
2024-03-22 01:08白霜吕庆浩李怡慧刘国平罗小勇李凌绪
草业学报 2024年3期
白霜,吕庆浩,李怡慧,刘国平,罗小勇,李凌绪*
(1. 青岛农业大学植物医学学院,山东省植物病虫害绿色防控工程研究中心,山东 青岛 266109;2. 安丘市景芝镇农业综合服务中心,山东 潍坊 262119)
菵草(Beckmannia syzigachne)是我国常见的禾本科二倍体杂草,喜湿,在长江中下游地区、山东省微山湖地区的稻茬麦田发生尤为严重[1]。精噁唑禾草灵是小麦田苗后选择性防除禾本科杂草的主要药剂之一[2],通过抑制禾本科杂草质体型乙酰辅酶A 羧化酶(acetyl co-enzyme A carboxylase, ACCase)的活性影响脂肪酸的合成,导致植物死亡[3-4]。但是,越来越多的杂草已被报道对ACCase 抑制剂产生抗药性,这种抗性往往与靶标基因变异有关。目前,已报道杂草ACCase CT 区域突变较多的氨基酸残基位点是2041 和1781 位,最少的是2088 位。不同位点氨基酸变异的形式也存在多样化,例如2041 位异亮氨酸(isoleucine, Ile)突变为天冬酰胺(asparagine, Asn)频率较高,1781 位异亮氨酸(isoleucine, Ile)变为亮氨酸(leucine, Leu)频率较高而变为缬氨酸(valine, Val)、苏氨酸(threonine, Thr)频率较低[5]。实验室前期采集自江苏丹阳的麦田菵草种群JS-26 已明确对ACCase 抑制剂精噁唑禾草灵、唑啉草酯、噁唑酰草胺、烯禾啶、氰氟草酯等均产生不同程度抗性,靶标酶质体型ACCase 1999 位点氨基酸变异(Trp 突变为Leu)是其对上述除草剂产生抗性的重要原因[6]。抗药性杂草一旦脱离除草剂的选择压力,其适应生存环境的能力可能会降低,即所谓的适合度代价[7-9]。菵草抗性机制的明确对其治理策略提供了理论基础,但是在无除草剂选择压力下,抗性菵草在田间如何演化,群落如何演替等一系列与生产密切相关的问题还亟待解决,而对抗性菵草适合度的深入研究将有效解释此类问题。因此,明确抗药性杂草在无除草剂选择压力下的适合度变化对抗性杂草的治理意义重大[4]。目前,质体型菵草ACCase Ile-1781-Leu、Trp-2027-Cys、Tle-2041-Asn、Asp-2078-Gly、Gly-2096-Ala 突变型的适合度代价已被明确,例如Ile-1781-Leu 突变能提高菵草的相对生长速率及净同化率,但对该基因型菵草的竞争力无显著影响;Trp-2027-Cys 突变能降低菵草种子产量但对营养期生长无影响;Tle-2041-Asn 和Gly-2096-Ala 突变则对菵草营养期生长及竞争能力均无显著影响;Asp-2078-Gly突变可降低菵草的生长速率及净同化率,同时还会影响菵草的植株生物量、株高及结种量[7]。但是Trp-1999-Leu突变对菵草的生长特性、繁殖力和竞争力等适合度变化的影响未见报道。
对抗性菵草适合度的分析需要注意选择与抗性菵草具有相似遗传背景的材料作对照,因为不同遗传背景下的种群往往表现出不同的生长特性。本研究的两种基因型菵草均分离自JS-26 种群,参照之前的衍生酶切扩增序列多态性技术(derived cleaved amplified polymorphic sequences, dCAPS)方法[10],通过错配引入Hind III 的酶切位点(AAGCTT)扩增JS-26 菵草质体Accase CT 区片段,分析菵草质体ACCase 的单核苷酸多态性(single nucleotide polymorphism, SNP)(G/T)。扩增产物经Hind III 内切酶消化后电泳检测,野生型种群只有一条390 bp 条带,纯合突变型种群有一条可见的349 bp 条带和一条不可见的41 bp 条带,杂合型同时存在390 和349 bp 两条条带。根据dCAPS 分析结果,纯合野生型和突变型种群被分别置于不同温室培养,收取纯合基因型菵草种子[11]。本研究以不存在遗传背景差异的ACCase 1999 位突变纯合体(Trp/Trp, RR)和1999 位纯合野生型(Leu/Leu, SS)为研究对象,研究1999 位点Trp 突变为Leu 对菵草的生长特性及生态适合度的影响,以期为该基因型菵草的进化及治理提供理论指导。
1 材料与方法
1.1 供试材料
抗性菵草种群JS-26 采集自江苏省丹阳市,其质体ACCase CT 区域1999 位氨基酸色氨酸(tryptophane, Trp)突变为Leu,通过dCPAS 法[10]从JS-26 种群中分离得到纯化的RR 及SS 基因型。
1.2 种子萌发及培养
将菵草种子放在铺有两层滤纸的9 cm 培养皿中,加入5 mL 清水,将培养皿置于光照培养箱中催芽,每天更换一次水。培养箱温度为白天20 ℃/晚上10 ℃,光照周期为12 h 昼夜交替。当幼苗长至1~2 cm 时,根据试验要求将其移栽至相应大小的花盆中,花盆均为直径22.5 cm×高15 cm。填盆基质为取自青岛农业大学牡丹园未施用过除草剂的壤土,过2 mm 筛。除田间竞争试验外,室内培养条件均为白天温度(25±5) ℃,夜间(15±5) ℃。
1.3 早期生长特性对比
催芽后的RR 和SS 基因型菵草移栽于花盆中培养。每盆移栽5 株,每个基因型20 盆。分别于移栽后第10 和15 天测量株高及地上生物量,整体试验重复两次。
1.4 营养生长期生长特性对比
萌发后的菵草移栽至花盆中培养,菵草叶面积和地上干重于移栽后的第25 和53 天进行测量和记录,每个基因型每次测量4 盆,取平均值。比较两种基因型菵草相对生长速率(relative growth rate, RGR)[12]、净同化率(net assimilation rate, NAR)和叶面积比(leaf area ratio, LAR)[13]的差异,整体试验重复两次。计算公式如下:
式中:t为时间,W为干物质量,w1、w2分别为t1、t2时间的干物质量。
式中:L为叶面积,L1、L2分别为t1、t2时间的叶面积。
1.5 成熟期生长特性对比
将RR 和SS 幼苗移栽入花盆中。在日光温室中培养,按需定量浇水,自然光照,随机摆放花盆并定期打乱,花盆大小、植株培养基质、培养条件如1.2 所述。观察记录菵草的分蘖数,评估每株植物的生殖生长情况。
从第一次成熟开始依次收集RR 和SS 植株的花序,将每一株植株的花序脱粒,分离种子,并对总种子质量进行量化。每棵菵草的种子产量(Sn)为:
式中:TSW为每株菵草产生的总种子重量,SW为每株菵草100 粒种子的重量。
根据Reekie 等[14]的方法估算每个基因型植物中分配给种子的资源的相对比例[即繁殖力(reproduction effort, RE)]。计算公式如下:
式中:Sb表示每株菵草所有种子的生物量,Tb表示每株植物产生的总生物量(根、茎、叶、花序和种子)。
1.6 室内条件下菵草的种间竞争力对比
在室内采用盆栽法比较两种基因型菵草和小麦(Triticum aestivum)的种间竞争力。把出芽的菵草移栽入直径24 cm×高12 cm 的花盆中,每盆4 株菵草。在种植菵草的花盆中植入小麦(宁麦13),使得盆中小麦密度依次为0、40、80、160、320、640 株·m-2。花盆随机摆放并每2 d 随机调整位置。在小麦成熟前,测定菵草的结种量、地上生物量[6],每处理包含4 次重复。
数据处理参照Vila-aiub 等[15]的方法,利用SigmaPlot 采用双曲非线性模型计算评价两种基因型菵草在室内盆栽条件下与小麦的竞争力,计算公式如下:
旅游作为消费者行为自古至今普遍存在,但医疗旅游、自然医药旅游、健康旅游等旅游的具体形式又因社会制度、发展阶段、经济水平、文化信仰等不同而存在巨大差异。
式中:x表示小麦密度,y表示菵草种子产量,a表示无小麦时菵草种子产量,b表示斜率。曲线坡度越大,表示与小麦的竞争力越弱。
1.7 田间条件下菵草的种间竞争力对比
为了验证两种基因型菵草和小麦的种间竞争力,分别于2019 和2020 年开展田间竞争试验,试验地分别位于山东省莱西市和青岛市即墨区。田间小麦品种均为济麦20,小麦行间距为18 cm,种植小麦后10 d,在小麦行间移栽三叶期菵草,菵草株间距为18 cm。小区设为1.5 m×1.2 m,每小区移栽20 株菵草,采用随机区组设计,每基因型移栽4个小区作为4 次重复,共计8个小区[16]。整个生长周期人工除草,田间管理方式按照当地常规操作进行。次年菵草成熟后收集其种子,测定种子产量,并测定其株高、分蘖数和地上生物量。
1.8 数据处理
试验结果均以平均值±标准误表示,使用Excel 2010 软件对数据进行统计、处理,采用IBM SPSS Statistics 26 软件对数据进行独立样本t检验(α=0.05),并通过SigmaPlot 12.5 进行绘图。
2 结果与分析
2.1 早期生长特性分析
为了明确两种基因型菵草的早期生长差异,本研究测定了菵草移栽第10 和15 天时的株高、地上生物量。结果表明,在菵草移栽第10 天时,抗性菵草RR 与敏感型菵草SS 的生物量和株高均无显著差异;但是在第15 天时,RR 型菵草的生物量和株高分别为2.05 g·株-1和86.65 mm,SS 型菵草的生物量和株高分别为2.73 g·株-1和97.92 mm,RR 植株的生物量和株高分别显著低于SS 植株24.9%和11.5%(表1),由此说明早期生长阶段抗性菵草RR的生长速率稍慢于SS 型菵草。
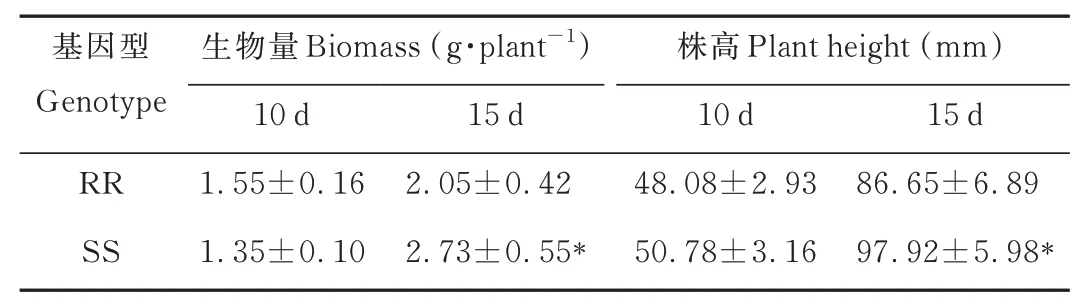
表1 不同基因型菵草早期生长阶段植株生物量和株高对比Table 1 Comparison of biomass and plant height of different B.syzigachnegenotypes in early growing stage
2.2 营养生长期生长特性对比
为明确两种基因型菵草在营养生长阶段的生长差异,本研究测定了两种基因型菵草在移栽后第25 和53 天时的地上生物量及叶面积,并分别测定了二者在移栽25~53 d 这一阶段的相对生长速率、净同化率和叶面积比。结果表明,两种基因型菵草在25 和53 d 时地上生物量及叶面积均无显著差异;然而在这一生长阶段,RR 型菵草表现出了较低的相对生长速率(RGR)和净同化率(NAR),分别比SS 型低18.2%和28.6%,但RR 型却具有更高的叶面积比(LAR),比SS 型高14.0%(表2),这可能是导致二者在53 d 时具有相似地上生物量的原因。而且在这一阶段,RR 和SS 型较早期生长差异有所减小。
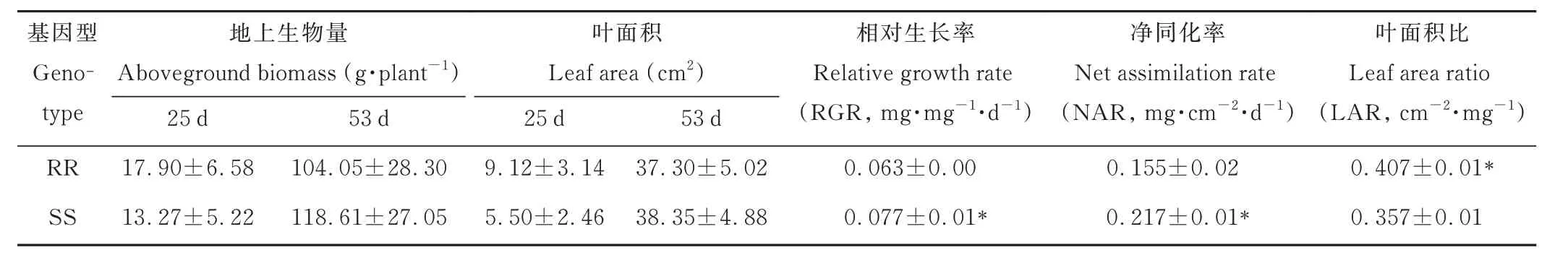
表2 不同基因型菵草营养生长期阶段的生长参数Table 2 Plant growth traits of different B. syzigachnegenotypesat the vegetative stage
2.3 成熟期生长特性对比
为了明确两种基因型菵草在成熟期时的生长特性差异,在温室条件下比较了两种基因型菵草在成熟期时的分蘖数、平均植株总生物量、平均植株种子数、平均植株总种子重量和繁殖力。结果表明,两种基因型菵草在植株成熟期时平均植株总生物量、分蘖数、平均植株种子数均无显著差异(表3)。但是RR 型菵草在平均植株总种子重量、繁殖力方面表现出劣势,与SS 型植株相比,RR 基因型平均植株总种子重量减少11.4%,繁殖力降低13.1%(表3)。

表3 不同基因型菵草植株繁殖力对比Table 3 Comparison of reproduction effort between different B. syzigachnegenotypes
2.4 室内条件下菵草与小麦的竞争力对比
为了明确两种基因型菵草与小麦的种间竞争力差异,在室内条件下,利用盆栽试验比较了两种基因型菵草对不同密度的小麦的竞争反应。结果表明,随着小麦密度的增加,两种基因型菵草的长势、结种量均受到抑制,但是RR 基因型菵草表现出更弱的竞争能力。在40、80、160、320、640 株·m-2的小麦密度下,RR 型菵草的地上生物量比SS 型分别降低了29.6%、35.2%、41.5%、46.6%、42.4%(图1);平均种子产量比SS 型分别降低了46.6%、51.5%、48.2%、64.9%、55.9%(图2)。

图1 RR 和SS 基因型菵草在不同小麦密度下的地上生物量Fig. 1 Aboveground biomass of RR and SS genotypes B.syzigachne at different wheat densities
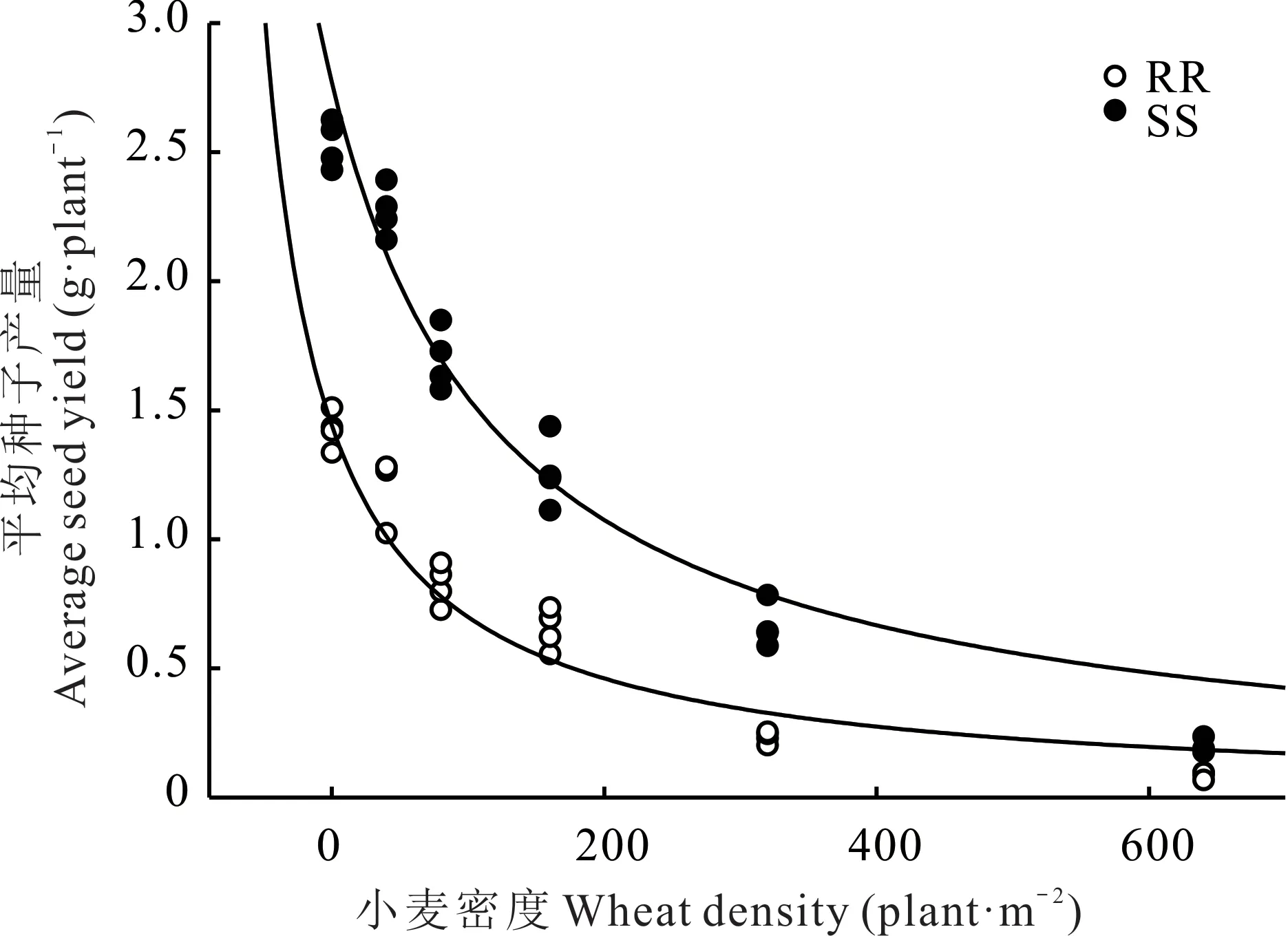
图2 RR 和SS 基因型菵草在不同小麦密度下的平均种子产量Fig. 2 Average seed yield of RR and SS genotypes B.syzigachne at different wheat densities
2.5 田间条件下菵草的种间竞争力对比
为了进一步验证室内两种基因型菵草与小麦竞争力差异的结果,在田间条件下比较了在小麦竞争下的两种基因型菵草的株高、分蘖数、地上生物量和种子产量。田间试验分两年两地进行,菵草生长期间未施用任何除草剂。两地试验结果均表明,经过8个月的生长,两种基因型菵草株高、分蘖数差异不显著;但RR 基因型菵草与SS 型相比其地上生物量和种子产量分别减少了17.6%、21.1%(即墨),30.2%、34.7%(莱西)(表4)。这表明RR 基因型菵草在地上生物量和种子产量方面表现出一定的适合度代价,这与室内盆栽试验结果一致。
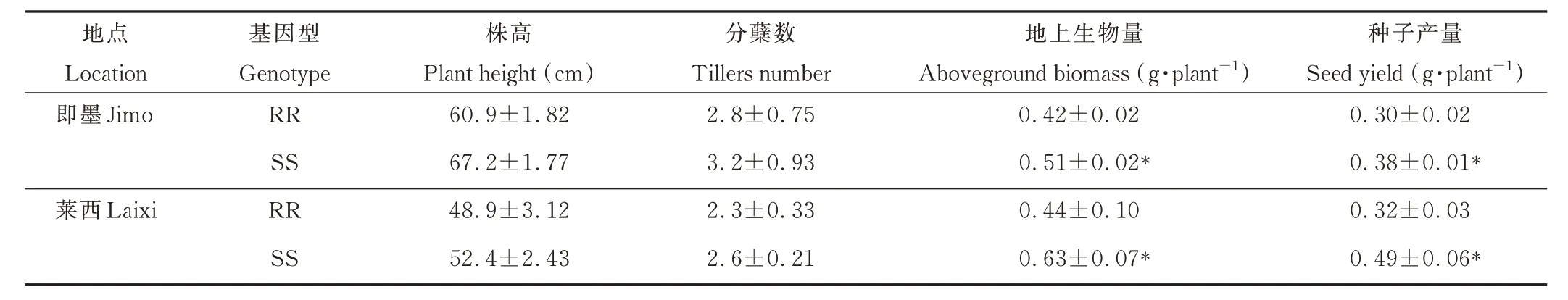
表4 不同基因型菵草在田间竞争压力下的生长参数Table 4 Growth traits under competitive pressure in the field of different B. syzigachnegenotypes
3 讨论
衡量个体或种群在不同环境下的适应程度,或者是个体对群体基因库的相对贡献程度即所谓的适合度,也是指特定基因型个体在同种生物群体中生存和繁衍后代的能力[17-19]。具有抗性基因存在的杂草在无除草剂选择压力下的适合度变化对预测其进化动态至关重要,也可以为利用其产生的适合度代价进行抗性杂草的治理提供参考。对抗性杂草适合度的分析需要选择遗传背景相似的试验材料作对照,本研究的两种基因型菵草不存在遗传背景差异。
本研究中,在菵草早期生长阶段,RR 基因型菵草生物量积累效率较低,仅在种植后的10~15 d,RR 型菵草的地上生物量和株高分别比SS 低24.9% 和11.5%(表1)。含有5-烯醇式丙酮酸-3-磷酸酯合成酶(5-enolpyruvylshikimate-3-phosphase synthase, EPSPS)双位点突变(Thr-102-Ile/Pro-106-Ser)的草甘膦抗性牛筋草(Eleusine indica)在早期生长阶段也表现出了较慢的生长速率[20],这说明在没有除草剂的选择压力下,ACCase Trp-1999-Leu 变异可能影响植株在早期生长阶段的生物量积累效率。在菵草营养生长阶段,RR 基因型菵草的相对生长速率和净同化率均低于SS 型(表2),这表明RR 型在捕获光、吸收二氧化碳或储存光同化物方面弱于SS植株[21]。但是RR 型具有较高的叶面积比,这也有可能是造成RR 和SS 在53 d 时具有相近生物量积累的原因。在抗ACCase 抑制剂类除草剂的硬直黑麦草(Lolium rigidum)中,ACCase Asp-2078-Gly 突变会导致其营养生长阶段RGR、NAR 降低,但是对LAR 没有明显影响,最终抗性植株表现出较低的生物量积累[22]。在抗ACCase 抑制剂类除草剂的菵草中,Asp-2078-Gly 突变导致菵草RGR、NAR、LAR 均有所降低,Ile-2041-Asn、Gly-2096-Ala突变则对菵草的RGR、NAR 和LAR 没有显著影响[21]。在抗性棒头草(Polypogon fugax)和大穗看麦娘(Hordeum glaucum)中,ACCase Ile-2041-Asn 突变同样没有影响植株的营养生长[23-24]。但是对乙酰乳酸合成酶(acetolactate synthase, ALS)抑制剂产生抗性的播娘蒿(Descurainia sophia),Pro-197-Leu、Pro-197-His、Pro-197-Thr、Asp-376-Glu、Trp-574-Leu 等基因突变可显著降低播娘蒿的生物量积累[25]。基于以上案例分析,相同基因的相同位点突变会因杂草种类或种群不同表现出不同的生长效应,同一种群杂草也会因相同基因的不同位点突变或同一位点的不同突变形式表现出不同的生长状态,可见基因突变对杂草生长的影响受杂草种类、种群不同而呈现多效性。
本研究发现Trp-1999-Leu 纯合突变型菵草(RR 型)和纯合野生型菵草(SS 型)在成熟期的分蘖数、平均植株总生物量和种子数量未见显著差异,但是Trp-1999-Leu 纯合突变型菵草平均植株总种子重量和繁殖力显著低于野生型菵草,这说明ACCase Trp-1999-Leu 基因突变使菵草在生殖生长方面产生明显的适合度代价。其他抗性杂草因除草剂靶标基因突变导致种子产量适合度降低也有类似的报道。例如对比抗ACCase 抑制剂类除草剂的硬直黑麦草ACCase Ile-1781-Leu 突变型和野生型的繁殖力发现,抗性突变个体产生的种子千粒重要低于野生型个体[26]。在ALS Trp-574-Leu 突变导致的抗ALS 抑制剂的早熟禾(Poa annua)中也发现了这种适合度代价,该突变导致抗性植株种子产量降低、分蘖减少[27]。而ACCase Ile-2041-Asn 突变则不会影响棒头草的繁殖力[24]。这说明,除草剂靶标基因抗性突变对抗性植株繁殖力的适合度影响是多样的。但是靶标基因抗性突变使植株产生繁殖力的适合度代价对杂草治理十分有利,如若发现靶标基因突变降低抗性种群繁殖力的杂草种群,不施用除草剂或许更有助于抗性杂草的治理。
本研究同样比较了Trp-1999-Leu 纯合突变型菵草和纯合野生型菵草与小麦竞争能力的差异。分别在室内和田间条件下展开试验。室内竞争试验表明,RR 和SS 型菵草的结种量和地上生物量均随小麦密度增加而降低,而RR 型菵草结种量和地上生物量,均显著低于SS 型菵草。田间试验与室内结果基本一致:Trp-1999-Leu 突变对菵草植株的株高和分蘖数均影响不大,但结种量和地上生物量均受到显著影响,在即墨和莱西地块均得到了一致的试验结果(表4)。因此,ACCase Trp-1999-Leu 突变可降低菵草与小麦的竞争力,尤其在繁殖和生物量积累方面使菵草表现出明显的适合度代价。本试验结果可说明,在有小麦竞争的条件下,抗性菵草在田间传播、扩散能力更弱。相似的结果在其他杂草中也有报道,例如ALS Ala-122-Asn 突变的抗性稗草(Echinochloa crus-galli)在存在种间竞争和非竞争条件下抗性植株均发育迟缓[28]。ACCase Trp-2027-Cys 突变型菵草结种量减少;Asp-2078-Gly 突变型菵草的生物量、结种量均显著降低[7]。综上说明,除草剂靶标基因的变异或许会降低某些杂草的种间竞争力。这种适合度代价对抗性杂草的治理十分有利,竞争力降低的抗性杂草在无除草剂的选择压力下,种群自然进化可能朝着减少的趋势发展。如若抗性杂草存在靶标基因变异但生长未受影响,这无疑会加大抗性杂草的治理难度。
4 结论
本研究以质体ACCase 等位基因纯合突变引起抗性的菵草及纯合野生型菵草为研究对象,探讨了Trp-1999-Leu 突变对菵草生长、繁殖及竞争力的影响。结果表明该基因突变可减缓菵草在早期阶段的营养生长。但是在生长的中期,RR 与SS 地上生物量积累差异并不显著。在存在小麦种间竞争条件下,ACCase Trp-1999-Leu 基因突变虽不影响菵草的株高及分蘖,但会显著降低菵草的繁殖力和地上生物量,产生明显的竞争力适合度代价,这种代价在无除草剂竞争压力下可降低该基因型菵草的治理难度。
猜你喜欢
科教新报(2022年22期)2022-07-02
空间科学学报(2021年1期)2021-05-22
今日农业(2019年15期)2019-01-03
环境保护与循环经济(2017年5期)2018-01-22
现代园艺(2017年19期)2018-01-19
中国果菜(2016年9期)2016-03-01
中国蔬菜(2015年9期)2015-12-21
营销界(2015年23期)2015-02-28
现代农业(2015年5期)2015-02-28
中国质量与标准导报(2013年8期)2013-03-11
