
马铃薯低温糖化形成机理及其调控因素研究进展
2024-03-28 10:15崔广璨李清清王庆国刘腾飞
食品研究与开发 2024年6期
崔广璨,李清清,王庆国,刘腾飞
(山东农业大学 食品科学与工程学院,山东 泰安 271018)
马铃薯(SolanumtuberosumL.)是世界第四大作物,其营养丰富,是多种加工食品的重要原料。由马铃薯直接加工的油炸产品,如油炸薯条和薯片等在马铃薯加工产业中占比达70%以上[1]。在马铃薯加工工业中,为了抑制块茎失水、发芽、腐烂及病害等采后问题,马铃薯块茎通常在低温条件下进行贮藏[2],但低温会促进马铃薯块茎中的淀粉降解为蔗糖,蔗糖进一步转运至液泡,在液泡酸性转化酶的作用下,分解为还原性的葡萄糖和果糖,导致还原性糖积累,这种现象称为低温糖化。对于马铃薯加工工业而言,低温糖化是制约原料薯周年供应的重要因素。糖化后的马铃薯块茎在后期高温油炸过程中易发生美拉德反应[3],即还原性糖和游离氨基酸发生非酶促反应,产生褐色物质以及有毒的潜在致癌物质丙烯酰胺[4],不仅使得油炸产品颜色变褐或变黑,口味发苦,也对人体健康造成危害,且贮藏温度越低,低温糖化现象越严重[5]。低温糖化严重的马铃薯中还原性糖含量高,高温油炸时更易产生有毒物质丙烯酰胺、杂环胺等[6]。在工业生产中,因低温糖化引起的块茎淀粉向还原糖的过量转化,是造成原料薯损失的重要因素[7]。对于马铃薯加工工业而言,低温糖化问题非常重要,需引起广泛关注。
关于针对低温糖化过程中淀粉与糖的转化机理及生物技术育种应用等方面的研究较多,但缺乏对这些相关研究进展的系统性总结和凝练。本文旨在阐明马铃薯低温糖化的危害、发生的机制及影响因素,使人们合理规避马铃薯低温糖化造成的不利影响,为油炸马铃薯加工产业的发展提供参考。
1 马铃薯的碳水化合物组分对块茎低温糖化和油炸加工品质影响
1.1 块茎淀粉结构对马铃薯低温糖化的影响
1.1.1 直链淀粉与支链淀粉
马铃薯中淀粉含量丰富,淀粉由于结构的差异可分为直链淀粉与支链淀粉[8]。直链淀粉分子量较小,聚合度为300~5 000,支链淀粉分子量较大,聚合度为9 000~16 000,一般情况下,支链淀粉比直链淀粉大几十倍,在某些特殊情况下会大几百倍。当直链淀粉与支链淀粉两者混合的比例不同时,会导致淀粉的性质不同,进而对马铃薯贮藏加工产生不同影响。可知,直链淀粉与支链淀粉的比例对淀粉的物理化学和功能特性产生决定性作用,对物理范畴修饰和化学范畴修饰的敏感性有一定影响作用[9]。
谷物淀粉中直链淀粉含量为15%~25%,马铃薯淀粉中直链淀粉含量略低,为10%~20%[10]。马铃薯中支链淀粉含量较高,在淀粉向糖转化过程中影响作用较大。支链淀粉形成的晶型不稳定,使得酶更容易作用在淀粉颗粒上。在低温条件下,支链淀粉含量高的马铃薯淀粉向糖的转化速度更快[11]。Jansky 等[12]发现,抗低温糖化马铃薯品种的直链淀粉含量大于不抗低温糖化品种。综合来看,影响淀粉与马铃薯转化的关键因素之一为直链淀粉与支链淀粉的比值,比值越大,越能够抑制淀粉向糖的转化,比值越小,越能够促进淀粉向糖的转化。在油炸马铃薯产业实际生产应用中应选择直链淀粉含量较高的品种,抑制淀粉向糖的转化,以此获得优良的油炸加工品质。
1.1.2 淀粉颗粒
大量淀粉分子聚合在一起形成淀粉颗粒,自然光下肉眼观察淀粉颗粒呈白色,有轻微光泽。不同的物种中淀粉颗粒的形状不同,有圆形、卵形、不规则形等。马铃薯淀粉颗粒形状为卵圆形,相较于稻米淀粉颗粒、玉米淀粉颗粒等,马铃薯淀粉颗粒较大,粒径为25~100µm。吕文河等[13]发现,淀粉颗粒在块茎中的分布呈现一定的规律,整体来说,在靠近块茎皮层部位相对块茎内层部位淀粉颗粒密度大。淀粉在马铃薯块茎中具有组织特异性,故在研究其性质时要求均匀取样,两侧、脐部、顶部、髓部均要等量的涵盖在研究样本中,同时不应该忽略个体差异,在测量时应选择大量的样本。淀粉粒大小对淀粉向糖的转化也有一定的作用,淀粉在贮藏过程中,小淀粉粒比例减小,大淀粉粒比例增大[14]。小淀粉粒比大淀粉粒降解更快,可能是由于淀粉粒的相对表面积较大,与酶反应的速度更快。
1.2 还原糖含量对块茎油炸加工品质的影响
还原糖类是马铃薯高温油炸过程中产生丙烯酰胺的重要底物。因此,要加工出优质油炸马铃薯产品,必须选择还原糖含量适宜的马铃薯块茎。一般要求生产中用于油炸的马铃薯还原糖含量低于0.4%[15]。但在实际生产中马铃薯块茎的还原糖含量存在几种不同情况:1)刚采收时与在不利条件贮存后,还原糖含量均低于工业要求值;2)刚采收时还原糖含量可接受,但在不利条件贮存后,还原糖含量增至不可接受水平;3)刚采收时与在不利条件贮存后,还原糖含量均不可接受[16]。要解决马铃薯块茎中还原糖含量过高的问题,可从育种和外源技术处理两方面入手。针对后两种情况可进行合理育种,研发出更多更优质的第一类马铃薯品种,但育种工作较为复杂,耗费人力物力较多。对于第2 种情况,应使用合理控制技术,使得其在不利条件贮存后还原糖含量仍可接受,以此解决块茎中还原糖含量较高的问题。Mccann 等[17]使用多元回归法分析了块茎油炸加工产品色泽与块茎成分的关系,结果表明,游离氨基酸(如天冬酰胺)含量和干物质百分比对薯片颜色的变化没有显著影响,还原糖(果糖或葡萄糖)含量是影响薯片颜色变化的主要原因;且在分析中同时存在葡萄糖和果糖时,对颜色变化影响的贡献值不同,一半品种中葡萄糖贡献值较大,另一半品种中果糖贡献值较大,而蔗糖浓度对于薯片颜色变化的影响较小。
2 马铃薯低温糖化发生的机理
2.1 淀粉体膜的稳定性影响马铃薯低温糖化
淀粉体膜一般含有半乳糖脂和磷脂。Gupta 等[18]认为,马铃薯在发生低温糖化的过程中淀粉体膜整体结构基本保持完整。但O'Donoghue 等[19]认为,在低温糖化过程中淀粉体膜结构发生一定变化,即脂膜流动性降低,膜通透性改变。低温处理下,发现低温糖化敏感型马铃薯淀粉体膜的衰老程度大于不敏感型马铃薯块茎淀粉体膜,说明低温糖化过程中淀粉向糖的转化也与淀粉体膜稳定性相关。在遇到低温时所有生物膜的流动性均会降低,但具体淀粉体膜结构是否出现大的改变,可能与温度阈值相关。膜结构的破裂或变化会在低于特定温度阈值时发生,这个阈值因品种而异。每种品种的低温糖化温度阈值不同,淀粉体膜结构变化的阈值温度与低温糖化阈值温度之间的差异也因品种而有所不同。未来,可进行大量马铃薯品种的低温糖化阈值温度和淀粉体膜结构改变阈值温度测量比较,以确定不同品种间是否存在共性,进而明确淀粉体膜结构的变化对马铃薯低温糖化过程中还原糖累积的影响。通过对淀粉体膜结构改变阈值温度的研究也可为探索淀粉体膜结构与低温糖化过程之间的关系提供基础,进一步揭示低温糖化的机理和影响因素。
2.2 低温糖化代谢途径以及相关酶及其调控蛋白
马铃薯块茎的低温糖化与块茎碳水化合物代谢途径紧密相关。淀粉在淀粉磷酸化酶、葡萄糖磷酸变位酶等作用下转化为6-磷酸葡萄糖(glucose-6-phosphate,G-6-P)。在适宜条件下,G-6-P 也会转化为淀粉,其中关键限速酶为腺苷二磷酸葡萄糖焦磷酸化酶(adenosine diphosphoglucose pyrophosphprylase,AGPase)。G-6-P 是多个代谢途径中的关键物质,在细胞质中,经碳水化合物循环,使其转化为蔗糖,其中关键限速酶为尿苷二磷酸葡萄糖焦磷酸化酶(udp-glucose pyrophosphorylase,UGPase)。蔗糖经液泡内的酸性转化酶作用后进一步降解为葡萄糖和果糖,在此过程中位于细胞质中的碱性转化酶和中性转化酶对蔗糖也会起到一定的降解作用[20],但整体作用不大。对低温贮藏一段时间的马铃薯块茎进行代谢分析显示,液泡内酸性转化酶活性是影响还原糖形成的关键因素[21]。
对于淀粉与糖的转化过程有几个比较关键的酶,如淀粉酶、AGPase、UGPase、液泡酸性转化酶(vacuolar acid invertase,StvacINV1)等。淀粉酶活性升高,会促进淀粉向糖方向的转化。淀粉酶分为α-淀粉酶和β-淀粉酶,在淀粉向糖的转化过程中,β-淀粉酶起到主要作用[22]。α-淀粉酶(α-amylase,Amy)的功能主要由Amy23基因控制,β-淀粉酶(β-amylase,BAM)的功能主要由StBAM1和StBAM9基因控制[23]。淀粉酶抑制子(amylase inhibitor,SbAI)能通过抑制低温条件下淀粉降解从而减缓淀粉向糖方向的转化,进一步通过酵母双杂交和双分子荧光互补实验证明,SbAI 蛋白与α-淀粉酶StAmy23 及β-淀粉酶StBAM1 和StBAM9 在植物细胞内互作[24]。StAmy23 定位于细胞质,而StBAM1 和StBAM9 分别定位于质体基质和淀粉颗粒[23]。StBAM1具有β-淀粉酶活性,主要通过降解淀粉发挥功能,StBAM9 不具有淀粉酶活性,但能通过与活性淀粉酶StBAM1 互作调控淀粉降解。StAmy23沉默导致糖原含量升高,还原糖含量降低,说明StAmy23 通过降解胞质糖原进行调控。环指蛋白1(ring finger protein 1,SbRFP1)可通过抑制淀粉酶活性和转化酶活性来减缓块茎淀粉降解和还原糖的积累,研究发现,其主要通过抑制β-淀粉酶StBAM1 的活性来减缓淀粉向糖方向的转化,且SbRFP1 具有E3 泛素连接酶活性[25-26]。根据SbRFP1 的泛素连接酶活性可推断该基因通过该途径对淀粉与糖转化过程中的酶活性进行调控,未来可进一步探究SbRFP1 调控的泛素化途径与淀粉与糖转化途径中的关系。
AGPase 活性升高,会促进糖向淀粉方向转化,其为淀粉再合成的限速酶。马铃薯AGPase 为异源四聚体,由2 个小亚基和2 个大亚基组成,AGPase 大亚基只起调节作用,而活性则由小亚基sAGP 控制,通过调控AGPase 的活性可促进还原糖向淀粉方向的转化[27]。成善汉等[28]研究发现,随着AGPase 活性降低,块茎主要贮藏蛋白——马铃薯糖蛋白(patatin)含量随之降低,patatin 是占块茎总蛋白40% 的糖蛋白,目前来看AGPase 活性与含量是否与patatin 存在关系、存在何种关系仍需进一步研究。AGPase 在低温条件下活性后期低于常温,总体趋势随低温贮藏的时间增加而活性降低[21],在研究淀粉与糖的转化过程中该规律可提供一定的参考。Shi 等[29]发现,马铃薯CBF/DREB-III 转录因子亚家族基因StTINY3,该基因能正向调控低温糖化抗性,StTINY3能够正调控AGPase的表达而提高AGPase 活性,同时激活转化酶抑制子(invertase inhibitor,StInvInh2)的表达以抑制液泡酸性转化酶活性,从而抑制低温糖化过程的进行。
UGPase 活性升高,也会促进淀粉向糖方向转化。该酶催化淀粉合成的前体物质1-磷酸-葡萄糖合成尿苷二磷酸葡萄糖,从而促进蔗糖的再合成。有研究对马铃薯进行长时间低温贮藏,结果显示在贮藏期间块茎中的蔗糖含量与UGPase 活性的相关性极强,说明在低温糖化过程中UGPase 对蔗糖合成过程有显著作用[30]。
低温糖化过程中,蔗糖的合成发生在细胞质中,UDP-葡萄糖焦磷酸化酶催化葡萄糖-1-磷酸生成UDP-葡萄糖,是蔗糖合成的速率限制步骤。研究发现,通过RNA 干扰抑制编码马铃薯UGPase 的StUGPase基因,能够明显降低低温贮藏后马铃薯块茎中的还原糖含量,提高油炸加工品质。该研究揭示了StUGPase通过调控低温糖化过程中的蔗糖再合成从而影响还原性糖的累积以及马铃薯加工质量[31]。蔗糖的分解主要在液泡中进行,蔗糖如何进入液泡这一过程一直未得到解释。研究表明,低温糖化过程中蔗糖由细胞质转运至液泡是由液泡膜糖转运蛋白(tonoplast sugar transporter,TST)StTST1 介导的[32],通过抑制StTST1的表达,不仅能抑制低温贮藏后块茎还原性糖的累积,还能促进冷藏块茎中的淀粉合成并抑制淀粉的降解。由此可见,由StTST1 介导的蔗糖向液泡的转运过程是低温糖化过程的关键因子之一。
酸性转化酶是直接催化蔗糖转化为还原糖的关键酶,其活性升高会促进淀粉向糖方向的转化。有研究比较不同温度下、不同转化酶的活性,发现在低温4 ℃条件下酸性、碱性转化酶活性上升均极显著,12 ℃与20 ℃条件下酶活性差异不大;对不同转化酶进行比较,发现酸性转化酶活性明显高于碱性转化酶,说明酸性转化酶在低温贮藏块茎中是主要的转化酶,该酶对低温条件较为敏感。对于调控酸性转化酶的基因也有大量研究,将马铃薯糖蛋白启动子驱动的烟草酸性转化酶抑制子基因全长cDNA 导入鄂马铃薯3 号植株,贮藏后发现酸性液泡转化酶活性明显降低,说明该基因能够抑制该酶的活性[15]。有研究发现,ERF-VII 转录因子花瓣化相关2.3(relative to apetala 2.3,StRAP2.3)[33],它可以直接调节StInvInh2基因的表达正向调控马铃薯低温糖化抗性,这是由于StRAP2.3与StInvInh2启动子区域的ACCGAC 顺式元件结合后激活转录,StInvInh2 是马铃薯内源液泡酸性转化酶StvacINV1 的抑制子,通过限制液泡转化酶活性来抑制低温糖化过程。此外Lin 等[34]研究发现,液泡酸性转化酶StvacINV1 和其抑制子StInvInh2 能与植物的能量感应器复合体蔗糖非发酵1 相关蛋白激酶1(sucrose nonfermenting-1-related protein kinase 1,StSnRK1)的α 和β 亚基形成转化酶活性的精细调控复合体。StSnRK1β 能封阻抑制子StInvInh2 对转化酶的抑制作用,而磷酸化的StSnRK1α 能拮抗StSnRK1β 的封阻,重新实现对转化酶活性的抑制,从而通过StSnRK1α的磷酸化实现对转化酶活性的精细调控。虽然大多数研究表明在淀粉与糖的转化过程中酸性转化酶起到较大的作用,但最近也有研究表明碱性及中性转化酶(neutral invertase,StNI)的作用[20],黄褐色伯班克品种块茎中StNI5转录丰度高,还原糖含量高,推测块茎对低温糖化的敏感性可能是由于NI 对细胞质中蔗糖的分解。在另一夏波蒂品种块茎中StNI4和StNI6转录丰度较高,也表明蔗糖降解需要不同的NI 亚型。对于酸性转化酶和碱性/中性转化酶在马铃薯淀粉与糖转化过程中发挥的作用有许多观点,目前普遍认可酸性转化酶在过程中起到主要作用,碱性/中性转化酶在还原糖调节和过程中也起到一定的作用,但其具体如何进行调节还需要进一步研究确认。影响马铃薯淀粉与糖转化的酶和调节因子如图1 所示。
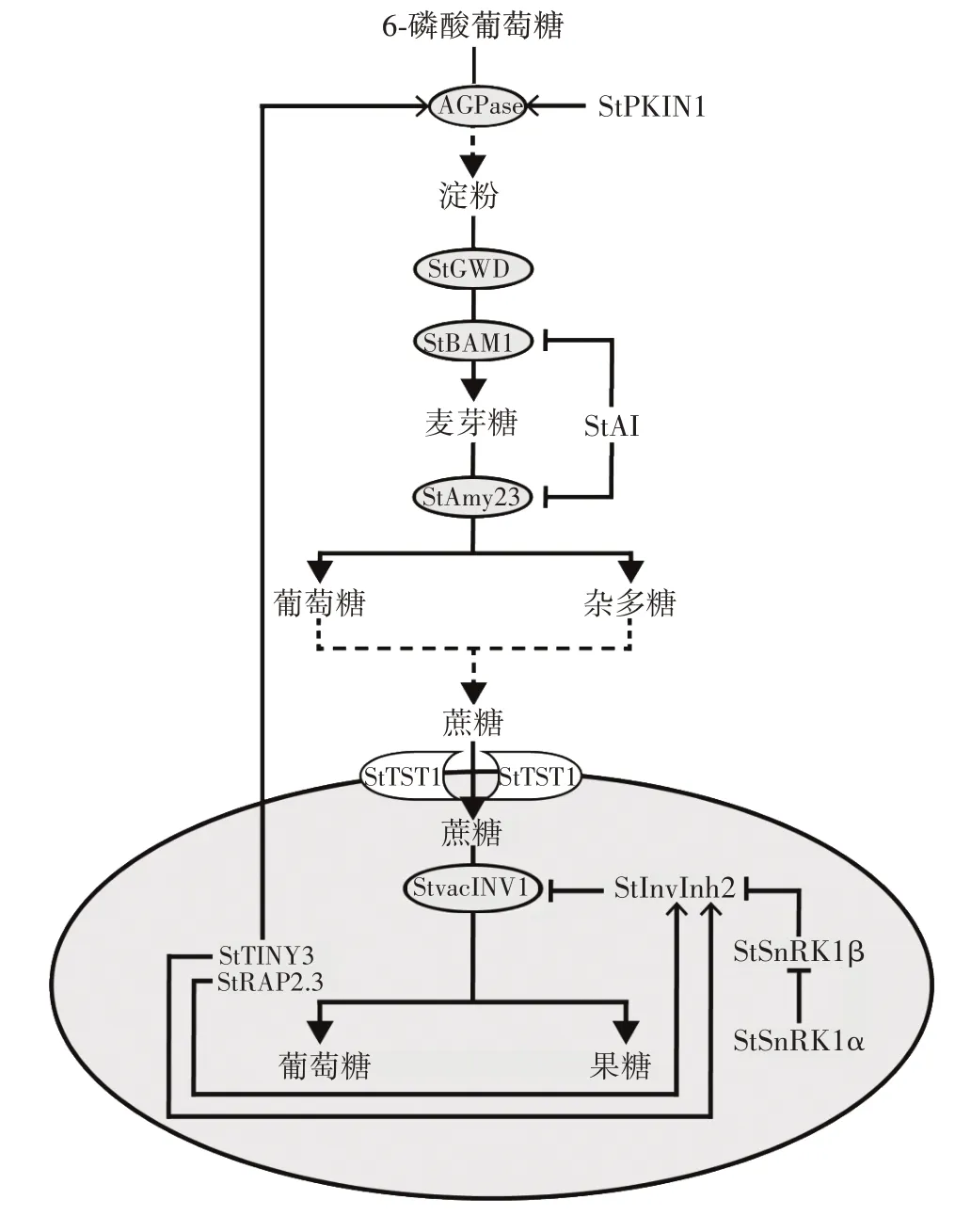
图1 影响马铃薯淀粉与糖转化的酶和调节因子Fig.1 Enzymes and regulatory factors affecting the conversion of starch to sugar in potatoes
由图1 可知,6-磷酸葡萄糖在淀粉合成限速酶AGPase 的作用下最终形成淀粉,其中转录因子StTINY3和StRAP2.3 及激酶StPKIN1 能激活其表达;在淀粉降解过程中,在StGWD1 和StBAM1 的协同作用下淀粉被降解为麦芽糖,麦芽糖在Amy23 的作用下生成葡萄糖和杂多糖,随后合成蔗糖;蔗糖由StTST1 转运至液泡中,进一步被StvacINV1 分解为葡萄糖和果糖,而StInvInh2 能抑制转化酶的活性,同时该抑制作用又受到StSnRK1 复合体的调控[20,23,26-27,29,32-33,35-41]。
2.3 G-6-P 代谢对低温糖化的影响
G-6-P 广泛存在于动植物体和微生物中,参与多种代谢途径。在糖酵解中,葡萄糖被己糖激酶催化生成G-6-P,随后通过磷酸葡萄糖异构酶催化形成6-磷酸果糖(fructose-6-phosphate,F-6-P)。在磷酸戊糖途径中,G-6-P 为第一个反应底物,脱氢生成6-磷酸葡糖酸内酯。低温抑制了糖酵解,导致磷酸己糖积累而转化为蔗糖,糖酵解过程若变慢,淀粉降解后蔗糖积累,在液泡转化酶作用下就会形成还原糖积累[15,42]。可见,G-6-P 是连接了糖酵解、磷酸戊糖途径以及低温糖化的一个关键中间代谢产物。
3 影响马铃薯低温糖化过程中淀粉与糖转化的因素
3.1 品种与基因型
不同品种间以及相同品种不同生长环境下均存在着一定的淀粉与糖转化的差异。将抗低温糖化马铃薯品种和低温糖化敏感的马铃薯品种块茎分别贮藏于20 ℃和4 ℃,在20 ℃贮藏时,未发生糖化,在4 ℃贮藏时,低温糖化敏感块茎的还原糖含量在5~30 d 内迅速上升到较高水平,从鲜重3.5 mg/g 上升到20.7 mg/g,抗低温糖化块茎的还原糖含量只有轻微增加,最大还原糖含量仅为3.4 mg/g。可见,不同品种或基因型在马铃薯淀粉与糖转化过程中影响作用较大[43]。
3.2 栽培条件与时间
马铃薯栽培条件、生长环境、收获期和块茎的成熟度影响淀粉与糖的转化过程。在生长环境中的土壤酸碱度也对淀粉与糖的转化过程有影响[44],生长环境偏酸性,会加速淀粉的分解,推测酸性环境下酸性转化酶的活性会升高,利于将淀粉转化为糖。马铃薯收获期以及块茎成熟度均会造成淀粉含量或形态结构的不同,进而影响整个淀粉糖的转化过程[45]。
3.3 贮藏条件
3.3.1 温度
温度对淀粉与糖的转化影响十分明显,马铃薯块茎尤其对低温较为敏感,在低温条件下极大促进了淀粉向还原糖方向的转化。以不同贮藏温度条件下的两种马铃薯为材料,分析表明在4 ℃贮藏条件下,腺苷二磷酸葡萄糖焦磷酸化酶、尿苷二磷酸葡萄糖焦磷酸化酶和蔗糖合成酶活性与块茎还原糖含量呈显著负相关,酸性转化酶和碱性转化酶活性与块茎还原糖的积累呈显著正相关,由此可见,温度通过改变上述几种酶活性来影响淀粉与糖转化的过程[46]。此外,温度越低时,马铃薯块茎淀粉向糖的转化速度越快[21]。
3.3.2 外源物质
乙烯和脱落酸均会促进马铃薯中的淀粉向糖转化,其促进淀粉降解酶活性,抑制淀粉合成酶活性,而对酸性转化酶活性无影响[47]。赤霉素也会促进淀粉向糖的转化,施用后使还原糖含量大幅增加384.4%,可溶糖增加35.3%,蔗糖增加20.8%,淀粉酶活性升高159%,处理后在低温、常温环境下,酸性液泡转化酶活性发生显著提高[9]。植物激素对马铃薯低温糖化的影响研究较多,但是激素信号对低温糖化的调控机理尚不清楚,今后仍需探索。
S-香芹酮是一种天然存在的化合物,其通过上调颗粒结合淀粉合成酶,会使淀粉合成速率加快,蔗糖分解速率减慢,最终抑制淀粉向糖方向的转化。UV-C 处理也能显著延缓马铃薯块茎中还原糖的积累,其通过降低蔗糖合成速率,抑制蔗糖进一步分解,最终减缓了马铃薯淀粉向糖的快速转化[48]。对于新型糖化抑制剂的开发也是马铃薯的重要研究方向。
3.3.3 气体条件
低氧条件下能够减缓淀粉向糖的转化,其作用在很大程度上是酸性转化酶活性降低的结果,说明低氧条件对酸性转化酶有一定的酶活抑制效果[49],进而对淀粉与糖的转化过程产生影响。另有研究发现,乙烯结合1-甲基环丙烯气调处理可有效减缓马铃薯淀粉向糖的过度转化现象,且在块茎出现萌芽时进行乙烯处理效果最佳[50]。目前主要通过物理、化学、生物方法对低温糖化进行抑制,不同处理方法对果蔬低温糖化的影响如表1 所示。
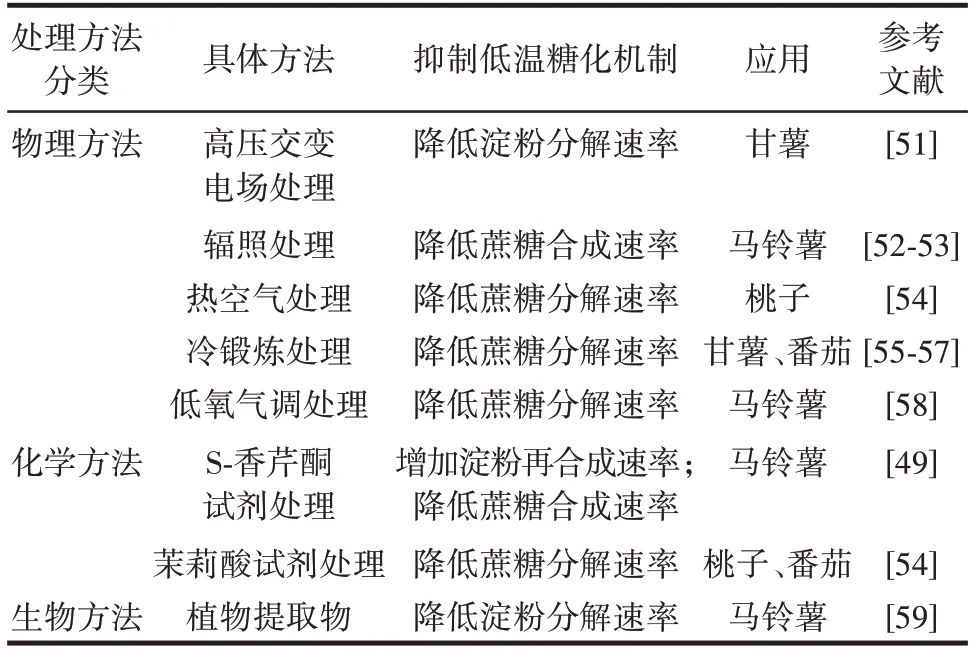
表1 不同处理方法对果蔬低温糖化的影响Table 1 Effects of different treatment methods on cold-induced sweeting of fruits and vegetables
4 展望
本文对马铃薯低温糖化的机理、代谢途径以及影响因素进行归纳总结。关于马铃薯的低温糖化过程的研究较多,但是仍存在很多科学问题需要解决。比如许多小分子信号物质在马铃薯低温糖化过程中的作用机理尚不明确;在不影响生长发育的前提下,根据遗传转化等手段可以调控低温糖化过程中的关键基因,能够改善低温贮藏的马铃薯加工特性,但对于其特性改变是否引起其它重要营养物质成分及含量变化尚未见报道;对于马铃薯低温糖化现象深究,是块茎低温胁迫的一种反应机制,推测采用适当外源处理技术提高马铃薯对低温胁迫的抗性,可改善低温糖化现象,但新型马铃薯低温糖化抑制剂的开发尚未有深入研究,高效马铃薯低温糖化控制技术尚待研发。
猜你喜欢
今日农业(2021年15期)2021-11-26
广西糖业(2020年3期)2020-09-25
中国农业科技导报(2020年1期)2020-03-15
中国真菌学杂志(2019年1期)2019-01-23
浙江工业大学学报(2017年5期)2018-01-22
现代园艺(2017年19期)2018-01-19
中国糖料(2016年1期)2016-12-01
云南中医学院学报(2015年2期)2015-07-31
中国果业信息(2015年11期)2015-01-23
农业科技与装备(2014年6期)2014-11-28
