
纳米塑料和酮康唑对霍甫水丝蚓肠道的影响
2024-03-28 08:13陆光华喻叶庭刘建超河海大学环境学院浅水湖泊综合治理与资源开发教育部重点实验室江苏南京210098
中国环境科学 2024年3期
陆光华,喻叶庭,薛 琪,刘建超(河海大学环境学院,浅水湖泊综合治理与资源开发教育部重点实验室,江苏 南京 210098)
随着个人护理品(PCPs)的生产和使用量与日俱增,其中添加的杀菌剂等活性成分和微塑料(MPs)等非活性成分引起了广泛关注.在中国,每年因沐浴露和洁面乳的使用而释放到环境中的微塑料分别为39 和307t[1-2].PCPs 中的活性成分和MPs 通过生活污水、工业废水、污水处理厂尾水等排到自然水体中,并蓄积在沉积物系统.酮康唑(KCZ)是广泛添加在PCPs 中的广谱抗真菌剂,已在全球各地的沉积物系统中广泛检出.KCZ 急性暴露降低了蛋白核小球藻的细胞活性,并诱导细胞凋亡和氧化损伤[3].
在PCPs 中添加的MPs 可能在机械加工等过程中破碎形成潜在危害更大的纳米塑料(NPs),环境MPs 也会不断降解形成NPs[4-5].NPs 的高比表面积和疏水性使其容易吸附环境中的疏水性有机污染物,被底栖动物摄取进入肠道后,在各种消化酶的促进下可能发生解吸,已解吸的污染物和未解吸的污染物则继续穿过各类生物屏障而转移到各个组织和器官,进而产生毒理学效应[6].此外,纳米粒子的表面功能化对其稳定性、在生物介质中的分散、生物相容性和生物分布至关重要[7].比如,氨基修饰聚苯乙烯纳米塑料(PSNH2)接触海胆胚胎会导致畸形并抑制胚胎的早期发育,而羧基修饰聚苯乙烯纳米塑料(PSCOOH)对胚胎发育几乎没有影响[8].此外,由于带电NPs 和质膜之间的静电相互作用,如与受体蛋白、粘蛋白和磷脂结合,NPs 很容易跨越生物屏障并引起毒性[9].NPs 的表面官能团所带的电荷是决定NPs 内化到上皮细胞和随后的细胞毒性的关键因素[10].
本文以原始聚苯乙烯NPs、PSNH2、PSCOOH和环境相关浓度的KCZ 为目标污染物,以底栖环节动物霍甫水丝蚓(Limnodrilus hoffmeistteri)为模式生物,研究NPs 对KCZ 诱导的霍甫水丝蚓肠道损伤及肠道菌群失调的影响,以期为理解PCPs 相关污染物复合污染对底栖生物的交互作用提供参考.
1 材料与方法
1.1 供试材料
参照OECD 225 方法进行人工沉积物制备[11].酮康唑(≥98%)购自南京荣华科学器材有限公司,100nm 原始聚苯乙烯(PS)、PSNH2和PSCOOH绿色荧光微球购自天津赛尔群科技有限公司.HPLC级甲醇和丙酮购自德国 Merck 公司(Darmstadt,Germany),胃蛋白酶、纤维素酶、D-乳酸盐和二胺氧化酶测定试剂盒购自滁州仕诺达生物科技公司.霍甫水丝蚓购自天津市花鸟鱼虫市场.
1.2 暴露实验
设置4 个处理组(KCZ、KCZ+PS、KCZ+PSNH2、KCZ+PSCOOH)和1 个空白对照组,包括3 组平行.根据OECD 225 方法[11]将KCZ 和NPs 添加到人工沉积物中,混合均匀.KCZ 和NPs 的暴露浓度为100ng/gdw,不会对水丝蚓产生急性毒性[12].取加标沉积物50g 置于250mL 烧杯中,加入曝气超纯水,使沉积物与上覆水的体积比约为1:4.室温下避光静置7d,使水-沉积物系统中污染物达到平衡状态.
挑选大小和成熟度一致的霍甫水丝蚓100mg(约40 条)加入到每个处理组中,避光连续暴露7d.每天用曝气超纯水补足蒸发掉的水分,使上覆水与沉积物比例稳定.
1.3 KCZ 和NPs 的定量分析
在暴露第1,3,7d,收集沉积物和水丝蚓样品,经冷冻干燥后,采用加速溶剂萃取仪(Dionex ASE 350,Sunnyvale,美国)萃取,将提取物通过旋转蒸发至干燥,用2mL 甲醇超声溶解,用0.22μm 有机滤膜(尼龙66)过滤.采用Waters AcquityTMUPLC-MS/MS(Waters,Miford,MA,USA)高效液相色谱-三重四极杆质谱系统测定KCZ 的浓度.沉积物和水丝蚓中KCZ 的检测限(LOD)分别为2.5ng/g dw 和0.045ng/g ww(湿重),定量限(LOQ)分别为 8.3ng/g dw 和0.15ng/g ww.沉积物加标回收率为90.49%~112.59%,水丝蚓加标回收率为85.95%~122.13%[12].
在暴露第7d,将每个处理组中所有水丝蚓挑出,用超纯水清洗干净,吸干表面水分,称重.然后取50mg 样品并用10mL 的KOH(10% W/V,60°C)进行消化处理24h.参考Zhou 等[13]的方法,采用日立F-7000 荧光分光度计对NPs 进行定量,激发波长为488nm,发射波长为518nm.KOH 消解提取NPs 的荧光强度回收率在97%~109%.
1.4 肠道组织病理学分析和消化酶活性测定
暴露结束后,取水丝蚓1cm 肠道在4%多聚甲醛溶液中固定24h 以上.切片脱蜡至水,用苏木精-伊红染色,脱水封片,在显微镜下进行组织病理学观察.
取水丝蚓肠道组织,按照1:9(m:V)的比例加入生理盐水,制备组织匀浆液,3000r/min 离心10min,取上清液.根据试剂盒的操作步骤进行胃蛋白酶、纤维素酶、二胺氧化酶活性和D-乳酸盐含量的测定.
1.5 肠道菌群16S rRNA 扩增子测序分析
暴露结束后,使用磁珠DNA 提取试剂盒从肠道内容物中提取微生物群的基因组DNA.通过ABI GeneAmp®9700PCR 热循环仪(ABI, CA)扩增细菌16S rRNA 基因的高可变V3-V4 区域,采用的引物为338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3').将同一样本的PCR 产物混合后用2%琼脂糖凝胶电泳检测.参照电泳初步定量结果, 将 PCR 产物用QuantiFluor™-ST 蓝色荧光定量系统(Promega 公司)进行定量.然后建立MiSeq 文库,进行Illumina 测序.
1.6 数据处理与分析
采用SPSS(ver. 26.0)对数据进行统计分析.各组之间的差异通过单因素方差分析和Tukey 检验进行评估,显著性水平设定为P<0.05.生物信息分析采用QIIME(ver. 1.6.0)完成.通过 UPARSE(7.1, http://drive5.com/uparse/)对具有97%相似性截止的操作分类单元(OTU)进行聚类、识别和删除拟合序列.通过RDP 分类器(http://rdp.cme.msu.edu/)用16S rRNA数据库(SILVA SSU 128)的置信阈值分析每个OTU代表序列的分类.
2 结果与讨论
2.1 KCZ 和NPs 在水丝蚓体内的累积
如图1 所示,在单独暴露组,蚓体内KCZ 的浓度随暴露时间的增加而增加,但是第3d 和第7d 没有显著性差异.在联合暴露组,KCZ 的浓度随时间的增加逐渐降低.与KCZ 单独暴露相比,除了共存的PSNH2在第1d 显著增加了KCZ 的生物累积(P<0.05)外,NPs 的存在都显著降低了KCZ 的生物累积水平(P<0.05).3 种NPs 对KCZ 生物累积的抑制率在第7d 达到最高(88.9%~94.8%).
塑料颗粒的尺寸是MPs 影响KCZ 生物有效性的主要因素之一,本实验室以前的研究表明,100μm的PS 共暴露7d 显著增加了KCZ 的累积[12],而本研究100nm PS 共存显著降低了KCZ 的累积.NPs 的载体效应可能会增加污染物在生物体内的积累水平和毒性,表现出明显的污染物增敏效应[14];也可能因为NPs 表面吸附了污染物后使团聚速率增加,从而降低了污染物在生物体内的生物有效性,表现出稀释和抑制作用[15].研究发现,100nm 和1μm 的PS 在第7d 时显著降低了菲在蚯蚓体内的累积,而10μm和100μm 的PS 在第7d 时显著增加了菲在蚯蚓体内的累积[16].此外,污染物在NPs 表面与生物体之间的逸度梯度是影响其生物有效性的重要因素[17].随着体内KCZ 浓度增加,使NPs 上负载的污染物浓度与生物体脂质中的污染物浓度之间存在负向的浓度逸度梯度,未吸附饱和的NPs 可以从生物体内竞争吸附污染物并排出[18],这可能是导致共暴露组生物体内KCZ 浓度逐渐降低的原因.
对比3 个共暴露组,PSNH2组在第1d 和3d 时KCZ 的累积水平显著高于原始PS 和PSCOOH 组.这可能与它们在水中不同的聚集水平有关.有研究表明,PSCOOH 在水中倾向于形成微米尺寸的聚集体,而PSNH2在水中形成纳米尺寸的聚集体[19],PSNH2聚集体具有更大的比表面积,使其吸附更多KCZ 进入生物体内,从而增加KCZ 的生物累积.
通过检测第7d 时NPs 在生物体内的浓度,发现PSNH2的累积水平最低(2.78±0.86)ng/mg, 而PSCOOH 的累积水平最高(22.85±0.97)ng/mg,PS 次之(20.03±1.38)ng/mg.这一方面说明PS 的表面官能团影响了其自身的生物累积性[20];另一方面,对KCZ生物有效性的影响也不同,PSNH2的影响更大.
2.2 NPs 和KCZ 诱导的组织损伤
PS-KCZ 和PSNH2-KCZ 共暴露7d,水丝蚓的体重比对照组显著降低,其他处理组与对照组没有明显差异.肠道组织形态的严重变化可能导致消化系统功能障碍以及能量代谢和营养摄入的改变[21],从而影响生物的生长发育.如图2 所示,KCZ 和NPs 对表皮和肠道有损伤作用,可观察到各种不连续的病理变化,包括表皮和肠细胞退化、表皮和肠细胞增生、表皮表面不规则和产氯组织变性.

图2 不同暴露组水丝蚓肠道的组织切片Fig.2 Histopathology of the worm on Day 7 in different exposure groups
细胞的退化可随着污染物浓度的增加而增加[22].KCZ 单独暴露组和PSNH2与KCZ 共暴露组中水丝蚓表现出严重的细胞退化,可能与这两组KCZ 的累积水平较高有关.此外,PSNH2与KCZ 共暴露组中水丝蚓肠道组织破环相比于其他暴露组更加严重.由于PSNH2比PSCOOH 穿过细胞的运输率更高[23],PSNH2-KCZ 更容易进入肠细胞,诱导炎症反应.另一个明显的组织损伤是产氯组织变性,产氯组织是经过修饰的腹膜细胞,通过捕获和储存毒物来发挥解毒系统的功能,从而降低污染物的毒性作用[22].带不同电荷的NPs 通过差异控制肠道稳态相关分子的表达,导致肠道屏障缺失,从而使KCZ 分子更容易直接作用于肠细胞,引发组织损伤.
2.3 NPs 和KCZ 对肠道消化酶活性的影响
暴露7d 后,如图3 所示,所有暴露组中水丝蚓的胃蛋白酶活性都显著低于对照组,KCZ 单独暴露的抑制效应最强,抑制率为44.4%.KCZ 单独暴露时显著抑制了纤维素酶的活性,而共暴露增加了纤维素酶的活性,尽管官能团修饰的NPs 与KCZ 共暴露组纤维素酶活性与对照组没有显著性差异.PSNH2与KCZ 共暴露组D 乳酸盐含量显著下降,PSCOOH 与KCZ 共暴露组二胺氧化酶活性显著降低,而其他处理组D 乳酸盐含量和二胺氧化酶活性与对照组没有显著性差异.

图3 不同暴露组水丝蚓胃蛋白酶(a)、纤维素酶(b)、二胺氧化酶(c)活性和D 乳酸盐含量(d)Fig.3 Activities of pepsin(a), cellulose(b), D-lactate(c) and diamine oxidase(d) in different exposed groups on day 7
胃蛋白酶是一种由软体动物肝胰腺分泌的蛋白水解酶,可将蛋白质分解为小片段肽和氨基酸,由细胞吸收.研究发现,一些药物进入胃肠道会与胃蛋白酶相互作用,影响其结构和活性[24];聚丙烯MPs 膳食暴露显著增加了淡水底栖软体动物(Pomacea paludosa)胃蛋白酶活性[25].本文中共存NPs 减轻了KCZ 对胃蛋白酶的抑制效应,一方面可能与塑料颗粒对胃蛋白酶活性的诱导效应有关,另一方面与共暴露组水丝蚓体内KCZ 的累积水平显著降低有关.
纤维素酶活性变化直接反映底栖生物分解有机质的能力[26].乙草胺暴露显著降低了蚯蚓体内的纤维素酶活性[27],而PS-NPs暴露显著增加了贻贝纤维素酶活性[28],底栖动物可能通过提高纤维素酶活性部分生物降解PS[29].
D 乳酸盐作为细菌发酵的代谢产物可以由肠道中的各种细菌产生.当肠黏膜通透性增加时,肠道细菌产生的大量D 乳酸盐会通过受损的黏膜进入血液,然后血清中D 乳酸盐水平增加,肠组织中相应减少[30].二胺氧化酶也是肠道屏障通透性指标,其在肠粘膜中比在其他细胞中更高度活化[31].当肠道受损时,二胺氧化酶会以更高的速度释放到血液中,从而导致组织中二胺氧化酶含量降低[32].与颗粒物的接触可引起肠道通透性的改变,塑料颗粒更倾向于与脂质膜相互作用并改变生物膜的性质[33].肠道通透性改变是消化道受损的迹象[34],而消化道损伤将阻碍营养吸收和能量平衡,最终恶化机体健康.
2.4 NPs 和KCZ 对肠道微生物多样性的影响
在水丝蚓肠道内容物中鉴定出1170 个OTUs.与对照组相比,KCZ 单独处理组增加了 323 个OTUs,PS-KCZ 组增加了637 个OTUs,PSNH2-KCZ组增加了52 个OTUs,PSCOOH-KCZ 组增加了176个OTUs(图4).与KCZ 单独处理相比,原始NPs 的共存进一步增加了OTUs,而官能团修饰的NPs 的共存降低了OTUs.
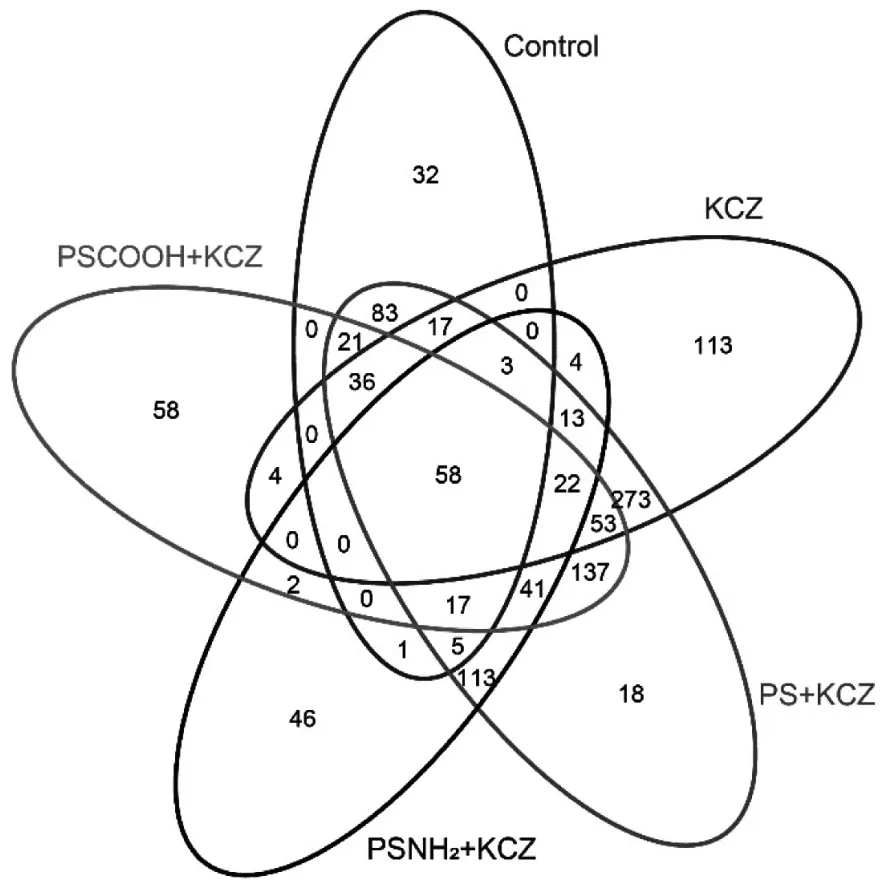
图4 不同处理组肠道微生物群的共有和特有OTUs 数量Fig.4 Shared and unique OTUs numbers in the different groups
如图5 所示,厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和拟杆菌门(Bacteroidota)为优势菌门,其相对丰度分别占总体的3.1%~86%,4.9%~64%和1.6%~20%.与对照组相比,KCZ 单独暴露显著提高了厚壁菌门相对丰度,降低了变形菌门和拟杆菌门的丰度;而共暴露降低了厚壁菌门的丰度,增加了变形菌门和拟杆菌门的丰度,尤其PSNH2与KCZ 共暴露组,变化最为显著.
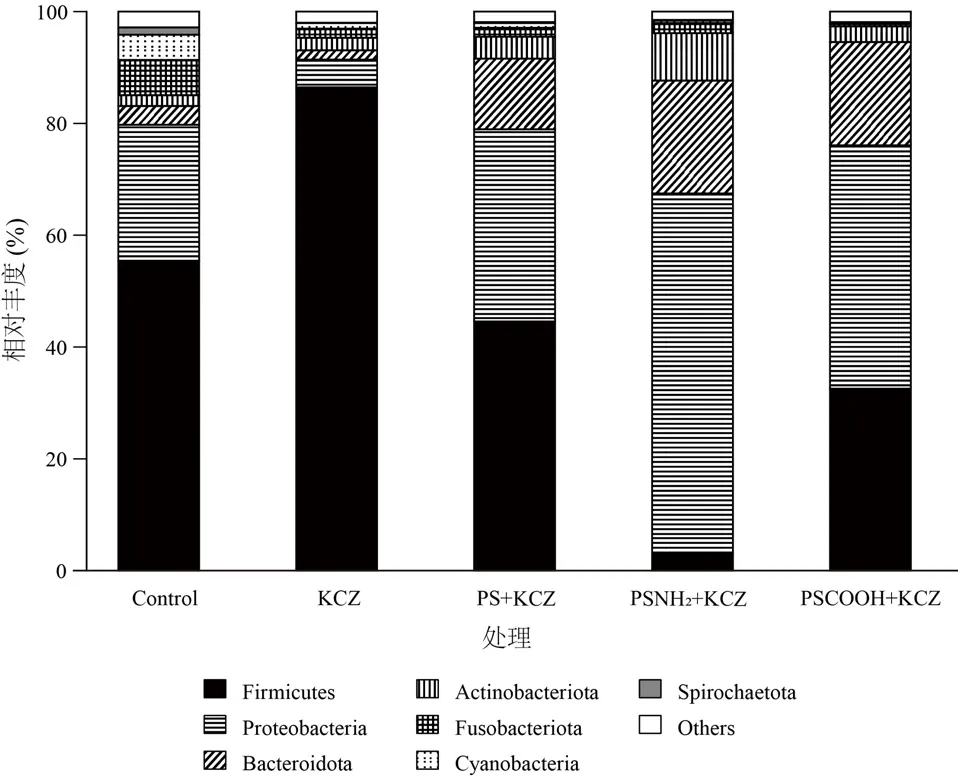
图5 各处理组肠道微生物组成在门水平的相对丰度Fig.5 Relative abundance of intestinal microbial composition at phylum level in each treatment group

图6 各处理组肠道微生物组成在属水平的相对丰度Fig.6 Relative abundance of intestinal microbial composition at genus level in each treatment group
不同处理组水丝蚓肠道前50 个优势菌属的变化见图 6.空白组优势菌属为促生乳杆菌属(Levilactobacillus)、乳杆菌属(Lactobacillus)和鲸杆菌属(Cetobacterium),KCZ 单独暴露组优势菌属为促生乳杆菌属(Levilactobacillus) 、 片球菌属(Pediococcus)和腐败乳杆菌属(Loigolactobacillus).NPs 的共存改变了优势菌属的组成,除了促生乳杆菌属仍是优势菌属外,罗河杆菌属(Rhodanobacter)和弧菌属(Vibrionimona)成为所有共暴露的优势菌属,而噬几丁质菌属(Chitinophaga)和分支杆菌属(Mycobacterium)成为KCZ-PSNH2共暴露组特有的优势菌属.
肠道微生物群在维持肠道上皮的屏障功能中起着至关重要的作用,并参与调节宿主生物的许多生理功能[35].近年来,不同的动物模型研究都报道了抗真菌剂对肠道微生物群的影响.抑霉唑和多菌灵暴露28d 可能通过改变厚壁菌、变形杆菌和放线菌丰度诱导小鼠结肠炎症和肠道微生物群失调[36-37].1000µg/L 的抑霉唑暴露21d 诱导了成年斑马鱼肠道微生物群失调,变形杆菌和拟杆菌丰度减少,而梭杆菌和厚壁菌丰度增加[38],这与本研究 KCZ 单独暴露结果一致.研究发现,150μm PS-MPs 会引起水生生物肠道菌群失调[39].化学修饰可能加剧NPs 对肠道微生物多样性的影响,Qiao等[40]发现不同官能团修饰的NPs 对小鼠肠道细菌的影响顺序为氨基修饰NPs>羧基修饰NPs>未修饰的NPs.
在门水平上,变形菌门和拟杆菌门与机体的免疫防御功能有关,其丰度下降不利于生物体防御外源污染[38].厚壁菌门/拟杆菌门的比例与营养摄入和健康状况呈正相关[41].而NPs 的共存导致厚壁菌门丰度降低,拟杆菌门丰度增加,表明水丝蚓肠道中能量摄入减少,这可能是导致共暴露组水丝蚓体重显著下降的原因之一.变形菌是一种已知的包含多种条件致病菌的门[40],NPs 共存不利于KCZ 对致病菌的消除.拟杆菌与纤维素的消化密切相关[41],前述共存的NPs 缓解了KCZ 对纤维素酶活性的抑制效应,这可能与拟杆菌丰度增加有关.
在属水平上,罗河杆菌属、弧菌属、噬几丁质菌属和分支杆菌属都属于革兰氏阴性菌,含有多种致病菌,由革兰氏阴性菌产生的脂多糖是对肠道屏障功能有害的物质[42],因此其过度生长往往伴随着肠道屏障功能的恶化[43].这表明NPs 的共存降低了 KCZ 对致病菌的抑制效应,特别是与PSNH2共存使优势菌属全部变成致病菌,增加了水丝蚓感染的风险.
3 结论
3.1 3 种NPs 在霍甫水丝蚓体内的累积水平不同,PSNH2的生物累积性最低;除了PSNH2在共暴露第1d 促进了KCZ 的生物累积,其他情况下共存NPs 均降低了KCZ 的生物累积水平.
3.2 3 种NPs 的共存显著降低了KCZ 对胃蛋白酶和纤维素酶活性的抑制;官能团修饰NPs 增加了水丝蚓肠道通透性,加重了KCZ 诱导的肠道损伤.
3.3 KCZ 单独暴露显著增加了水丝蚓肠道微生物的多样性,官能团修饰NPs 的共存使微生物多样性降低,并改变了门水平的丰度和优势菌属,尤其是PSNH2共存时优势菌属均为致病菌,增加了水丝蚓感染的风险.
猜你喜欢
化工时刊(2023年3期)2023-03-18
河南医学研究(2022年19期)2022-10-19
上海理工大学学报(2021年3期)2021-07-20
中国比较医学杂志(2020年4期)2020-05-26
水生生物学报(2019年4期)2019-07-20
生态学报(2019年11期)2019-07-08
生物安全学报(2019年3期)2019-02-15
川北医学院学报(2019年6期)2019-02-10
西北工业大学学报(2015年1期)2016-01-19
中国塑料(2014年8期)2014-10-17
