
内质网应激的免疫抑制效应
2024-04-09 08:01蒋丽娜蒋孙班赵自刚河北北方学院微循环研究所张家口075000
中国免疫学杂志 2024年3期
蒋丽娜 蒋孙班 赵自刚 (河北北方学院微循环研究所,张家口 075000)
内质网是蛋白质加工的细胞器,任何因素引起细胞生存环境改变均可引起内质网内稳态变化,导致内质网中错误折叠或未折叠的蛋白积聚,称为内质网应激(endoplasmic reticulum stress,ERS)[1]。ERS进一步激活未折叠蛋白反应(unfolded protein response,UPR),长时间UPR激活会诱导细胞凋亡、黏膜屏障损伤等一系列变化[2]。近年研究显示,过度ERS引起的免疫细胞损伤及免疫器官损伤与免疫功能障碍相关疾病联系紧密。为探究ERS引起免疫细胞、免疫器官功能障碍的作用与机制,阐明ERS免疫抑制效应在相关疾病发生发展中的作用,本文综述ERS在免疫器官、免疫细胞中的作用及其在炎症性疾病和肿瘤等发展进程中的作用,为减少ERS对免疫细胞的损伤、恢复免疫细胞功能稳态提供依据,为相关疾病预防、治疗及预后提供新思路。
1 免疫器官ERS
1.1 脾脏 作为最大的外周免疫器官,脾脏含有人体约1/4淋巴细胞,是免疫应答发生的重要场所[3-4]。研究指出单个突变的溶菌酶可通过激活肌醇需求酶1(inositol-requiring enzyme 1,IRE1)-X盒结合蛋白1(X-box binding protein 1,XBP1)途径以及促进葡萄糖调节蛋白78(glucose regulated protein 78 kD,GRP78),亦称免疫球蛋白重链结合蛋白(immunoglobulin heavy chain binding protein,BIP)表达,导致脾脏等器官中的细胞为维持内质网稳态发生凋亡,并发生家族性系统性淀粉样变性[5]。但为什么突变的溶菌酶仅诱导IRE1系统而未动员其他系统还有待研究。氟化钠可诱导以伴侣蛋白BIP、GRP94分泌增加为特点的脾细胞ERS,进而激活UPR,观察到应激相关凋亡蛋白Caspase-12、磷酸化c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)以及ERS相关蛋白环磷酸腺苷反应元件结合转录因子同源蛋白(C/EBP-homologous protein,CHOP)升高,结合已知细胞凋亡通路可说明氟化钠通过CHOP、IRE1-JNK、IRE1-肿瘤坏死因子受体相关因子2(tumor necrosis factor receptor associated factor 2,TRAF2)-Caspase-12 3条途径使脾细胞活力下降,凋亡数增加,使用ERS抑制剂4-苯基丁酸(4-phenylbutyric acid,4-PBA)后该现象被有效遏制[6]。可见ERS确实在凋亡通路激活过程中发挥作用。曲霉毒素A可通过引发ERS激活TNF-α通路同时降低IL-2分泌。IL-2在淋巴细胞增殖分化中发挥重要作用。曲霉毒素A通过抑制IL-2分泌降低猪脾嗜硌核细胞、淋巴细胞和T细胞生存能力及抗氧化能力,破坏脾脏组织结构[7]。最近研究表明,玉米赤霉烯酮(Zearalenone,ZEN)可通过激活细胞内ERS产生大量活性氧(reactive oxygen species,ROS),破坏谷胱甘肽系统稳定,降低抗氧化相关酶浓度。同时ZEN诱导凋亡蛋白Caspase-12、Caspase-3产生,破坏抗凋亡蛋白B细胞淋巴瘤/白血病-2(B cell lymphoma/lewkmia-2,Bcl-2)浓度稳定,最终诱导脾内淋巴细胞凋亡[8]。抗氧化剂,如硒可减少脾细胞损伤。研究证实硒作为维持机体免疫功能的必需微量元素,其缺乏通过改变鸡脾脏氧化应激状态、调控炎症因子基因表达和破坏脾细胞核结构导致ERS过度激活,使脾脏免疫功能受抑制[9]。总之,ERS产生后,脾细胞及脾内淋巴细胞生存活力下降,凋亡率上升,免疫系统不可避免地受到抑制。
1.2 骨髓 骨髓作为中枢免疫器官,是各类血细胞(包括免疫细胞)的发源地,也是多种细胞(如B细胞)分化、发育、成熟的场所。骨髓功能缺陷不仅严重影响机体造血功能,也导致严重的细胞免疫和体液免疫功能障碍。研究发现急性髓细胞白血病患者骨髓中,早期侵入的白血病细胞可通过分泌细胞外囊泡将ERS信号在组织间隔中传递,使基质细胞成分发生变化,可能与白血病发生发展机制有关[10]。类似研究表明获得性再生障碍性贫血、多发性骨髓瘤及周期性粒细胞减少疾病中均有ERS增强现象[11-13]。另有研究发现UPR参与脊髓物理损伤,如上调ztbt38基因表达,可使URP通路中ATF4与其启动子结合不受干扰,逆转造血细胞凋亡[14]。最新研究发现骨髓细胞中ERS激活影响鞘脂合成。作为细胞膜重要结构之一,鞘脂减少导致干细胞分化障碍、粒细胞生成减少[15]。曾有鞘脂生物合成途径几种酶及其相关代谢物与急性和慢性髓系白血病的相关报道[16]。特发性肺纤维化病变发生后,骨髓干细胞可通过分泌肝细胞生长因子(hepatocyte growth factor,HGF)发挥调节作用,如在肺泡上皮细胞表面分泌一些活性蛋白,维持内质网稳态以修复上皮细胞损伤[17]。骨髓间充质干细胞(bone mesenchymal stem cell,BMSC)是骨髓基质中造血微环境的重要细胞,有研究指出地塞米松增加BMSC PERK、ATF6和IRE1a表达,并诱导Nrf2核移位进而引发细胞凋亡,而抑制剂逆转了其对细胞的不利作用[18]。说明抑制过度ERS维持内质网稳态有助于治疗骨髓相关疾病,并有利于保护免疫系统。
2 免疫细胞的ERS
2.1 T淋巴细胞 T淋巴细胞是介导细胞免疫的核心细胞,在细胞免疫和体液免疫中发挥重要作用。急性感染能够增加T淋巴细胞凋亡导致免疫应答障碍,抑制机体免疫功能[19]。研究发现小鼠口服除草剂阿特拉津(Atrazine,ATR)可激活ERS中的PERKEIF2-ATF4-CHOP轴,引起T淋巴细胞凋亡,损害小鼠免疫功能,产生免疫毒性[20]。研究表明免疫抑制剂FK506处理后的T细胞因激活ERS进而激活细胞凋亡通路导致T细胞凋亡[21]。除成熟的T淋巴细胞直接发生免疫应答外,幼稚T细胞正常存活也对分化成不同类型T细胞以应对不同微生物入侵起重要作用[22]。初始T细胞存活和正常功能依赖于持续静止,这样才能获得对细胞凋亡的抗性和合适的生存环境[23]。研究发现小鼠体内初始T细胞内ERS会诱导胆固醇从头合成增加,使脂质和胆固醇稳态紊乱,导致其静止状态破坏,抑制小鼠免疫功能[22]。此外,T细胞ERS也影响其抗肿瘤免疫效能。研究表明小鼠中高度分化的肿瘤浸润性淋巴细胞(tumor infiltrating lymphocytes,TILs)CD8+T表面表达的程序性细胞死亡蛋白1(programmed death-1,PD-1)与肿瘤细胞表达的肿瘤细胞表达程序性死亡配体1(programmed death-ligand 1,PD-L1)结合后,启动ERS过表达,诱导UPR中PERK轴活化,激活的CHOP进一步活化其下游靶标内质网氧化还原酶1α(ER oxidoreductase 1α,ERO1α),使线粒体中ROS过量积累,线粒体衰竭而使相应TIL凋亡,肿瘤免疫抑制作用明显[24]。研究发现胰腺癌细胞通过分泌外泌体向T淋巴细胞传递信号,通过激活p38 MAPK诱导ERS,导致T淋巴细胞凋亡[25]。HIV最主要的特征是CD4+T细胞数量减少,ERS与HIV引发的CD4+T细胞凋亡有关。研究发现HIV反式激活蛋白(HIV transactivator protein,Tat)可通过激活ERS和UPR反应的3条通路激活其下游Caspase凋亡通路,导致CD4+T细胞时间依赖性凋亡,HIV患者抗病毒免疫力逐渐下降,维持或恢复内质网功能可能为治疗HIV相关疾病的假设提供了实验依据[26]。
2.2 单核巨噬细胞 单核巨噬细胞属于专职抗原提呈细胞,也是细胞内致病细菌和寄生虫的主要防卫系统,是机体防御系统的重要组成。细菌感染或暴露于某些持续性病毒感染情况下,细胞个体可能发生过早免疫衰老,抗菌免疫力下降[27]。研究表明单核细胞衰老与其ERS及UPR激活有关,UPR的IRE1-XBP1分支激活通过膜结合Toll样受体激活单核细胞/巨噬细胞促进炎症相关细胞因子产生[28]。同时IRE1可激活其下游JNK等凋亡途径,与高尔基体应激共同促进衰老相关分泌表型(senescence-associated secretory phenotype,SASP)。SASP活化导致DNA损伤积累,加速细胞衰老进程,导致单核细胞早衰[29]。关于重金属镉的研究主要停留于其对器官的损害层面,近来研究表明单核细胞中,镉通过影响钙代谢并抑制内质网膜中的钙泵扰动钙稳态,导致内质网腔中Ca2+含量降低和内质网功能受损,引起ERS。钙蛋白酶被激活以响应氧化应激介导的ERS,进而触发镉诱导的自噬和细胞凋亡,导致机体体液免疫应答或抗原呈递功能受损[30]。ERS还通过影响单核细胞分化产生免疫抑制作用。研究发现巨噬细胞极化与肿瘤细胞ERS密切相关。抑制ERS能够促进M1型巨噬细胞向M2型极化,利于肿瘤发生、发展[31]。研究表明单核细胞被乙型疱疹病毒亚型HHV-6B病毒感染后诱导UPR选择性激活,增加IRE1α和eIF2α磷酸化并上调ATF4和CHOP表达,但不影响ATF6和BIP表达。该病毒最终通过CHOP激活凋亡蛋白导致单核细胞凋亡,且单核细胞分化为树突状细胞(dendritic cells,DCs)能力明显降低,其抑制单核细胞分化的机制尚不清楚[32]。同T淋巴细胞一样,脂质异常堆积也会抑制单核细胞生存。研究发现二氧化硅纳米颗粒(silica nanoparticls,SiNPs)在巨噬细胞中与氧化极低密度脂蛋白(oxidized low-density lipoprotein,oxLDL)共同暴露,通过ERS中的PERK/eIF2α/ATF4和IRE1α/XBP1信号级联反应诱导泡沫细胞产生以及对胆固醇流入/流出平衡的干扰,加剧单核细胞凋亡[32]。
2.3 DCs DCs作为专职抗原递呈细胞,通过向T细胞呈递抗原进一步促进T细胞活化、分化和免疫记忆形成。DCs发生ERS后免疫抑制作用主要体现在热损伤和肿瘤微环境中。研究表明肿瘤微环境中常见的不利条件(如缺氧、营养剥夺和/或氧化应激)会导致蛋白质错误折叠,触发ERS,并在肿瘤相关DC(tDC)中强烈激活IRE1α-XBP1途径。tDC中高水平的ROS促进细胞内脂质过氧化作用,导致细胞内血脂异常蓄积,进而抑制抗原肽有效结合MHCⅠ分子,从而损害DC对T细胞的抗原呈递作用[33]。研究表明XBP1缺陷型tDC无法积累细胞内脂质,反而有更强的增殖能力,可在体内外支持T细胞功能,并产生更多记忆性T细胞[34]。也有研究指出肾小球炎症发病过程中,聚集的尿调节蛋白通过激活ERS诱导吞噬后的DC耐受,减少基因突变导致的组织损伤[35]。此外,热损伤后机体免疫抑制作用中也观察到DCs内发生了ERS。除促进分泌凋亡相关蛋白外,ERS还降低DCs表面IL-12、调节关键介质TNF-α等分子水平,抑制其成熟。热损伤中XBP通路缺失导致免疫抑制更严重,如CD80、CD86、MHCⅡ共刺激分子表达下降,因此XBP-1在DCs成熟和免疫调节中起关键作用[36]。提示调节ERS信号通路可能在烧伤治疗中有重要意义。
3 ERS的免疫抑制效应
免疫系统的器官、组织和细胞参与免疫应答,这些组分正常行使其功能以及相互协调,在维持机体免疫功能稳态中起重要作用。任何环节出现问题都会造成免疫功能障碍、机体抗感染能力减弱或产生自身免疫。免疫应答过强会产生自身抗体,这些抗体攻击正常组织造成组织损伤。机体处于免疫抑制状态则易受细菌、真菌及病毒感染,引起炎症性疾病,同时免疫抑制带来的免疫逃逸也成为肿瘤发生的关键因素。
3.1 炎症性疾病中ERS的免疫抑制效应 一般来说,慢性炎症状态下,ERS可激活免疫抑制以减轻过度炎症。但免疫抑制状态在持续炎症性疾病中具有诸多有害作用,可能导致免疫缺陷、细胞衰老和组织变性。研究表明慢性炎症中的ERS会增加免疫细胞抑制表型,如巨噬细胞M1和M2转换。M1巨噬细胞在急性炎症中被激活,而M2巨噬细胞的功能则是消退炎症和修复受损组织。ERS会进一步激活免疫抑制网络,参与衰老相关疾病,如阿尔茨海默病[37]。免疫抑制细胞分泌的免疫抑制细胞因子,如TGF-β、IL-10在受累组织中仅起旁观者效应,进一步表明ERS在控制炎症反应和免疫抑制平衡中起重要作用[38]。研究发现ERS条件下,肾小球肾炎时肾小球系膜细胞表现出对促炎因子的无应答性。ERS不仅诱导产生锌指蛋白A20(NF-κB主要负调节剂),抑制抑制性κB激酶(inhibitory kappa B kinase,IKK)磷酸化,抑制NF-κB激活以进一步抑制细胞对炎症细胞因子刺激的反应。同时ERS促进TRAF2蛋白降解,而TRAF2对NF-κB的激活至关重要,参与TNF-α合成。说明TNF-α激活的NF-κB比IL-1β激活的NF-κB更易受ERS影响,这些机制可能在阻止急性炎症进展和自发性免疫失能中发挥重要作用[39]。
3.2 肿瘤中ERS的免疫抑制效应 癌细胞ERS诱导曾是诱导癌细胞凋亡的有效策略。但越来越多的研究发现,肿瘤细胞通过诱导免疫细胞ERS,损伤这些细胞同时产生免疫抑制效应以逃避攻击,维持生长。肿瘤靶向的免疫细胞除前文所提,还有骨髓来源抑制细胞(myeloid-derived suppressor cell,MDSC)等。MDSC是异质细胞群,由不成熟的骨髓细胞组成,包括不成熟的巨噬细胞、粒细胞和DCs,在肿瘤细胞中可见[40]。研究表明宿主肿瘤细胞中的ERS通过增强肿瘤微环境中MDSC的免疫抑制功能诱导肿瘤生长。具体机制可能是提高肿瘤浸润MDSC中精氨酸酶1(Arginase-1,ARG1)和诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)水平,增强其免疫抑制功能。iNOS和ARG1在MDSC中的表达对以抗原依赖方式抑制T细胞功能尤其重要[41];进一步证明髓样分化因子(myeloid differentiation factor 88,Myd88)是激活ARG1的重要衔接分子,提示Toll样受体通路与MDSC的免疫抑制效应有重要关联。另一项MDSC研究表明UPR相关激酶PERK通过刺激转录因子NRF2促进肿瘤中MDSC功能,限制细胞质线粒体DNA-STING-IFN-Ⅰ免疫刺激轴,引发抗肿瘤免疫抑制。而MDSC中的PERK缺失减少了NRF2信号传导,通过扩增胞质DNA促进STING驱动的IFN-Ⅰ产生,抗肿瘤免疫增强[42]。类似研究表明PERK或CHOP激活会促进肿瘤内T细胞功能障碍[43]。此外,PERK缺失会减弱肿瘤细胞对缺氧、DNA损伤、营养缺乏和高ROS的适应性,导致肿瘤生长变缓[44-45]。除PERK通路外,IRE1α- XBP1信号在肿瘤微环境的DCs中也有免疫抑制作用,主要机制为细胞内脂质异常蓄积。血脂异常抑制抗原肽与MHCⅠ分子结合,影响DCs将抗原交叉呈递给T细胞的过程[33]。相似地,无法累积脂滴的XBP1缺陷型DC在体内外均显示出增强的支持T细胞功能的能力,且缺乏XBP1的卵巢癌小鼠产生的记忆性(肿瘤反应性)T细胞转移到野生型卵巢后,显示出增强的抗肿瘤能力[46]。类似地,肿瘤微环境下IL-6和IL-4刺激的骨髓来源巨噬细胞形成后,IRE1的免疫抑制作用下调其趋化作用,影响正常免疫效应发挥[47]。此外,慢性淋巴细胞白血病中,中性粒细胞ERS抑制其识别抗原并产生细胞毒作用,从而下调参与肿瘤免疫的能力[48]。表明UPR的IRE1通路可调节肿瘤相关细胞产生免疫抑制。
4 小结
综上,各种免疫器官和免疫细胞均可通过激活ERS相关信号通路发挥相应生物学效应。细胞生存环境变化可导致ER过度活化,引发过度ERS和UPR,并通过内质网ATF6、IRE1α、PERK及其下游信号分子作用于细胞核内免疫应答相关基因,诱导细胞毒性,进而对免疫系统产生损伤作用,最终导致机体抗原递呈细胞抗原递呈能力下降,效应细胞功能障碍、细胞因子分泌减少、趋化作用减弱等免疫抑制效应,引发多种疾病(图1)。目前ERS在某些疾病中的分子机制和信号通路尚不清楚。今后应靶向ERS深入揭示其在免疫抑制相关疾病发生发展中的作用与机制,以期为相关疾病防治提供新的措施,为相关疾病疗效与预后判断提供新指标。
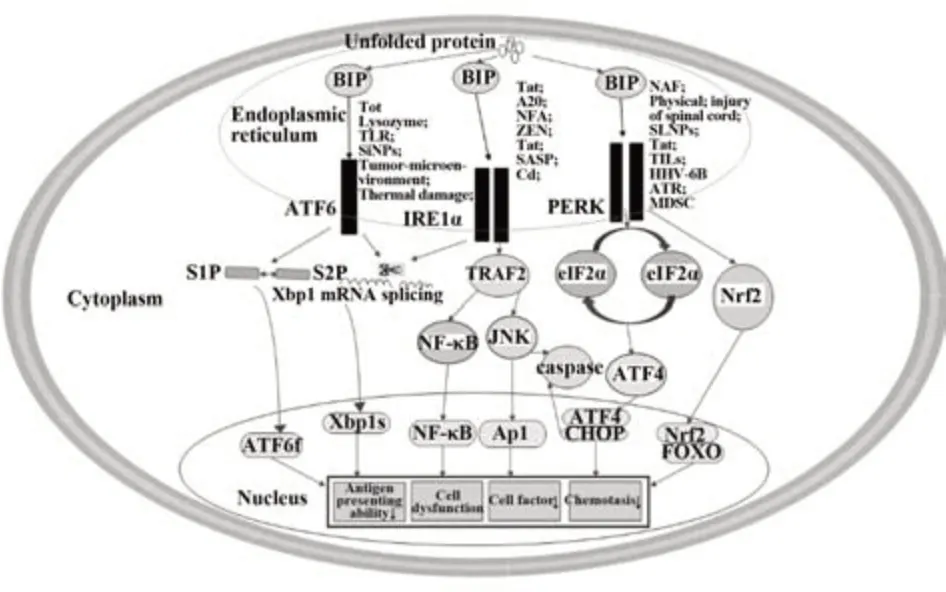
图1 参与ERS的信号通路以及ERS引起的免疫抑制效应Fig.1 Signaling pathways involved in ERS and immunosuppressive effects induced by ERS
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
解放军医学杂志(2021年12期)2022-01-18
现代畜牧科技(2021年7期)2021-07-28
现代临床医学(2021年1期)2021-01-26
兽医导刊(2019年1期)2019-02-21
安徽医科大学学报(2016年12期)2017-01-15
中国药理学与毒理学杂志(2015年3期)2015-12-16
中国当代医药(2015年33期)2015-03-01
现代检验医学杂志(2015年6期)2015-02-06
现代检验医学杂志(2014年6期)2014-02-02
