
Na2CO3胁迫对赖草叶片光合、叶绿素荧光、超微结构的影响
2024-04-23 05:45王凯鑫杜利霞
山西农业科学 2024年1期
王凯鑫 杜利霞

摘要:赖草作为黄土高原乡土草,具有耐盐碱的特点。为揭示黄土高原乡土草赖草(Leymus secalinus)的抗碱能力,采用盆栽控制试验,设置8 个Na2CO3 浓度梯度(0、50、75、100、150、200、300、400 mmol/L)对赖草幼苗进行胁迫,研究Na2CO3 胁迫对赖草幼苗的光合作用和叶绿素荧光的影响,并采用透射电镜观察叶肉细胞超微结构的变化。结果表明,随着Na2CO3 胁迫浓度的升高,赖草幼苗的净光合速率、叶绿素含量、初始荧光(Fo)和最大荧光(Fm)呈现先升高后降低的趋势,PSⅡ 最大光化学量子产量(Fv/Fm)、PSII 的潜在活性(Fv/Fo)逐渐降低。150 mmol/L Na2CO3胁迫时赖草叶片净光合速率、Fo 和Fm 达到最大,叶绿素含量较高;当Na2CO3浓度大于200 mmol/L 时,赖草幼苗叶片叶绿素含量、净光合速率和Fo 下降显著。透射电镜观察结果显示,50 mmol/LNa2CO3 胁迫下,叶绿体外膜清晰,基粒片层扭曲;150 mmol/L Na2CO3 胁迫下,叶绿体的内外膜有解体迹象,线粒体膜有破损趋势,但结构完整;400 mmol/L 胁迫下,叶绿体、线粒体内外膜解体,细胞结构严重受损。赖草对150 mmol/L 以下的Na2CO3胁迫表现出一定的适应性,叶片结构未受损伤。
关键词:赖草;Na2CO3胁迫;光合特性;叶绿素荧光;叶肉细胞超微结构
中图分类号:S543.9 文献标识码:A 文章编号:1002?2481(2024)01?0086?08
随着全球环境的加剧恶化,气候变暖导致土壤水分加剧蒸发,土壤盐碱化加重,黄土高原存在降雨量低(150~800 mm)、蒸发量高(900~1 300 mm)、碱性黄土(pH 均值为8.5)等特征[1-2],盐土中含有碳酸根,是以碳酸钠为主的苏打盐碱土壤[3],选育适应盐碱地的耐盐碱植物[4],可以将盐碱地开发利用起来。土壤盐碱化不仅会使植物幼苗的株高、生物量等表型性状受到显著抑制,还会导致叶绿体结构变形、膜系统解体[5],使叶绿素含量下降,植物光系统Ⅰ 和光系统Ⅱ 被影响[6]。因此,光合作用、叶绿素荧光和细胞结构变化可以系统地衡量逆境胁迫对植物的伤害机理[7-8]。郜新强等[9]研究表明,在低Na2CO3 浓度时,金丝楸的叶绿素总量和光合速率高于对照,随着胁迫浓度的增加,金丝楸的叶绿素总量显著下降并且光合速率受到明显抑制。刘建等[10]对柳枝稷的研究发现,随着盐碱浓度的增加,Fv/Fm、Fv/Fo 呈逐渐降低的趋势。张慧芳等[11]研究表明,随着盐碱浓度的增加,唐古特白刺叶肉细胞的叶绿体发生变形,基粒片层破坏,嗜锇颗粒及淀粉粒出现。这些研究结果为探究植物的抗盐碱能力提供了重要依据,对盐碱地改良中的抗鹽碱植物的选育具有重要作用。
赖草(Leymus secalinus)作为多年生禾草,具有根茎发达、无性繁殖能力强、耐盐碱[12]、营养价值高等特点,能在pH 值为9~10 的盐碱地上正常生长,是可合理利用的优质牧草[13]。目前,对赖草的研究主要集中在植物群落特征及生态恢复等方面,对赖草抗性方面的研究报道较少。培养和选育黄土高原区域耐盐碱植物,对实践应用和理论研究都具有很重要的意义[14]。
本试验以Na2CO3 溶液模拟碱胁迫,通过研究胁迫对赖草叶片光合作用、叶绿素荧光及细胞超微结构影响,以探究赖草的抗盐碱能力,为盐碱环境下耐碱赖草的选育提供一定的理论依据。
1材料和方法
1.1 试验材料
试验赖草种子于山西省右玉县(112°19 ′E、39°59 ′N)采集,海拔1 329 m,属温带大陆性季风气候。年平均气温4.7 ℃,生长季平均气温16.7 ℃,全年月平均最高气温22.4 ℃ ,全年月平均最低气温-17 ℃;年降水量为435 mm。土壤pH 值为9.2,属于中度盐渍化草地。
1.2 试验设计
试验在晋中市太谷区山西农业大学草业学院日光温室采取盆栽方式进行,昼夜温度25 ℃/17 ℃ ,相对湿度65%~80%。以基质蛭石和珍珠岩按2∶1 的比例均匀混合并装盆(直径25 cm、高20 cm),选取饱满一致的赖草种子温水浸泡24 h 后播种。在出苗后用Hoagland 营养液浇灌,每周一次,用称重法每天确定失水量,并在17:00—20:00 用蒸馏水对盆栽进行补充。出苗2~3 周后定苗,并于2 个月后(苗高15 cm 左右)设置7 个浓度(50、75、100、150、200、300、400 mmol/L)Na2CO3 进行胁迫,每处理重复3 次,为避免盐激伤害,第1 次浇Na2CO3浓度为50 mmol/L,此后每天按50 mmol/L 浓度梯度进行递增,直到达到处理浓度;对照(CK)只浇灌Hoagland 营养液。每个处理胁迫10 d。
1.3 测定指标及方法
1.3.1 叶绿素含量的测定
胁迫10 d 后,每个处理选取3 个长势一致且良好的赖草植株,用丙酮浸提法测定叶绿素含量[15]。
1.3.2 光合指标的测定
胁迫10 d 后,选取长势良好且一致的植株3 株,选好叶片后标记固定。在天气晴朗的9:00—11:00,用Li-6400 便携式光合测定系统测定赖草的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2 浓度(Ci)[16]。每株重复记录3 个观测值。
1.3.3 荧光参数的测定
胁迫10 d 后,用便携式叶绿素荧光仪(PAM-2500)的叶夹夹住待测叶片进行暗处理25~30 min 后,测定初始荧光(Fo)、最大荧光(Fm),计算可变荧光(Fv)、PSⅡ 最大光化学量子产量(Fv/Fm)及PSII 的潜在活性(Fv/Fo)。每个处理重复5 次。
光响应曲线分别用0、1、5、63、140、270、473、784、1 159、1 662、2 439 μmol/(m2·s)光化光照射叶片20 s,测定各光强下的实际光合量子产量(Y(Ⅱ))、光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)、表观电子传递速率(ETR)[17-18],取5 次测定的平均值。
1.3.4 叶片超微结构的观察
胁迫结束后剪下自上至下第2 片叶片,去除叶脉后切成小段(0.5 mm×1.0 mm),用3% 戊二醛溶液在0~4 ℃前固定4 d 后用0.1 mol/L PBS(pH 值7.2)漂洗3 次后,样品制备参照罗孟容等[19]的方法,Epon-812 环氧树脂渗透包埋,超薄切片机(Leica EMUC6)切片,醋酸双氧铀和柠檬酸铅双染色,透射电镜(日本JEM-1400)观察、拍照。
1.4 数据处理
利用SPSS 26.0 和Microsoft Excel 2016 进行数据整理与分析;单因素方差分析后用LSD 进行多重比较。
2结果与分析
2.1 Na2CO3胁迫对赖草幼苗叶片色素含量的影响
由表1 可知,Na2CO3胁迫对叶绿素含量的影响较明显,总叶绿素含量最大为11.93 mg/L,最小为6.78 mg/L;在胁迫浓度为300 mmol/L 时,叶绿素a、叶绿素b 和总叶绿素含量与对照间差异显著(P<0.05)。叶绿素a/b 的值在2.7~3.3,变化范围更大。当Na2CO3在75 mmol/L 浓度以下时,类胡萝卜素含量相较于CK 无显著差异,在大于100 mmol/L 浓度时含量开始显著下降(P<0.05)。
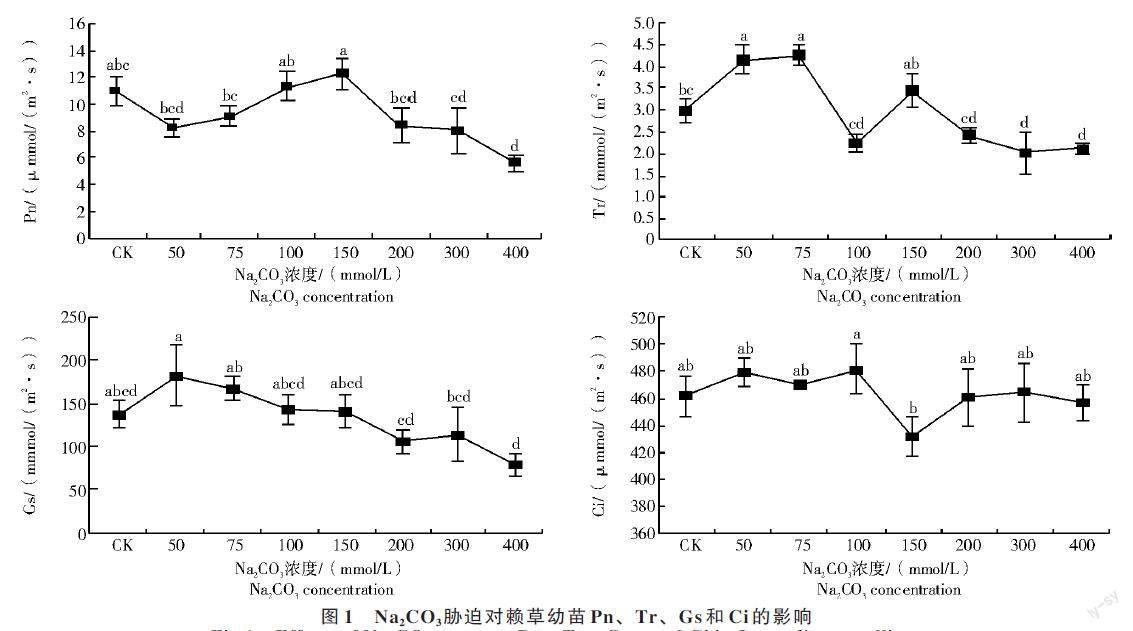
2.2 Na2CO3胁迫对赖草幼苗光合特性的影响
从图1 可以看出,当胁迫浓度低于150 mmol/L时,Pn 随胁迫浓度的升高逐渐升高,并在浓度达到150 mmol/L 时最大,浓度高于150 mmol/L 后随胁迫浓度的增加Pn 开始下降,在400 mmol/L 时降到最低,较对照组显著降低了49%(P<0.05)。Tr 在低的胁迫浓度时,比CK 增加了42%;当胁迫浓度达到100 mmol/L 时开始下降,胁迫浓度为400 mmol/L时较对照组显著降低32%(P<0.05)。Gs 值在50 mmol/L 胁迫浓度下达到最大,之后逐渐下降,当胁迫浓度升到400 mmol/L 时比对照组降低了43%。Ci 的变化与Tr 的变化一致,胁迫对Ci 的影响与对照组相比差异不显著。
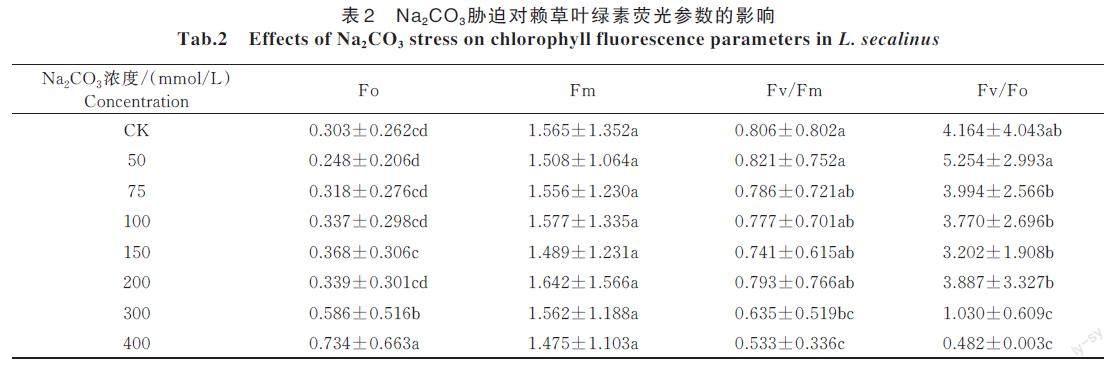
2.3 Na2CO3 胁迫对赖草幼苗叶绿素荧光参数的影响
从表2 可以看出,Fo 随Na2CO3 浓度的升高而增加,在300 mmol/L 的胁迫下Fo 与对照差异显著(P<0.05),在400 mmol/L 胁迫时差异最大;Fm 在不同的胁迫浓度下均无显著差异;在胁迫浓度小于50 mmol/L 时,Fv/Fm 和Fv/Fo 的变化趋势一致均有增加趋势,之后随Na2CO3 浓度的增加逐渐降低,Fv/Fo 的下降幅度比较大,比对照组显著降低了88.4%(P<0.05)。
2.4 Na2CO3 胁迫对赖草幼苗光响应曲线荧光参数的影响
所有Na2CO3 胁迫浓度下的Y(II)均随光合有效辐射(PAR)的增强而下降(图2);在0~140 μmol/(m2·s)范围内下降迅速,之后变得平缓。Na2CO3 浓度为50~75 mmol/L 的曲线与CK 一致,均高于高浓度下Y(II)曲线。
ETR-PAR 为不同光强下通过PSII 的表观电子传递速率。PAR 小于270 μmol/(m2·s),随着光强的增加,ETR 先迅速增加后缓慢增加;ETRPAR曲线在PAR 升高到1 159 μmol/(m2·s)时出现分歧:浓度为50、75 mmol/L 的曲线随光强增加而升高,CK 及100 mmol/L 的曲线趋于稳定,浓度高于100 mmol/L 的曲线随光强的增加开始下降(图2)。
由图2 可知,在试验光强范围内,NPQ 随光强的增加而增大,且增速较快,NPQ 未达到稳定状态,此时赖草幼苗把多余的光量子通过快速的热耗散消耗掉。浓度为300~400 mmol/L Na2CO3 胁迫的NPQ 曲线在最下面,NPQ 增加缓慢,热耗散慢,受到了光抑制,使ETR 传递的速率下降。qP 随着PAR 的增加而降低,当PAR 小于475 μmol/(m2·s)时,所有浓度下的qP 均先迅速下降,后趋于平缓,并在2 000 μmol/(m2·s)左右达到稳定。对照和浓度小于100 mmol/L 的qP 曲线下降速度较慢,qP 越大,PSII 的电子传递活性越强(图2)。
2.5 Na2CO3 胁迫对赖草幼苗光响应曲线拟合参数的影响
初始斜率在Na2CO3 浓度低于100 mmol/L 时增加(表3),随后随浓度增加而降低;最大电子传递速率ETRmax 不同浓度下均显著小于CK,在200 mmol/L 开始迅速下降。对强光的耐受能力Ik隨浓度增加而降低,所有胁迫浓度下Ik 与CK 均差异显著(P<0.05)。
2.6 Na2CO3胁迫对赖草叶肉细胞超微结构的影响
由图3-A 可知,在50 mmol/L 胁迫下,赖草叶肉细胞的液泡较大,叶绿体在细胞壁周围。叶绿体外膜清晰,基粒片层扭曲(图3-C);有淀粉粒,嗜锇体小且数量较少(图3-B)。
由图3-D 可知,在150 mmol/L 胁迫下,叶肉细胞形状不规则,叶绿体与细胞壁存在一定程度的分离现象,液泡缩小。叶绿体的内外膜有解体迹象,类囊体片层排列不整齐,有增宽现象,基粒片层排列松散且扭曲(图3-E)。嗜锇体小,但数量增多,无淀粉粒。线粒体膜有破损趋势,但结构完整,能观察到嵴结构(图3-F)。
在400 mmol/L 胁迫下,细胞呈不规则形状,细胞壁折叠扭曲严重,液泡所占比例很小(图3-G)。多个叶绿体融合粘连,难以区分,片层间隙增大、基粒排列混乱,嗜锇体显著增多,增大;叶绿体内外膜解体,有大的嗜锇体游离在外,有少量淀粉粒。解体的叶绿体内出现空洞(图3-H)。线粒体膜解体,结构不完整,嵴消失(图3-I)。
3结论与讨论
在逆境胁迫下,叶绿素的合成效率降低而降解速率增加,最终导致叶片中的叶绿素含量降低[19]。有研究发现,低浓度的盐胁迫对西瓜叶片中叶绿素含量的提高有促进作用,而高浓度则存在显著抑制作用[20]。本试验结果表明,低浓度Na2CO3 胁迫后,赖草幼苗的叶绿素a、b 高于对照组但显著不差异,但当Na2CO3的浓度逐渐升高后,导致叶绿素a、b 的含量均显著降低,叶绿素a/b 显著变化(P<0.05)。而叶绿素含量能够反映光合作用强弱,光合作用是植物生长发育的重要过程[21]。李焕勇等[22]研究表明,随着Na2CO3 胁迫浓度的升高,甜樱桃叶片的净光合速率、气孔导度显著下降。本试验结果表明,在一定浓度Na2CO3 胁迫下赖草能够维持正常的光合作用。对赖草光合作用指标进一步测定发现,Pn和Gs 在低浓度Na2CO3 脅迫下与对照组均无显著差异,但Pn 在高浓度400 mmol/L 时显著下降。但郑国琦等[23]研究表明,Pn 降低,Ci 下降,是气孔限制;Pn 降低,Ci 升高,是非气孔限制。赖草的Ci 浓度在整个胁迫浓度下较高,与对照组相比无显著变化,说明气孔不是赖草Pn 降低的主要限制因子,这可能是因为胁迫造成赖草叶片的光合结构受损导致。低浓度胁迫处理促进叶绿素合成和光合速率,而在高浓度胁迫下叶绿素合成和光合速率受到抑制,与SANTOS[24]不一致,但与郜新强等[9]的研究结果一致,这可能与物种的差异有关,也可能与试验设计的胁迫过程有关,有待进一步研究。
初始荧光(Fo)是暗适应的PSII 中心全部开放时的荧光强度,Fo 增加量越多,表明植物受损伤程度越严重[25],且光能转化效率(Fv/Fm)是早期检测逆境胁迫的最佳指标[26]。本试验结果显示,低盐碱浓度(50~75mmol/L Na2CO3)下Fo 降低,之后逐渐上升;同时Fv/Fm、Fv/Fo 也随Na2CO3浓度增加而降低,在300 mmol/L 胁迫下与CK 出现显著差异。正常叶片的Fv/Fm 在0.8 左右,300 mmol/LNa2CO3 胁迫,使Fv/Fm 的值下降,进而导致PSII的潜在光合活性降低。本试验表明,低浓度盐碱处理对赖草叶片的PSII 反应中心影响不大,但高浓度盐碱处理使植物细胞结构受损,细胞的光合结构破坏,降低了植株对光能的利用率,这与汉麻[27]的相关研究结果一致。在低光强范围内,ETR 会随光强的增加而增大,光强达到一定强度后就会稳定,稳定后多余的光能就会以热耗散的方式消耗掉(NPQ),若热耗散达到饱和,则会损伤植物(Y(NO))。在低Na2CO3浓度胁迫下,Y(II)和qP 的曲线比高浓度下的曲线高,说明低浓度(50~75 mmol/L)胁迫下幼苗能够正常生长,叶绿素吸收的光量子充分进行光合作用,随着胁迫浓度逐渐增加,叶片结构被破坏,光合能力则逐渐下降。NPQ 作为光保护的重要指标,在试验光强范围内随PAR 的增加而增大,并在所有胁迫浓度下均未达到稳定状态,表明赖草幼苗能忍受一定程度的高光强照射;Fv/Fm 随胁迫浓度升高而下降的主要原因是NPQ 增加及非光化学能量耗散的加强。
叶绿体是植物光合作用的场所,其结构完整性能够直接影响光合作用的强弱,植物受到逆境时,叶绿体结构变形,可作为植物受损的标志[28]。赖草幼苗在400 mmol/L Na2CO3 胁迫下,虽然叶绿体的内外膜消失,但基粒片层存在,叶肉细胞中存在叶绿体,这与韦存虚等[29]高浓度胁迫下叶绿体片层结构消失的研究结果不一致,从这点来说,赖草的耐碱能力更强。嗜锇体是由叶绿体解体、脂类物质聚集而成的,可用来指示叶绿体膜的完整性。本试验中,Na2CO3 浓度在150 mmol/L 时嗜锇体增加不明显,Na2CO3 浓度在400 mmol/L 时体积增大,数量增加。这与郑文菊等[30]对碱茅的研究一致,增多的嗜锇体能提高细胞质浓度、减少水分的流失,提高抗盐能力,赖草叶肉细胞中嗜锇体的增多受碱胁迫或是抵抗碱胁迫的结果有待进一步研究。作为光合作用的产物,影响淀粉粒增加的主要原因有2 个,一是由于夜间低温使得光合产物运输受阻,导致其增加;二是由于增多增大的淀粉粒,不仅为细胞的活动提供能量,而且能提高渗透压,保证了水分的吸收和利用[31],所以植物抵抗逆境的表现会使淀粉粒增多。该研究在低浓度的Na2CO3 胁迫下,能观察到较大的淀粉粒,而当浓度高于150 mmol/L 几乎没有淀粉粒,可能由于高浓度导致叶绿体结构破坏使得光合受阻,光合产物很少或没有,也可能是叶绿体结构破坏,光合产物合成速率小于运输速率,有待进一步研究。
低浓度至中浓度(50~150 mmol/L Na2CO3)的碱胁迫下,赖草对碱害表现出一定适应性。高浓度Na2CO3 胁迫导致赖草叶片叶绿体片层结构损伤,光合速率下降,幼苗生长受阻。嗜锇体增多和淀粉粒的增加,体现赖草对盐碱胁迫的适应力较强。
参考文献:
[1] LIU Q,WANG Y Q,ZHANG J,et al. Filling gullies to createfarmland on the loess plateau[J]. Environmental Science &Technology,2013,47(14):7589-7590.
[2] PENG S Z,DING Y X,WEN Z M,et al. Spatiotemporalchange and trend analysis of potential evapotranspiration overthe Loess Plateau of China during 2011-2100[J]. Agriculturaland Forest Meteorology,2017,233:183-194.
[3] 冯辽. 山西省盐碱地改良研究[D]. 太谷:山西农业大学,2017.
FENG L. The control measures of alkali-saline land in ShanxiProvince[D]. Taigu:Shanxi Agricultural University,2017.
[4] 杨鹤. 习近平主持召开深入推动黄河流域生态保护和高质量发展座谈会并发表重要讲话[EB/OL].(2021-10-22).https://www.gov.cn/xinwen/2021-10/22/content_5644385.htm#1.
YANG H. Xi jinping presided over a symposium on promotingecological protection and high-quality development of the Yel?low River Basin and delivered an important speech[EB/OL].(2021-10-22). https://www. gov. cn/xinwen/2021-10/22/con?tent_5644385.htm#1.
[5] LI N,ZHANG Z H,GAO S,et al. Different responses of twoChinese cabbage(Brassica rapa L. ssp. pekinensis) cultivars inphotosynthetic characteristics and chloroplast ultrastructure tosalt and alkali stress[J]. Planta,2021,254(5):102.
[6] OI T,ENOMOTO S,NAKAO T,et al. Three-dimensional ul?trastructural change of chloroplasts in rice mesophyll cells re?sponding to salt stress[J]. Annals of Botany,2020,125(5):833-840.
[7] BAKER N R. Chlorophyll fluorescence:a probe of photosynthe?sis in vivo[J]. Annual Review of Plant Biology,2008,59:89-113.
[8] 汪月霞,孙国荣,王建波,等. NaCl 胁迫下星星草幼苗MDA 含量与膜透性及叶绿素荧光参数之间的关系[J]. 生态学报,2006,26(1):122-129.
WANG Y X,SUN G R,WANG J B,et al. Relationshipsamong MDA content,plasma membrane permeability and thechlorophyll fluorescence parameters of Puccinellia tenuifloraseedlings under NaCl stress[J]. Acta Ecologica Sinica,2006,26(1):122-129.
[9] 郜新强,王小艳,焦伟,等. 金丝楸幼苗响应盐碱胁迫的生理和转录组分析[J]. 林业科学研究,2023,36(1):166-178.
GAO X Q,WANG X Y,JIAO W,et al. Physiological and tran?scriptomic analysis of Catalpa bungei seedlings in response tosaline-alkali stresses[J]. Forest Research,2023,36(1):166-178.
[10] 刘建,齐宝林,于洪柱,等. 盐碱胁迫对3 种柳枝稷生长性状、叶片性状及叶绿素荧光特性的影响[J]. 黑龙江畜牧兽医,2023(1):85-91.
LIU J,QI B L,YU H Z,et al. Effects of saline alkali stress ongrowth traits,leaf traits and chlorophyll fluorescence of threeswitchgrass species[J]. Heilongjiang Animal Science and Vet?erinary Medicine,2023(1):85-91.
[11] 张慧芳,闫海冰,冯帆,等. 碱胁迫对唐古特白刺生长及叶片超微结构的影响[J]. 中国水土保持科学(中英文),2021,19(1):35-42.
ZHANG H F,YAN H B,FENG F,et al. Effects of alkalistress on the growth and ultrastructure of Nitraria tangutorum[J]. Science of Soil and Water Conservation,2021,19(1):35-42.
[12] CUN W. Epicuticular wax of leaf epidermis:a functional struc?ture for salt excretion in a halophyte Puccinellia tenuiflora[J].Acta Ecologica Sinica,2004,24(11):2451-2456.
[13] 郭瑞. 松嫩平原四种禾本科植物耐盐碱生理生态机制研究[D]. 长春:东北师范大学,2010.
GUO R. The study of saline and alkaline tolerant ecophysiologicalmetabolism in four Gramineae in the Songnengrasslands[D]. Changchun:Northeast Normal University,2010.
[14] 杜利霞,董寬虎,乔志宏,等. NaCl 胁迫对赖草幼苗生理特性的影响[J]. 草原与草坪,2013,33(1):1-4,10.
DU L X,DONG K H,QIAO Z H,et al. Effect of NaCl stresson physiological characteristics of Leymus secalinus seedling[J].Grassland and Turf,2013,33(1):1-4,10.
[15] 高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006:15-17,74-77.
GAO J F. Experimental guidance for plant physiology[M]. Bei?jing:Higher Education Press,2006:15-17,74-77.
[16] 王喆,李萌,郝瑞丽,等. 糜子穗型突变体光合特性比较[J]. 山西农业科学,2020,48(6):879-883.
WANG Z,LI M,HAO R L,et al. Comparison of photosyn?thetic characteristics of broomcorn millet spike mutants[J].Journal of Shanxi Agricultural Sciences,2020,48(6):879-883.
[17] 杜利霞. 赖草对盐碱胁迫的生理响应及LsCDPK 基因的克隆与功能分析[D]. 太谷:山西农业大学,2015.
DU L X. Physiological responses of Leymus secalinus seedlingunder saline-alkali stress and the clone and function analysis ofLsCDPK gene[D]. Taigu:Shanxi Agricultural University,2015.
[18] 杨娟,王有科,李捷,等. 不同枸杞品种叶绿素荧光参数差异分析[J]. 干旱区研究,2014,31(3):550-555.
YANG J,WANG Y K,LI J,et al. Parameter difference ofchlorophyll fluorescence in leaves of different cultivars of Ly?cium barbarum[J]. Arid Zone Research,2014,31(3):550-555.
[19] 罗孟容,梁文斌,杨艳,等. 干旱胁迫对栀子光合作用及叶绿体超微结构的影响[J]. 经济林研究,2021,39(3):165-174.
LUO M R,LIANG W B,YANG Y,et al. Effect of droughtstress on photosynthesis and chloroplast ultrastructure of Gar?denia jasminoides[J]. Non-Wood Forest Research,2021,39(3):165-174.
[20] 孙凤岭,陈昆,姜涛,等. 盐胁迫对西瓜幼苗光合色素、光合特性及抗氧化保护酶系统的影响[J]. 园艺与种苗,2022,42(12):4-6.
SUN F L,CHEN K,JIANG T,et al. Effects of salt stress onphotosynthetic pigments,photosynthetic characteristics and an?tioxidant protective enzyme system of watermelon seedlings[J].Horticulture & Seed,2022,42(12):4-6.
[21] DEMMIG-ADAMS B,ADAMS W W III. Photoprotectionand other responses of plants to high light stress[J]. Annual Re?view of Plant Physiology and Plant Molecular Biology,1992,43:599-626.
[22] 李焕勇,廖方舟,刘景超,等. 盐胁迫对甜樱桃砧木生理特性及光合荧光参数的影响[J]. 西北植物学报,2023,43(1):127-135.
LI H Y,LIAO F Z,LIU J C,et al. Effect of salt stress onphysiological characteristics and photosynthetic fluorescence pa?rameters of sweet cherry rootstock[J]. Acta Botanica Boreali-Occidentalia Sinica,2023,43(1):127-135.
[23] 郑国琦,许兴,徐兆桢,等. 盐胁迫对枸杞光合作用的气孔与非气孔限制[J]. 西北植物学报,2002,22(6):75-79.
ZHENG G Q,XU X,XU Z Z,et al. The effect of salt stress onthe stomatal and non-stomatal limitation of photosynthesis ofLycium barbarum[J]. Acta Botanica Boreali-Occidentalia Si?nica,2002,22(6):75-79.
[24] SANTOS C V. Regulation of chlorophyll biosynthesis anddegradation by salt stress in sunflower leaves[J]. Scientia Horti?culturae,2004,103(1):93-99.
[25] 李旭新,刘炳响,郭智涛,等. NaCl 胁迫下黄连木叶片光合特性及快速叶绿素荧光诱导动力学曲线的变化[J]. 应用生态学报,2013,24(9):2479-2484.
LI X X,LIU B X,GUO Z T,et al. Effects of NaCl stress onphotosynthesis characteristics and fast chlorophyll fluorescenceinduction dynamics of Pistacia chinensis leaves[J]. Chinese Jour?nal of Applied Ecology,2013,24(9):2479-2484.
[26] WANG L Z,WANG L M,XIANG H T,et al. Relationship ofphotosynthetic efficiency and seed-setting rate in two contrast?ing rice cultivars under chilling stress[J]. Photosynthetica,2016,54(4):581-588.
[27] 张晓艳,孙宇峰,曹焜,等. 苏打盐碱胁迫对汉麻光合特性的影响[J]. 江西农业学报,2022,34(8):53-59,64.
ZHANG X Y,SUN Y F,CAO K,et al. Effects of soda salinealkalistress on photosynthetic characteristics of hemp[J]. ActaAgriculturae Jiangxi,2022,34(8):53-59,64.
[28] 周蕴薇,刘艳萍,戴思兰. 用叶绿素荧光分析技术鉴定植物抗寒性的剖析[J]. 植物生理学通讯,2006,42(5):945-950.
ZHOU Y W,LIU Y P,DAI S L. Identification of cold resis?tant plants by chlorophyll fluorescence analysis technique[J].Plant Physiology Communications,2006,42(5):945-950.
[29] 韦存虚,张军,王建军,等. 星星草營养器官适应盐胁迫的结构特征[J]. 植物资源与环境学报,2006(1):51-56.
WEI C X,ZHANG J,WANG J J,et al. Observation on struc?tural characters of vegetative organs of Puccinellia tenuifloraunder salt stress[J]. Journal of Plant Resources and Environ?ment,2006(1):51-56.
[30] 郑文菊,王勋陵,沈禹颖. 几种盐地生植物同化器官的超微结构研究[J]. 电子显微学报,1999,18(5):507-512.
ZHENG W J,WANG X L,SHEN Y Y. A study on the ultra?structure of assimilative organs of some plants in saline habitate[J]. Journal of Chinese Electron Microscopy Society,1999,18(5):507-512.
[31] 刘卫国,丁俊祥,邹杰,等. NaCl 对齿肋赤藓叶肉细胞超微结构的影响[J]. 生态学报,2016,36(12):3556-3563.
LIU W G,DING J X,ZOU J,et al. Ultrastructural responsesof Syntrichia caninervis to a gradient of NaCl stress[J]. ActaEcologica Sinica,2016,36(12):3556-3563.
猜你喜欢
山东农业科学(2017年2期)2017-03-15
江苏农业科学(2016年8期)2017-02-15
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
吉林农业·下半月(2016年10期)2016-12-16
现代园艺(2016年2期)2016-03-15
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年9期)2015-10-20
