
龙眼DlSWEET1基因的克隆、表达及功能分析
2024-04-30 07:51包雨莹李韵江文洁谢涛方庭
果树学报 2024年4期
包雨莹 李韵 江文洁 谢涛 方庭


摘 要:【目的】SWEET(sugars will eventually be exported transporters)是一类参与植物生长发育多个过程的糖转运蛋白,分析DlSWEET1基因在龙眼不同组织和处理下的表达,探究其在果实糖积累中的功能。【方法】以龙眼松风本果实为材料,克隆DlSWEET1基因,采用实时荧光定量PCR分析DlSWEET1在龙眼不同组织器官的表达以及在激素、冷、热、干旱胁迫下的表达模式。通过亚细胞定位、糖转运活性分析及草莓瞬时转化研究DlSWEET1基因的功能。【结果】DlSWEET1基因开放阅读框(ORF)全长为750 bp,编码249个氨基酸,包含一个PQ-loop保守结构域和蛋白典型保守结构域 MtN3_slv。DlSWEET1在龙眼根、茎、叶、果肉等组织中均有不同程度的表达,在叶中的表达量较高,在果肉中的表达量次之,而在茎和根中表达量较低;不同浓度的蔗糖、葡萄糖和果糖处理龙眼叶片后,DlSWEET1在叶片中的表达量均有显著升高;低温、干旱及MeJA(茉莉酸甲酯)处理可显著提高DlSWEET1的表达。农杆菌侵染本氏烟草发现DlSWEET1蛋白定位在细胞膜和细胞核。糖转运活性分析证明DlSWEET1蛋白可以转运葡萄糖、果糖、蔗糖和甘露糖。草莓中瞬时转化DlSWEET1可以显著提升果实中的可溶性糖含量。【结论】瞬时过表达DlSWEET1导致转基因草莓果实的可溶性糖含量增加,为进一步解析DlSWEET1在龙眼果实糖积累中的作用提供理论依据。
关键词:龙眼;DlSWEET1;表达分析;亚细胞定位;糖积累
中图分类号:S667.2 文献标志码:A 文章编号:1009-9980(2024)04-0679-11
Cloning, expression and functional analysis of longan DlSWEET1 gene
BAO Yuying1, 2, LI Yun1, 2, JIANG Wenjie 1, 2, XIE Tao1, 2, FANG Ting1, 2*
( 1College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China; 2Institute of Horticultural Plant Genetics and Breeding, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China )
Abstract: 【Objective】 Longan (Dimocarpus longan L.) is one of the important economic fruit crops in southern China. Longan has the function of nourishing the heart, spleen and blood, and calming the mind of peaple. It has been regarded as a precious supplement since ancient times. The sugar content in fruits is a key factor affecting fruit quality, and improvement of fruit sugar content is of great significance for promoting the high-quality and efficient development of Chinas longan industry. The SWEET sugar transporters protein (SWEETs) not only plays an important role in plant stress and hormone response, but also plays a crucial role in the normal growth and development of plants, especially in promoting sugar accumulation. However, there has been limited research on the DlSWEETs in longan, especially on sugar accumulation. The purpose of this study is to screen and validate the functions of the candidate DlSWEETs that may be involved in sugar accumulation processes. 【Methods】 The CDS sequence of the DlSWEET1 was cloned using the cDNA of the preserved Songfengben fruit in the laboratory as a template. The DNAMAN software was used to translate the correctly sequenced DlSWEET1 gene nucleotide sequence into amino acid sequence, and its conserved domain was predicted by NCBI. The protein transmembrane domains were analyzed using the TMHMM2.0. We extracted the total RNA from different tissues (roots, stems, leaves, and fruits) of longan and leaf samples after different treatments, reversed the transcribe to obtain cDNA, and then used real-time fluorescence quantitative PCR (qRT-PCR) to detect the expression level of DlSWEET1 in different tissues and organs of longan, as well as its expression level under hormone, cold, heat, and drought stress. All the experiments were repeated three times in terms of biology and technology, and the relative expression levels of the genes were calculated using the 2–??CT method, and then statistically analyzed by t test. p<0.05 indicated significant difference, and the error line represented the standard deviation of three biological repeats. the CDS sequence of the DlSWEET1 was cloned and connected to the pMD18-T vector. Using this plasmid as a template, PCR amplification of the DlSWEET1 was performed using primers. The pH7LIC5.0-ccdBrc-N-eGFP vector enzyme was cleaved using Stu Ⅰ enzyme cleavage, the pSAK277 vector enzyme was cleaved +using EcoR Ⅰ and Hind Ⅲ enzyme cleavage, and the pDR196 vector enzyme was cleaved using Pst Ⅰ and Spe Ⅰ enzyme cleavage. The amplification product was inserted into the multi clone sites of each vector. The first two recombinant vectors were transformed into GV3101 strain, and the last recombinant vector was transformed into EBYVW4000 yeast strain. The lower epidermis of tobacco leaves was injected with Agrobacterium and cultured in the dark room (25 ℃) for 2 days. The distribution of green fluorescence was observed by confocal laser microscope. The strawberries injected with Agrobacterium were cultured under 16 h light/8 h darkness in a greenhouse at 25 ℃ for 9 days. The relative expression of the DlSWEET1 in strawberries and the determination of soluble sugar content were analyzed by qRT-PCR. The bacterial solutions with OD600 of 0.1, 0.01 and 0.001 were taken 5 ?L into various sugar substrate media, and the media were cultured in a constant temperature incubator at 28 ℃ for 2-3 days, and the growth was observed and recorded. The function of the DlSWEET1 gene was investigated by subcellular localization, sugar transport activity analysis and transient transformation of strawberry. 【Results】 The DlSWEET1 contained 750 bp of ORF (open reading frame) and encoded 249 amino acids, which contained a PQ-loop conserved domain and a protein typical conserved domain MtN3_slv. Further analysis indicated that DlSWEET1 protein contained seven transmembrane domains. The qRT-PCR analysis results showed that the DlSWEET1 was expressed in different tissues such as the roots, stems, leaves, and pulp, with higher expression levels in the leaves, followed by in the pulp, and lower expression levels in the stems and roots. After treating the leaves with different concentrations of sucrose, glucose, and fructose, the expression level of the DlSWEET1 showed varying degrees of increase. The expression level of the DlSWEET1 in the leaves treated with sucrose was significantly higher than that in the control group, but there was no significant difference among different concentrations. The expression of glucose increased significantly with the increase of glucose concentration. In the treatment of fructose, lower concentration (0.5 g·L-1) and higher concentration (5 g·L-1) could significantly increase the expression of the DlSWEET1. The expression of the DlSWEET1 was significantly increased under low temperature, drought and MeJA treatments, and significantly decreased under ABA treatment. However, there was no significant change in its expression after high temperature, 6-BA and GA3 treatments. The fluorescence signals of GFP were mainly concentrated and overlapped in the cell membrane and nucleus. The sugar transport activity analysis showed that the DlSWEET1 protein could transport glucose, fructose, sucrose and mannose. After transient transformation, the contents of sucrose, glucose and fructose in the strawberry were significantly higher than those in the control treatment, and the expression level of the DlSWEET1 was significantly increased. 【Conclusion】 The DlSWEET1 gene was cloned from longan fruit, and its expression level could be induced by different sugar components (sucrose, glucose, and fructose), stress (low temperature and drought), and hormones (MeJA and ABA). The subcellular localization revealed that the gene is localized on the cell membrane and nucleus. The analysis of sugar transport activity showed that it could transport various sugar components, such as sucrose, glucose, fructose and mannose, but could not transport the toxic substrate deoxyglucose. The transient overexpression of the DlSWEET1 resulted in increased soluble sugar content in the transgenic strawberry fruits. The transient conversion of this gene in the strawberry significantly increased the relative expression of the DlSWEET1. These results indicated that the DlSWEET1 has the function of promoting sugar accumulation in fruits of longan. The article would provide a theoretical reference for improving fruit quality of longan.
Key words: Longan (Dimocarpus longan); DlSWEET1; Expression analysis; Subcellular localization; Sugar accumulation
果实的糖含量是影响果实风味品质的重要因素。前人研究表明,植物通过光合作用合成蔗糖,源器官叶片中的蔗糖需经过韧皮部装载、长距离运输、韧皮部卸载后进入果实的液泡中进行储存,糖转运蛋白在这一系列过程中起到关键的介导糖类跨膜运输作用[1]。截至目前,在植物中鉴定出各种类型的糖转运蛋白,主要可以分为单糖转运蛋白(monosaccharide transporter-like,MST)、蔗糖转运蛋白(sucrose transporters,SUT)和SWEET(sugars will eventually be exported transporters)3种类型[2]。
SWEET是2010年发现的一类广泛存在于动植物中的具有糖转运功能的蛋白家族[3],参与植物生长发育、胁迫响应和果实糖积累等多个过程[4]。在野草莓中过表达白梨的PbSWEET4 基因会降低叶片中的叶绿素含量,加速叶片衰老[5];敲除拟南芥AtSWEET17基因后侧根减少,进而导致耐旱性降低[6];胡晓波等[7]发现在过表达CitSWEET11d基因的柑橘愈伤组织和番茄果实中蔗糖含量显著增加,表明CitSWEET11d 促进了蔗糖的积累。路静等[8]通过组织表达分析发现,MdSWEET1基因主要在苹果的茎和花中表达,在番茄中异位表达该基因可提高果实的蔗糖和果糖含量。
龙眼是重要的热带、亚热带常绿果树,已经有2000多年栽培历史,广泛种植于东南亚、 南亞、澳大利亚和美国夏威夷等地区。龙眼原产并盛产于中国,栽培面积与产量都居于世界首位[9],主要集中栽培于海南、广东、广西、福建等省份,为我国热区第四大水果[10]。龙眼果实口感和风味的影响因素较多,而起主导作用的是含糖量,糖的组分及其各组分含量的多少直接决定着果实风味的好坏,除此之外,糖类还是类胡萝卜素、有机酸和维生素等营养物质生成的基础原料[11]。由于龙眼栽培管理相对粗放,常导致果实品质欠佳,直接影响了产业的健康持续发展。笔者在课题组前期龙眼SWEET基因家族成员鉴定的基础上,从龙眼果实中克隆DlSWEET1基因,然后将此基因构建到eGFP载体上,利用激光共聚焦观察荧光信号,获得DlSWEET1基因的亚细胞定位。通过观察酵母的生长情况及表达情况来研究DlSWEET1的糖转运活性,并在草莓果实中瞬时过表达DlSWEET1研究其在果实糖积累中的功能,为龙眼高糖性状改良提供理论依据和基因资源。
1 材料和方法
1.1 试验材料与处理
本试验中所用的龙眼材料为4个月苗龄的红核子幼苗。在高温和低温胁迫中,幼苗分别在40 ℃和4 ℃下处理;通过浇灌PEG6000(20%)模拟干旱胁迫;在激素处理中,分别用50 mmol·L-1 ABA、50 mmol·L-1 GA3、75 mmol·L-1 6-BA和100 mmol·L-1 MeJA(茉莉酸甲酯)喷施叶片;在可溶性糖喷施处理中,分别用0.5、1和5 g·L-1的葡萄糖、果糖和蔗糖喷施叶片;对照为28 ℃生长的植株,所有处理均包含3次生物学重复,处理4 h后取相同位置的叶片并用液氮速冻后保存于-80 ℃冰箱。用于瞬时转化的为红颜草莓,用于亚细胞定位的烟草为本氏烟草,所有植物材料均种植于福建农林大学园艺学院遗传育种实验室的培养室中。用于基因克隆的松风本龙眼果实取自福建省农业科学院国家龙眼枇杷种质资源圃,取材时期为成熟期(花后120 d)。
1.2 龙眼DlSWEET1基因的克隆
在龙眼基因组中查询Dlo_004842.1,即DlSWEET1,下载编码区序列。以实验室保存的松风本龙眼果实cDNA为模板,设计特异性引物(引物序列5-ATGGATATCGCACATTTCATATTCG-3/5-CTACACTCCAAACCGTGACCCG-3)克隆得到DlSWEET1的CDS序列并连接至pMD18-T载体上进行测序。
1.3 龙眼DlSWEET1基因序列的生物信息学分析
使用DNAMAN软件将测序正确的DlSWEET1基因核苷酸序列翻译成氨基酸序列,保守结构域通过NCBI(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测;使用TMHMM2.0网站(https://services.healthtech.dtu.dk/services/TMHMM-2.0/)在线分析蛋白的跨膜结构域。利用MEGA7.0软件构建系统进化树,方法为邻接法,Bootstrap设置为1000。
1.4 龙眼DlSWEET1基因的表达分析
采用天根RNAprep Pure多糖多酚植物总RNA提取试剂盒(DP441)提取龙眼不同组织(根、茎、叶片和果实)及不同处理后的叶片样品的总RNA,利用TransScript? One-Step gDNA Removal and cDNA Synthesis SuperMix试剂盒反转录得到cDNA。使用全式金的PerfectStart? Green qPCR SuperMix,在荧光定量PCR仪(Roche,LightCycler 96)上进行基因表达量的检测。DlSWEET1基因的引物序列为5-CGGGCTCCTGATGCTTGT-3/5-TCTTGGTGTTGCCGTGCA-3,内参基因Actin引物序列为5-TGCTATCCTTCGGTTGGACC-3/5-CGGACGATTTCCCGTTCAG-3。所有试验都进行3次生物学及技术重复,基因相对表达量的计算方法选用的是2–??CT法。
1.5 龙眼DlSWEET1基因的亚细胞定位
以连有DlSWEET1基因CDS的pMD18-T载体为模板,使用引物(引物序列5-ACAAGGATTACGC-
CGAGGCCTATGGATATCGCACATTTCATATTCG-
3/5-ATATCATTAGGGAAGAGGCCTCTACACTCCAAACCGTGACCCG-3),利用2 × Taq Master Mix-V21.1(Vazyme Biotech,P111/P112)进行PCR扩增。选择StuⅠ酶将pH7LIC5.0-ccdB rc-N-eGFP载体进行酶切使其线性化,利用无缝克隆试剂盒ClonExpress? Ⅱ One Step(C112,南京诺唯赞)将扩增产物插入已切开的目标载体。将构建好的载体通过农杆菌注射转化烟草,注射后的烟草于培养室(25 ℃)暗培养2 d,再借助激光共聚焦显微镜观察绿色荧光的分布。
1.6 龙眼DlSWEET1基因的糖转运活性检测
以含有DlSWEET1基因CDS 的质粒为模板,使用引物(引物序列5-CTTGATATCGAATTCCTGCAGATGGATATCGCACATTTCATATTCG-3/5-TATACCCCAGCCTCGACTAGTCTACACTCCAA-ACCGTGACCCG-3)进行PCR扩增。用PstⅠ和SpeⅠ酶切将pDR196酵母表达载体进行酶切使其线性化,利用无缝克隆试剂盒ClonExpress? Ⅱ One Step(C112,南京诺唯赞)将扩增产物插入目标载体。将构建好的融合载体转化酿酒酵母菌株EBYVW4000。以OD600=1的菌液为原液,用ddH2O稀释,分别调节OD600为0.1、0.01和0.001,吸取5 ?L原液以及稀释过的菌液点入不同糖底物培养基,倒放在28 ℃恒温培养箱。经过2~3 d的培养后,观察酵母能否正常生长,并比较不同浓度菌液之间酵母的生长状态。
1.7 龙眼DlSWEET1基因在草莓果实中的瞬时过表达
以含有DlSWEET1基因CDS的质粒为模板,使用引物(引物序列5-TCCAAAGAATTCAAAAAGCTTATGGATATCGCACATTTCATATTCG-3/5-TCAT-
TAAAGCAGGACTCTAGACTACACTCCAAACCGTGACCCG-3)进行PCR扩增。选择EcoRⅠ和Hind Ⅲ将pSAK277载体线性化,利用无缝克隆试剂盒ClonExpress? Ⅱ One Step(C112,南京诺唯赞)将扩增产物插入目标载体。将构建好的融合载体瞬时转化白果期红颜草莓果肉,具体操作方法参考Cheng等[12]的方法。注射农杆菌后的草莓于25 ℃温室培养,光照情况为16 h光照/8 h黑暗,9 d后取草莓果肉样品进行qRT-PCR分析以及糖含量测定。
2 结果与分析
2.1 DlSWEET1基因的克隆与生物学信息分析
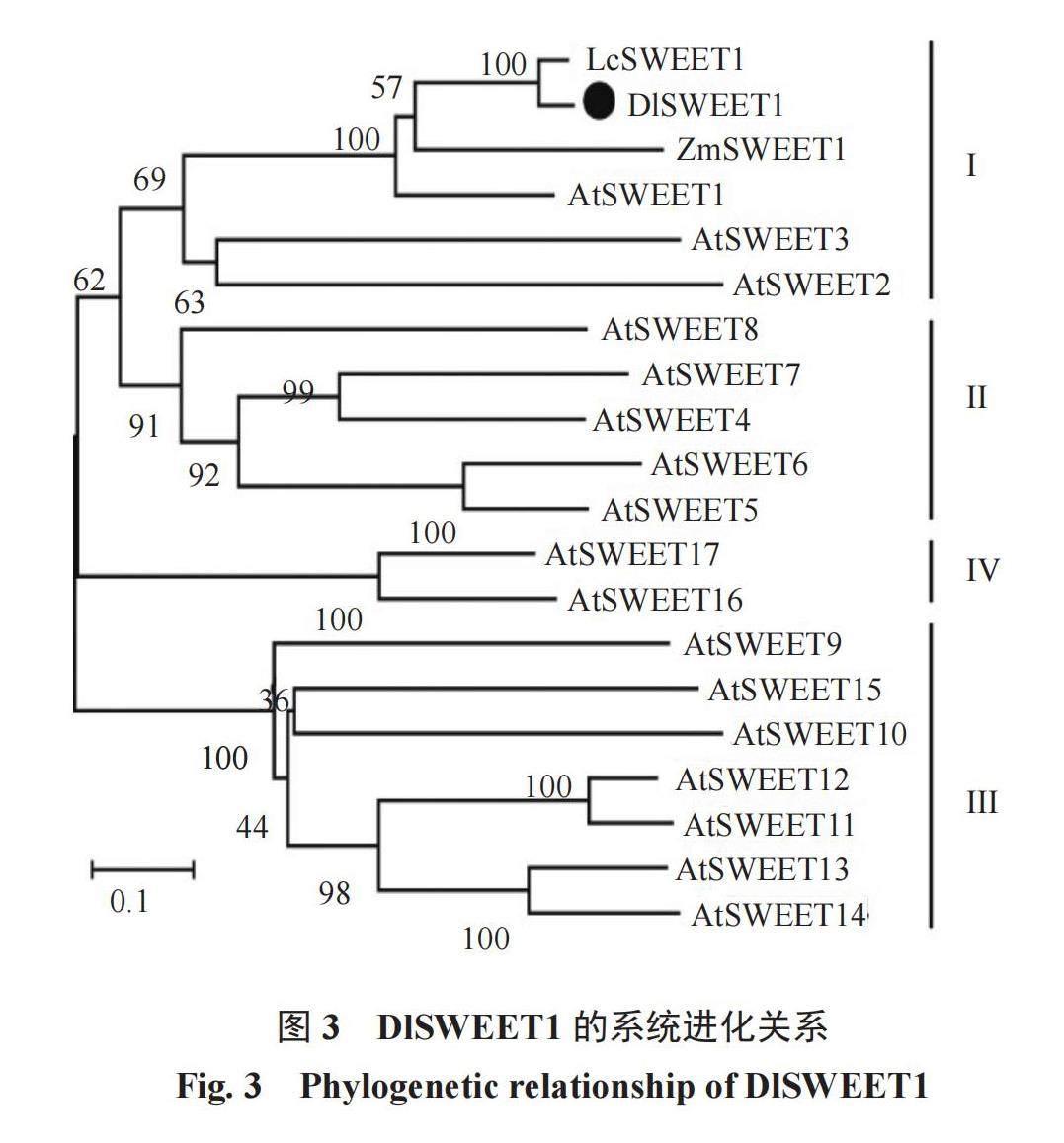
从松风本果实cDNA中克隆得到一条长度为750 bp左右的单一条带。将此条带进行胶回收得到目的片段,用pMD18-T载体与胶回收产物进行连接转化,大肠杆菌PCR鉴定结果表明,扩增到与目的片段大小一致的条带,且与基因组中的序列完全一致(图1)。蛋白保守结构分析表明,DlSWEET1蛋白在6~94位氨基酸之间含有1个PQ-loop保守结构域,在131~213位氨基酸之间含有1个MtN3_slv结构域(图2-A)。跨膜结构域分析表明,DlSWEET1蛋白含有7个跨膜结构域(图2-B)。系统进化分析表明,DlSWEET1与LcSWEET1[13]、ZmSWEET1[14]和AtSWEET1为直系同源,并与AtSWEET2和AtSWEET3同属于SWEETⅠ类(图3)。
2.2 DlSWEET1基因在龙眼不同组织器官中的表达分析
利用qRT-PCR检测DlSWEET1基因在龙眼根、茎、叶和果肉中的表达模式,结果表明,DlSWEET1基因在上述的龙眼组织部位中均有一定的表达,但表达模式有所差异。通过比较分析发现,DlSWEET1基因的表达量在叶中最高,其次是在果肉和根中,在茎中最低(图4)。
2.3 不同浓度糖处理下龙眼DlSWEET1基因表达模式
为了研究DlSWEET1基因对可溶性糖的响应,对龙眼叶片进行了不同浓度葡萄糖、果糖和蔗糖喷施处理,并进行DlSWEET1的表达情况检测。结果表明,DlSWEET1基因的表达量在叶片喷施糖处理后均呈现不同程度的上升趋势,其中喷施蔗糖处理后期表达量相较于对照都有显著上升(图5-A);葡萄糖处理后期DlSWEET1基因表达量随着浓度上升呈现显著上升趋势(图5-B);在果糖处理中,较低质量浓度(0.5 g·L-1)和较高质量浓度(5 g·L-1)均可显著提高DlSWEET1基因表达量(图5-C)。
2.4 不同胁迫及激素处理下龙眼DlSWEET1基因的表达分析
为了进一步探讨DlSWEET1基因是否响应激素及胁迫处理,利用qRT-PCR方法分析了该基因在不同激素及胁迫处理下的表达情况。结果表明,DlSWEET1基因的表达量在低温、干旱及MeJA处理下呈现显著上升趋势;在ABA处理下表达量显著下降。然而,在高温、6-BA和GA3处理后期其表达量没有显著变化(图6)。
2.5 DlSWEET1亚细胞定位分析
为进一步明确DlSWEET1蛋白的亚细胞定位情况,笔者通过农杆菌瞬时转化了本氏烟草叶片。激光共聚焦定位观察发现,DlSWEET1蛋白定位在植物细胞的细胞膜和细胞核中(图7)。
2.6 DlSWEET1的糖转运活性分析
将酵母表达载体成功转入EBYVW4000酿酒酵母感受态细胞中,观察酵母菌在含有不同底物(葡萄糖、果糖、甘露糖、蔗糖、麦芽糖和脱氧葡萄糖)的SD(-ura)固體培养基中能否正常生长及不同浓度菌液下的生长状态(图8)。转基因酵母在含有葡萄糖、果糖、甘露糖、蔗糖及麦芽糖和脱氧葡萄糖为共同底物的培养基上能正常生长,说明DlSWEET1蛋白对某些糖有转运能力,能转运的糖组分包括葡萄糖、果糖、蔗糖、甘露糖,但没有能力转运脱氧葡萄糖这种毒性底物。
2.7 瞬时过表达DlSWEET1基因对草莓果实可溶性糖含量的影响
为验证DlSWEET1基因在果实糖积累中的功能,利用草莓瞬时转化体系在红颜草莓白果期果实中过表达DlSWEET1基因。qRT-PCR结果(图9)表明,相对于对照,DlSWEET1基因的表达量在瞬时转化果实中显著升高(图9-A)。此外,经瞬时转化后草莓果实中的蔗糖、葡萄糖和果糖的含量均显著高于对照(图9-B~D)。
3 讨 论
作为韧皮部糖类装载的重要参与者,近年来大量研究表明,SWEET糖转运蛋白参与果实糖转运,影响果实中的可溶性糖含量[15-17]。课题组此前已从龙眼基因组中筛选鉴定出龙眼的SWEET基因家族成员[2]。笔者选取DlSWEET1基因,并对其糖转运功能及其调控机制进行研究。
有研究证明植物SWEET基因的表达具有组织差异,这种差异可能在其所调控的很多植物生理代谢过程中都起着决定性作用[18-19]。PwSWEET1基因在云杉花粉和花粉管中特异性表达,蔗糖和葡萄糖可以诱导PwSWEET1表达,PwSWEET1还可以恢复酵母菌EBY对葡萄糖的吸收[20];SWEET1基因在拟南芥花器官优先表达,在体外鉴定具有葡萄糖转运活性[21];MeSWEET1在木薯不同组织器官中的相对表达量不同,其中在成熟叶片中最高,而在新叶和果实中最低[22]。笔者在本研究中利用qRT-PCR检测了龙眼根、茎、叶、果肉的相对表达量,发现DlSWEET1在龙眼叶片中表达量最高,其次是果肉,而在根和茎中表达量较低,说明DlSWEET1具有组织特异性,在叶片和果肉中表达量较高说明其可能参与了龙眼叶片发育及果实的糖积累。糖转运活性分析表明DlSWEET1可以转运葡萄糖、蔗糖、果糖和甘露糖,推测该基因可能通过在龙眼不同组织器官中转运不同种类的糖,进而参与多种植物生长发育过程。
除了参与植物生长发育外,SWEET糖转运蛋白在植物胁迫和激素响应方面还发挥重要作用[23-24]。大蒜AsSWEET14基因参与逆境胁迫,在干旱和低温胁迫下均显著上调表达[25];小黑麦多个TwSWEETs基因在干旱或低温胁迫下呈现出显著差异表达[26];在苹果中,ABA可能会通过调节MdWRKY9-MdSWEEET9b途径来影响果实糖的积累[27]。笔者在本研究中发现在低温和干旱处理下,龙眼DlSWEET1的表达量显著上升,与前人在6个月苗龄红核子龙眼幼苗上的研究结果相一致[2],但6个月苗龄条件下该基因的表达量上升幅度较4个月苗龄条件下大,推测该基因可能在龙眼幼苗发育的较长时间中都会参与低温和干旱胁迫响应,但不同发育时期的作用大小有所差异。此外,DlSWEET1的表达量在MeJA处理下呈现显著上升趋势,与前人关于该基因启动子具备多个激素响应元件的结果相一致[2]。
果实中的糖积累高度依赖于糖转运体[28]。越来越多的研究表明,SWEET蛋白可以促进可溶性糖的积累。有研究发现 LcSWEET10的表达模式与荔枝假种皮中糖的积累呈正相关[29],异源表达IbSWEET15基因可提高拟南芥种子中的可溶性糖含量[30]。为了验证DlSWEET1在龙眼果实糖积累中的功能,笔者通过农杆菌注射瞬时侵染草莓,检测出转基因草莓的可溶性糖含量显著上升,说明DlSWEET1在促进龙眼糖分积累中起重要作用,为后期深入研究龙眼糖积累调控机制提供理论依据。
4 结 论
從龙眼果实中克隆得到DlSWEET1基因,其表达量可被不同糖组分(蔗糖、葡萄糖和果糖)、逆境(低温和干旱)和激素(MeJA和ABA)等条件诱导。亚细胞定位发现DlSWEET1基因定位于细胞膜和细胞核。糖转运活性分析表明其可以转运蔗糖、葡萄糖、果糖和甘露糖等多种糖组分。草莓瞬时转化该基因可显著提高草莓果实的可溶性糖含量。基于上述研究结果,初步推测DlSWEET1具有促进龙眼果实糖积累的功能,为龙眼果实品质改良提供了一定的理论参考。
参考文献References:
[1] YAMAKI S. Metabolism and accumulation of sugars translocated to fruit and their regulation[J]. Journal of the Japanese Society for Horticultural Science,2010,79(1):1-15.
[2] FANG T,RAO Y,WANG M Z,LI Y,LIU Y J,XIONG P P,ZENG L H. Characterization of the SWEET gene family in longan (Dimocarpus longan) and the role of DlSWEET1 in cold tolerance[J]. International Journal of Molecular Sciences,2022,23(16):8914.
[3] CHEN L Q,HOU B H,LALONDE S,TAKANAGA H,HARTUNG M L,QU X Q,GUO W J,KIM J G,UNDERWOOD W,CHAUDHURI B,CHERMAK D,ANTONY G,WHITE F F,SOMERVILLE S C,MUDGETT M B,FROMMER W B. Sugar transporters for intercellular exchange and nutrition of pathogens[J]. Nature,2010,468(7323):527-532.
[4] 张计育,王刚,王涛,贾展慧,宣继萍. SWEET蛋白在植物生长发育中的功能作用研究进展[J]. 植物资源与环境学报,2023,32(5):1-15.
ZHANG Jiyu,WANG Gang,WANG Tao,JIA Zhanhui,XUAN Jiping. Research progress on functional roles of SWEET proteins in plant growth and development[J]. Journal of Plant Resources and Environment,2023,32(5):1-15.
[5] NI J P,LI J M,ZHU R X,ZHANG M Y,QI K J,ZHANG S L,WU J. Overexpression of sugar transporter gene PbSWEET4 of pear causes sugar reduce and early senescence in leaves[J]. Gene,2020,743:144582.
[6] VALIFARD M,LE HIR R,M?LLER J,SCHEURING D,NEUHAUS H E,POMMERRENIG B. Vacuolar fructose transporter SWEET17 is critical for root development and drought tolerance[J]. Plant Physiology,2021,187(4):2716-2730.
[7] 胡晓波. 柑橘SWEET11d对果实蔗糖积累的转录调控机制研究[D]. 杭州:浙江大学,2021.
HU Xiaobo. Transcriptional regulation mechanism of citrus SWEET11d on fruit sucrose accumulation[D]. Hangzhou:Zhejiang University,2021.
[8] 路静,马齐军,康慧,李文浩,刘亚静,郝玉金,由春香. 苹果糖转运蛋白基因MdSWEET1在番茄中异源表达提高其耐盐性[J]. 园艺学报,2019,46(3):433-443.
LU Jing,MA Qijun,KANG Hui,LI Wenhao,LIU Yajing,HAO Yujin,YOU Chunxiang. Ectopic expressing MdSWEET1 in tomato enhanced salt tolerance[J]. Acta Horticulturae Sinica,2019,46(3):433-443.
[9] LIN Y L,MIN J M,LAI R L,WU Z Y,CHEN Y K,YU L L,CHENG C Z,JIN Y C,TIAN Q L,LIU Q F,LIU W H,ZHANG C G,LIN L X,ZHANG D M,THU M,ZHANG Z H,LIU S C,ZHONG C S,FANG X D,WANG J,YANG H M,VARSHNEY R K,YIN Y,LAI Z X. Genome-wide sequencing of longan (Dimocarpus longan Lour.) provides insights into molecular basis of its polyphenol-rich characteristics[J]. GigaScience,2017,6(5):1-14.
[10] 齊文娥,陈厚彬,彭朵芬,晏发发. 中国龙眼产业发展现状、问题与对策建议[J]. 广东农业科学,2016,43(8):169-174.
QI Wene,CHEN Houbin,PENG Duofen,YAN Fafa. Present situation,problems and suggestions of the development of Chinese longan industry[J]. Guangdong Agricultural Sciences,2016,43(8):169-174.
[11] LUO T,SHUAI L,LIAO L Y,LI J,DUAN Z H,GUO X M,XUE X Q,HAN D M,WU Z X. Soluble acid invertases act as key factors influencing the sucrose/hexose ratio and sugar receding in longan (Dimocarpus longan Lour.) pulp[J]. Journal of Agricultural and Food Chemistry,2019,67(1):352-363.
[12] CHENG J T,WEN S Y,XIAO S,LU B Y,MA M R,BIE Z L. Overexpression of the tonoplast sugar transporter CmTST2 in melon fruit increases sugar accumulation[J]. Journal of Experimental Botany,2018,69(3):511-523.
[13] XIE H H,WANG D,QIN Y Q,MA A N,FU J X,QIN Y H,HU G B,ZHAO J T. Genome-wide identification and expression analysis of SWEET gene family in Litchi chinensis reveal the involvement of LcSWEET2a/3b in early seed development[J]. BMC Plant Biology,2019,19(1):499.
[14] 李明,郑德志,郝立冬,郭海滨,孙莹莹. 玉米SWEET基因家族的全基因组鉴定与分析[J/OL]. 分子植物育种,2022:1-11. (2022-10-31). http://kns.cnki.net/kcms/detail/46.1068.S.20221031.1009.008.html.
LI Ming,ZHENG Dezhi,HAO Lidong,GUO Haibin,SUN Yingying. Genome-wide identification and analysis of SWEET gene family in Maize[J/OL]. Molecular Plant Breeding,2022:1-11. (2022-10-31). http://kns.cnki.net/kcms/detail/46.1068.S.20221031.1009.008.html.
[15] ZHANG Z,ZOU L M,REN C,REN F R,WANG Y,FAN P G,LI S H,LIANG Z C. VvSWEET10 mediates sugar accumulation in grapes[J]. Genes,2019,10(4):255.
[16] GENG Y Q,WU M J,ZHANG C M. Sugar transporter ZjSWEET2.2 mediates sugar loading in leaves of Ziziphus jujuba Mill.[J]. Frontiers in Plant Science,2020,11:1081.
[17] LI X Y,GUO W,LI J C,YUE P T,BU H D,JIANG J,LIU W T,XU Y X,YUAN H,LI T,WANG A D. Histone acetylation at the promoter for the transcription factor PuWRKY31 affects sucrose accumulation in pear fruit[J]. Plant Physiology,2020,182(4):2035-2046.
[18] KO H Y,HO L H,NEUHAUS H E,GUO W J. Transporter SlSWEET15 unloads sucrose from phloem and seed coat for fruit and seed development in tomato[J]. Plant Physiology,2021,187(4):2230-2245.
[19] JEENA G S,KUMAR S,SHUKLA R K. Structure,evolution and diverse physiological roles of SWEET sugar transporters in plants[J]. Plant Molecular Biology,2019,100(4/5):351-365.
[20] ZHOU Y N,CUI X Y,HU A N,MIAO Y H,ZHANG L Y. Characterization and functional analysis of pollen-specific PwSWEET1 in Picea wilsonii[J]. Journal of Forestry Research,2020,31(5):1913-1922.
[21] 辛红佳,李鹏程,滕守振,李圣彦,汪海,郎志宏. 拟南芥SWEET1/2/3基因突变体构建及功能鉴定[J]. 中国农业科技导报,2020,22(2):39-49.
XIN Hongjia,LI Pengcheng,TENG Shouzhen,LI Shengyan,WANG Hai,LANG Zhihong. Construction and functional characterization of mutants of Arabidopsis SWEET1/2/3 genes[J]. Journal of Agricultural Science and Technology,2020,22(2):39-49.
[22] 劉秦,马畅,冯世鹏,唐枝娟,陈银华,罗丽娟,牛晓磊. 木薯SWEET1基因的分子克隆、亚细胞定位与功能分析[J]. 分子植物育种,2017,15(7):2502-2509.
LIU Qin,MA Chang,FENG Shipeng,TANG Zhijuan,CHEN Yinhua,LUO Lijuan,NIU Xiaolei. Molecular cloning,subcellular localization and function analysis of a MeSWEET1 gene from Manihot esculenta[J]. Molecular Plant Breeding,2017,15(7):2502-2509.
[23] CHANDRAN D. Co-option of developmentally regulated plant SWEET transporters for pathogen nutrition and abiotic stress tolerance[J]. IUBMB Life,2015,67(7):461-471.
[24] 孙文杰,左开井. SWEET转运蛋白家族的发现、结构及功能研究进展[J]. 分子植物育种,2016,14(4):878-885.
SUN Wenjie,ZUO Kaijing. The finding of SWEET transporter family and the research advance on its structure and function[J]. Molecular Plant Breeding,2016,14(4):878-885.
[25] 郑晓雯,徐庭亮,田洁. 大蒜糖转运蛋白基因AsSWEET14克隆与胁迫表达分析[J]. 植物生理学报,2023,59(8):1555-1565.
ZHENG Xiaowen,XU Tingliang,TIAN Jie. Cloning and expression analysis of sugar transporter gene AsSWEET14 under stress in garlic[J]. Plant Physiology Journal,2023,59(8):1555-1565.
[26] 李根,牛奎举. 小黑麦SWEET家族基因鉴定及其在不同逆境下表达模式分析[J]. 草地学报,2023,31(11):3310-3321.
LI Gen,NIU Kuiju. Characterization and analysis of SWEET gene family and response to abiotic stress in Triticosecale[J]. Acta Agrestia Sinica,2023,31(11):3310-3321.
[27] 张淑辉. MdWRKY9-MdSWEET9b响应ABA信号调控苹果果实糖积累的机理研究[D]. 泰安:山东农业大学,2023.
ZHANG Shuhui. Mechanisms of MdWRKY9-MdSWEET9b regulating sugar accumulation in apple fruits in response to ABA signal[D]. Taian:Shandong Agricultural University,2023.
[28] CHEN T,ZHANG Z Q,LI B Q,QIN G Z,TIAN S P. Molecular basis for optimizing sugar metabolism and transport during fruit development[J]. aBIOTECH,2021,2(3):330-340.
[29] 谢涵涵. 荔枝SWEET基因家族的克隆及其功能初步分析[D]. 广州:华南农业大学,2020.
XIE Hanhan. Cloning and preliminary functional analysis of SWEET gene family in Litchi Chinensis[D]. Guangzhou:South China Agricultural University,2020.
[30] 吳旭莉,吴正丹,晚传芳,杜叶,高艳,李賾萱,王志前,唐道彬,王季春,张凯. 甘薯糖转运蛋白IbSWEET15的功能研究[J]. 作物学报,2023,49(1):129-139.
WU Xuli,WU Zhengdan,WAN Chuanfang,DU Ye,GAO Yan,LI Zexuan,WANG Zhiqian,TANG Daobin,WANG Jichun,ZHANG Kai. Functional identification of sucrose transporter protein IbSWEET15 in sweet potato[J]. Acta Agronomica Sinica,2023,49(1):129-139.
猜你喜欢
小学生必读(低年级版)(2022年9期)2022-02-09
作文大王·低年级(2019年4期)2019-05-13
世界热带农业信息(2018年5期)2018-11-09
世界热带农业信息(2018年6期)2018-03-05
中国中药杂志(2017年1期)2017-03-06
岭南音乐(2016年5期)2017-01-17
中国中药杂志(2016年20期)2016-11-19
学苑创造·A版(2016年9期)2016-10-10
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年5期)2015-10-20
