
梨果实石细胞木质素合成调控机制研究进展
2024-04-30 09:34王红宝王永博王晋李勇李晓王迎涛王亚茹
果树学报 2024年4期
关键词:木质素
王红宝 王永博 王晋 李勇 李晓 王迎涛 王亚茹
摘 要:石細胞是由木质素沉积形成的厚壁组织细胞,是制约梨果实品质提升的重要因素。梨果肉石细胞的形成与木质素的生物合成、转移和沉积密切相关。因此,探究梨果实发育过程中木质素合成调控机制对调节石细胞形成及果实品质改良具有重要意义。总结近年来与梨果实木质素合成调控机制有关的研究,对转录因子、激素、糖、钙、活性氧、光质及花粉直感等调控因子在梨果实木质素合成中的作用机制进行综述,旨在为梨果实木质素的调控网络深入研究及品质改良提供参考。
关键词:梨;石细胞;木质素;调控机制
中图分类号:S661.2 文献标志码:A 文章编号:1009-9980(2024)04-0750-14
Research progress in the regulation mechanism of lignin synthesis in pear stone cells
WANG Hongbao, WANG Yongbo, WANG Jin*, LI Yong, LI Xiao, WANG Yingtao, WANG Yaru*
(Shijiazhuang Institute of Fruit Trees, Hebei Academy of Agriculture and Forestry Sciences, Shijiazhuang 050061, Hebei, China)
Abstract: Stone cells are sclerenchyma cells formed by deposition of lignin, which is the most significant factor limiting the quality of pears. Therefore, it is of great significance to explore the mechanism of lignin synthesis regulation in pear fruit development for the genetic regulatory network of stone cell traits. In this paper, the mechanisms of transcriptional factors, hormones, sugars, calcium, reactive oxygen species, light quality and pollen sensitivity in lignin synthesis in pear fruits are reviewed, aiming to provide reference for the regulatory network of lignin in pear fruits and genetic improvement of fruit quality. Several transcription factors involved in lignin biosynthesis have been identified in pears, such as MYB, NAC, bZIP, KNOX and zinc finger protein, among which MYB and NAC transcription factors play key regulatory roles in lignin synthesis. Study has showed that PbrMYB169, an R2R3MYB transcription factor of Pyrus bretschneideri, positively regulates lignification of stone cells in pear fruits. On the one hand, PbrMYB24 activates the transcription of lignin and cellulose biosynthesis genes by binding to different cis-element. On the other hand, PbrMYB24 binds directly to the promoters of PbrMYB169 and PbrNSC, activating the gene expression. Moreover, both PbrMYB169 and PbrNSC activate the promoter of PbrMYB24, enhancing gene expression. Research has identified PbMYB61 and PbMYB308 as candidate transcriptional regulators of stone cell formation, revealing that PbMYB61 regulates stone cell lignin formation by binding to the AC element in the PbLAC1 promoter to upregulate expression. Exogenous application of 200 μmol·L-1 NAA can reduce stone cell content and also significantly decrease the expression level of PbrNSC encoding a transcriptional regulator. In addition, PbrARF13-PbrNSC-PbrMYB132 regulatory cascade mediates the biosynthesis of lignin and cellulose in stone cells of pear fruit in response to auxin signals. Research has found several hormone-responsive elements in the upstream regulatory sequences of PbPALs family members. ABA, SA and MeJA could regulate the expression of PbPAL1 and PbPAL3 genes, and affect the formation of fruit stone cells. A series of experiments have proved that PbUGT72AJ2 mediates glycosylation by catalyzing the glucose conjugation of monolignols and may affect the expression of downstream genes as well as the content of monolignols to affect the lignin deposition and stone cell development in pear fruit. The treatment of exogenous glucose has significantly enhanced the accumulation of lignin in pear calli. Expression of structural genes (PbPAL, PbHCT, PbCOMT and PbPRX) in lignin biosynthesis is up-regulated after glucose treatment. Transien expression of PbPFP has resulted in a significant increase of lignin content in Dangshansuli fruits on 35th day after full bloom (DAB) and in tobacco leaves, indicating that PbPFP might be associated with the enhancement of lignin biosynthesis in response to glucose treatment. Ca2+ is known to inhibit stone cells in pear fruits. In order to further explore how calcium-nitrate treatment affects lignin synthesis, the PbCML3 has been identified in pears and relevant experiments have been conducted to find that the overexpression of PbCML3 would increase the content of pear stone cell. Further analysis has identified a transcription factor, PuDof2.5, and its targets gene PuPRX42- like (lignin polymerase gene) expression has decreased in CaCl2-treated samples, which are involved in suppressing lignin biosynthesis in pear fruit. ROS is closely associated with lignin deposition and stone cell formation. Research has showed that PuRBOHF, an RBOH isoform, plays an important role in secondary wall formation in pear stone cells. Inhibitors of RBOH activity suppress ROS accumulation and stone cell lignification in pear fruit. Moreover, it has been showed that PuMYB169 regulates PuRBOHF expression, while PuRBOHF-derived ROS induces the transcription of PuPOD2 and PuLAC2. Research has showed that secondary cell wall thickening and lignin accumulation in pears may regulate by different wavelengths of light. It is reported that CRY-mediated blue-light signal plays an important role in cell wall lignification and promotes the formation of stone cells in pears by regulating downstream genes. Results have showed that blue light induces the expression of lignin structure genes and promotes lignin accumulation. Furthermore, four blue light receptors cryptochromes have been identified in white pear, named PbCRY1a, PbCRY1b, PbCRY2a and PbCRY2b. Previous studies have reported that pollination affects the expression of laccase gene microRNA in pear fruits, and the expression of peroxidase 47 (PER47), β-glucosidase (BGLU15) and laccase-4 (LAC4), thus affecting lignin synthesis. This finding demonstrates that pollination with different sources of pollens affects the synthesis of lignin in pear fruit on the levels of gene and protein expression.
Key words: Pear; Stone cell; Lignin; Regulation mechanism
梨(Pyrus)是中国乃至世界范围内重要的水果之一,爽脆可口,香甜多汁,深受消费者的喜欢[1]。梨果实品质受多种因素的综合影响,其中一类木质化的厚壁组织细胞——石细胞便是重要的因素之一[2]。石细胞含量过高会导致果肉粗糙、口感低劣、硬度增加,同时也会影响梨果实糖分、有机酸、维生素含量等品质指标[3]。石细胞由薄壁组织细胞在其初生壁上沉积木质素等而形成。木质素在细胞壁外层开始发生,逐步向质膜发展,以分散颗粒状沿纤维素微纤丝的方向分层沉积,交替排列,直到充满整个细胞腔,形成石细胞[4]。木质素由多个木质素单体聚合而成,其生物合成过程由苯丙氨酸起始,经过一系列羟基化、甲基化、连接和还原反应生成木质素单体,木质素单体被运送到外质体氧化聚合形成愈创木基木质素(Guaiacyl lignin,G-型木质素)、对-羟基苯基木质素(Hydroxy-phenyl lignin,H-木质素)和丁香基木质素(Syringyl lignin,S-木质素)[5]。梨果实石细胞木质素属于G-S木质素,充分发育的梨石细胞含有约40%的木质素[6]。因此,调控木质素的积累是降低梨果实石细胞含量的关键。
目前,梨果实木质素生物合成的调控机制不断被解析,其合成调控过程受到信号途径、环境因素等多重因子的调控,要实现从分子层面抑制木质素的合成需要更为完整的理论支撑。为此,笔者围绕近些年梨果实木质素生物合成调控机制研究(转录因子、激素、糖、钙、活性氧、光质、花粉直感)展开综述,旨在为深入研究梨果实木质素合成调控网络奠定理论基础,为通过生物技术手段进行梨果实品质的遗传改良提供参考。
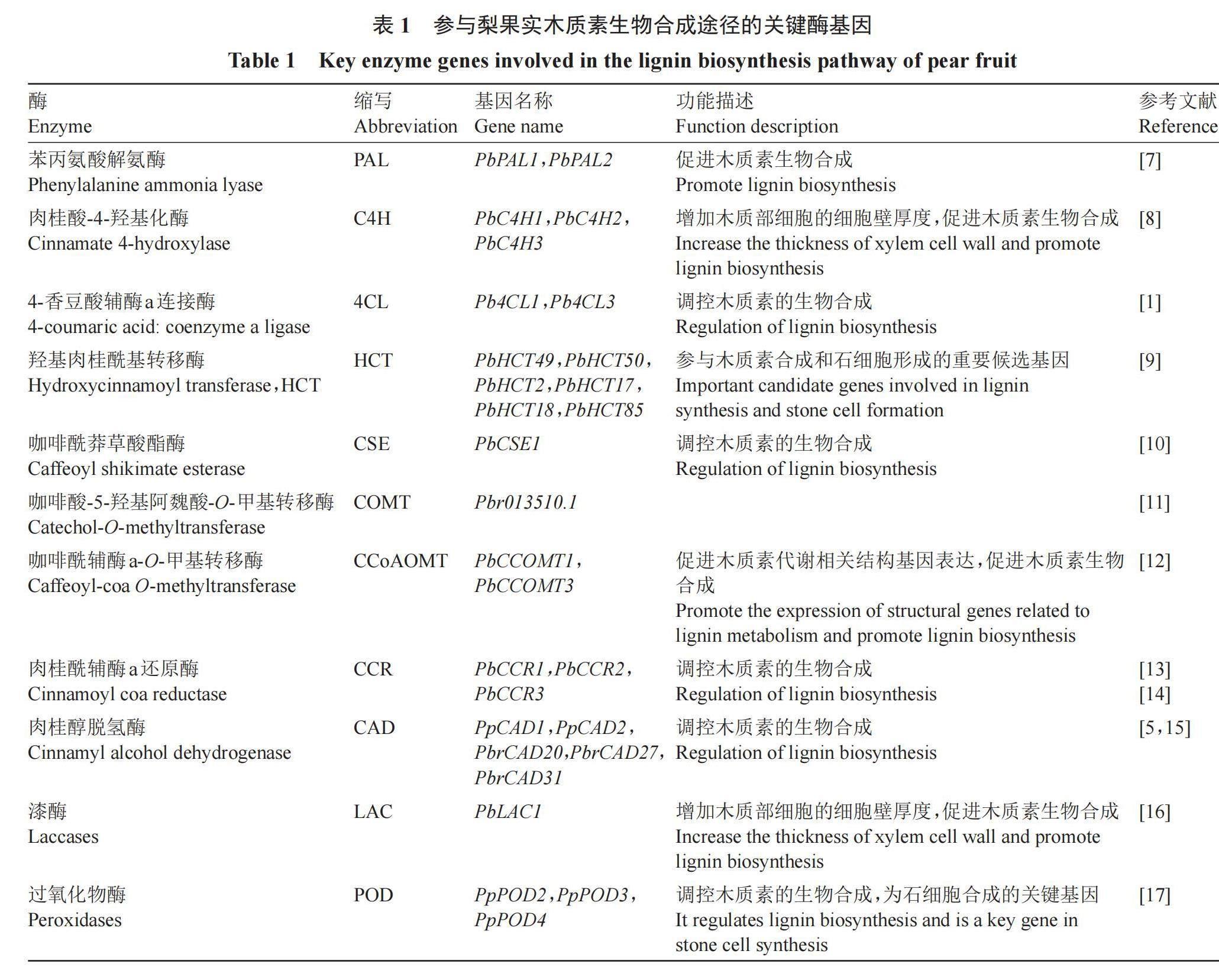
1 梨果实木质素合成相关的酶基因
木质素的生物合成由PAL、C4H、4CL、HCT、C3H、CSE、COMT、CCoAOMT、CCR等酶催化完成(表1)[7-17]。其中PAL和4CL是调控木质素含量的关键酶,C3H、CCoAOMT和COMT是木质素特异途径中的关键酶,CCR、CAD和POD是木质素单体合成及聚合的关键酶。PAL是木质素代谢途径中的关键酶和限速酶,位于苯丙烷代谢途径的入口[18]。在梨果实的不同发育阶段,PbPAL1和PbPAL2表现出与木质素含量一致的变化趋势,并与木质素生物合成的关键基因表达模式相似,在拟南芥中过表达结果显示随着木质素含量的增加,维管间纤维和木质部细胞的细胞壁增厚[7]。作为苯丙烷代谢途径中的第2步关键酶C4H,PbC4H1、PbC4H2和PbC4H3不仅可以增加维管间纤维和木质部细胞的木质素含量,还可以增加细胞壁厚度[8,19]。
在砀山酥梨基因组中鉴定到29个4CL基因和82个HCT基因,其中Pb4CL1和Pb4CL3可能参与了梨果实木质素合成,PbHCT49、PbHCT50、PbHCT2、PbHCT17、PbHCT18和PbHCT85的基因表达与梨果实石细胞表达模式相符,被鉴定为参与石细胞形成的重要候选基因,4CL活性抑制和HCT基因转录末端序列的缺失都会导致木质素含量降低[1,9]。Xu等[10]认为PbCSE1与木质素沉积和石细胞形成有关,过表达PbCSE1可提高梨果实木质素含量,并且参与木质素生物合成的相关基因的表达量也有所增加。为鉴定梨OMT基因家族,Cheng等[12]对26个OMT基因进行时空表达分析,其中PbCCOMT1和PbCCOMT3的启动子区域都含有木质素合成的调控基序,并且在梨果实发育过程中PbCCOMT1和PbCCOMT3的转录水平与石细胞和木质素含量呈正相关。COMT主要参与S-木质素的合成,催化G-木质素转化为S-木质素,20世纪梨中COMT基因与木质素含量呈正相关,与石细胞形成有很大相关性[11]。
Cheng等[13]从梨基因组中挖掘出31个CCR基因和26个CAD基因,构建了CCRs和CADs的系统发育树,通过实时定量聚合酶链式反应(qRT-PCR)明确了PbCAD2、PbCCR1、PbCCR2和PbCCR3参与了梨果实中木质素的合成[14]。其中PbCCR1在砀山酥梨木质素的生物合成中起着积极的作用,过表达PpCAD2具有更高的木质素含量和CAD酶活性,CAD活性下降可以改变木质素结构,而对木质素总量没有显著影响[5]。此外,研究发现,PbrCAD1、PbrCAD20、PbrCAD27和PbrCAD31的表达模式与梨果实发育过程中石细胞含量的变化相似[15]。在Wangkumbae梨发育过程中PpPOD1与木质素变化趋势一致,PpPOD2、PpPOD3、PpPOD4等基因可能是梨果实中石细胞合成的关键基因[17]。将Pyr-miR1890的两个靶基因PbLAC1和PbLAC14进行遗传转化,过表达PbLAC1可以显著增加木质素含量和木质部细胞的细胞壁厚度,参与了木质素的生物合成[16]。
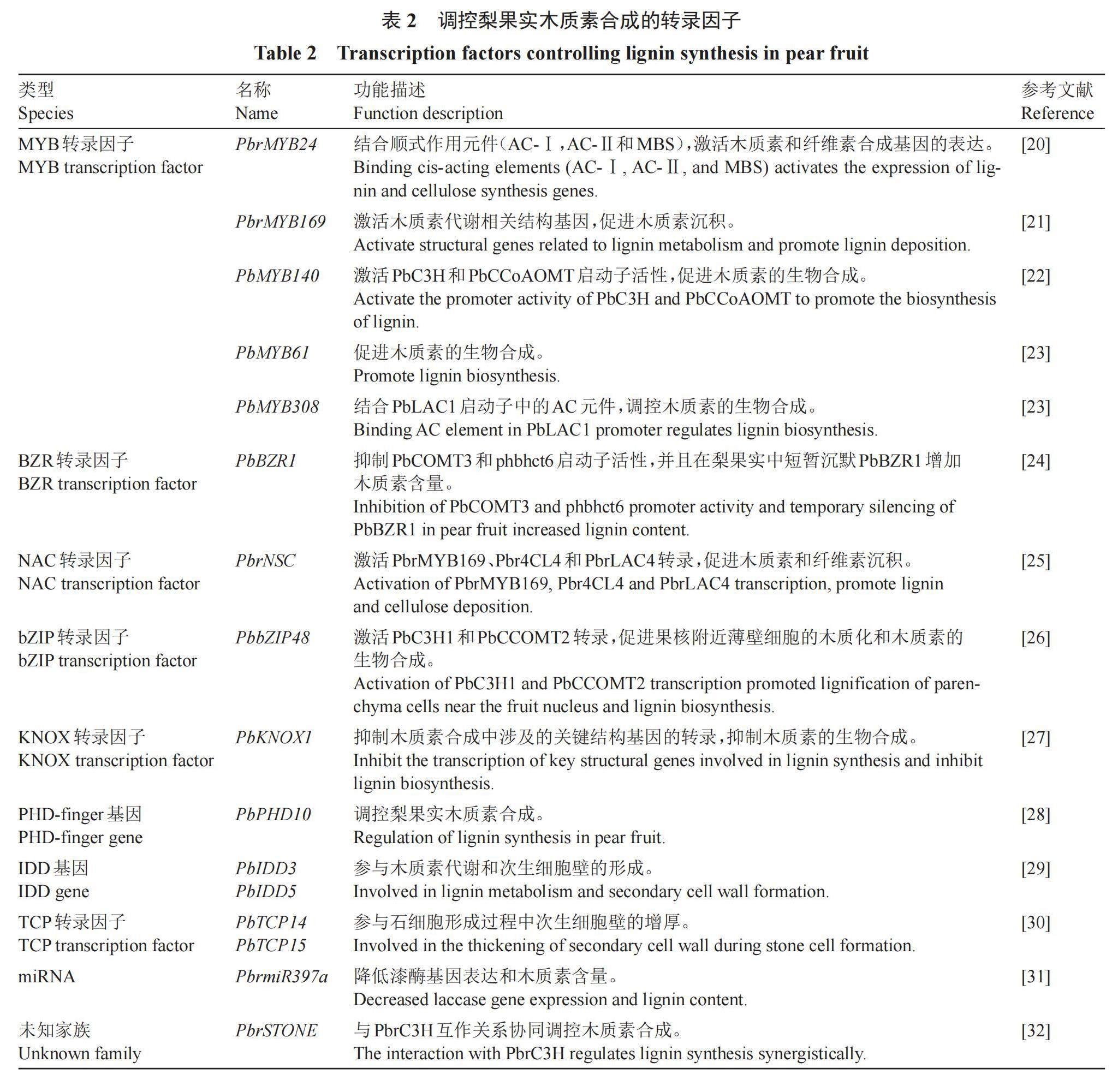
2 调控梨果实木质素合成的转录因子
在石细胞形成过程中,木质素生物合成基因形成了一个复杂的网络,转录因子的调节作用在其中发挥着重要作用,如MYB(v-myb avian myeloblastosis viral oncogene homolog)、NAC(NAM、ATAF和CUC)、bZIP(basic region-leucine zipper)、KNOX(Knotted1-like homebox)和鋅指蛋白(zinc-finger)等。其中MYB转录因子在木质素生物合成共表达网络中数量更多,并且与木质素通路结构基因的连接更显著(表2)[20-32]。
2.1 MYB
在梨中已鉴定出多个与木质素生物合成相关的MYB转录因子。通过qRT-PCR分析PbMYB的表达,鉴定出PbMYB25和PbMYB52参与梨果实发育过程中木质素生物合成的调控[33]。R2R3-MYB转录因子作为MYB转录因子中最大的一类,在梨基因组中已鉴定出104个,其中有28个被认为是参与木质素生物合成途径的调控因子[34]。近期研究发现,在梨果肉愈伤组织和拟南芥花序茎中过表达PbrMYB24,可显著提高木质素、纤维素含量,增厚次生细胞细胞壁。双荧光素酶试验、酵母单杂交试验和EMSA表明,PbrMYB24通过结合不同的顺式作用元件(AC-Ⅰ,AC-Ⅱ和MBS)直接激活木质素和纤维素合成基因的表达。此外,PbrMYB24可以调控PbrNSC和PbrMYB169的表达,同时也能被PbrNSC和PbrMYB169所激活,形成的层级调控网络协同调控石细胞中木质素和纤维素合成[20]。
PbrMYB169是木质素生物合成激活型转录因子,可通过选择性识别启动子中的AC-原件(ACCTAACC)结合,显著激活木质素代谢相关8个结构基因(C3H1、CCR1、CCOMT2、CAD、4CL1、4CL2、HCT2和LAC18)启动子活性,引起木质素沉积。在转基因拟南芥中,过表达PbrMYB169可有效增强木质素基因的表达,促进木质素沉积和增加细胞壁厚度,但未改变丁香基和愈创木酰基木质素单体的比例[21]。这与PbBZR1相反,PbBZR1抑制木质素生物合成基因PbCOMT3和phbhct6启动子的活性,并且在梨果实中短暂沉默PbBZR1增加木质素含量,诱导15个木质素生物合成基因的表达[24]。宋林艳[22]发现PbMYB140受上游调控因子PbMYB46和PbMYB83的激活调控,PbMYB83也可间接促进木质素生物合成。
参与次生细胞壁合成和木质素生物合成调节的MYB转录因子是高度保守的。通过瞬时过表达和基因沉默以及稳定转化苹果愈伤组织,发现PbMYB61可以促进木质素的生物合成,PbMYB308具有相反的作用。进一步的试验结果表明,PbMYB61通过与PbLAC1启动子中的AC元件结合上调表达来调节石细胞中木质素的形成。然而,PbMYB308通过与PbMYB61结合形成不能激活PbLAC1表达的二聚体来负调节石细胞木质素合成。由此阐明了PbMYB308-PbMYB61-PbLAC1模块参与梨石细胞中木质素的生物合成的分子机制[23]。
2.2 NAC、bZIP、KNOX
Wang等[25]通过整合木质素纤维素共表达网络和梨果实石细胞eQTL图谱,在206个梨品种中发现了与AtNST1/2/3相关的NAC转录因子,鉴定出一个NAC候选转录因子PbrNSC(NAC stone cell promoting factor)。研究证实PbrNSC可以通过激活PbrMYB169、Pbr4CL4和PbrLAC4启动子发挥调控功能,促进梨果实石细胞形成和木质素沉积。研究还发现PbrNSC蛋白C端存在两个保守的LP和WQ结构域,突变任何一个保守结构域都会影响PbrNSC转录因子对下游基因的转录调控作用,从而影响PbrNSC转录因子正向调控次生细胞壁合成的生物学功能。Gong等[26]采用转录组学、蛋白质组学和代谢组学等手段,构建了梨果实内石细胞与维管束的分布规律模型。通过共表达网络和基因表达模式分析,发现一个关键转录因子PbbZIP48,该转录因子基因在果核附近高度表达,通过激活PbC3H1和PbCCOMT2的转录,促进了果核附近薄壁细胞的木质化,导致石细胞的大量形成。同时研究显示,PbbZIP48通过与PbC3H1和PbCCOMT2启动子区域的G-box(ACGTG)基序结合促进两者表达量上调。KNOX基因家族中的BP(brevipedicellus,BP)在木质素代谢和细胞壁发育中起负调控作用。Cheng等[27]在梨基因组中鉴定出18个非重复的KNOX基因,主要分布在STM-like、BP-like、KNAT2/6-like、KNAT7-like和KNAT3-5-like亚家族中。系统发育树聚类和序列比对表明,PbKNOX1与拟南芥BP基因是梨同源基因。同时研究发现PbKNOX1作为木质素代谢的负调节因子,其过表达不仅使导管细胞次生细胞壁厚度降低了约19%,木质素含量降低约13%,还可以下调木质素代谢途径中多个关键结构基因(C4H、C3H、HCT、CCOMT、CCR、F5H、COMT和CAD)的表达,抑制石细胞木质化。
2.3 其他转录因子
植物同源结构域(plant homeodomain,PHD)是锌指结构域家族的一类转录调控因子,广泛参与植物的生长发育过程。Cao等[28]利用qRT-PCR技术鉴定了梨果实中10个表达的PHD-finger基因,其中PbPHD10被鉴定为调控木质素合成的重要候选基因。IDD(indeterminate domain)家族蛋白编码作为锌指蛋白转录因子之一,也参与梨果实木质素合成。如PbIDD3和PbIDD5被推断参与了梨果实木质素代谢和次生细胞壁的形成[29]。TCP(teosinte branched1,cycloidea,and proliferating cell factor family)蛋白是植物特有的转录因子,参与调节次生细胞壁的形成和程序性细胞死亡[35]。根据砀山酥梨不同发育阶段表达模式分析,表明PbTCP14和PbTCP15与果实木质素的积累模式和石细胞含量趋势一致,这两个转录因子可能参与了梨果实石细胞形成过程中次生细胞壁的增厚[30]。
除转录因子调控外,许多小分子RNA(microRNA,miRNA)能够参与调节次生细胞壁合成过程中的木质化。PbrmiR397a已被证明通过转录后调控PbrLAC抑制漆酶基因表达,从而调控梨果实木质素沉积,最终降低果实石细胞含量;研究还发现了PbrmiR397a启动子区域与石细胞含量相关的重要SNP位点,为开发分子检测标记和辅助育种奠定了基础[31]。Zhang等[32]通过全基因组关联研究,鉴定出一个与梨石细胞发育相关的新基因PbrSTONE,通过梨果实的瞬时转化和拟南芥的稳定转化验证,发现PbrSTONE可以调控梨果实石细胞和木质素的形成,并明确了其与木质素合成通路中关键基因PbrC3H存在互作关系,从而协同调控石细胞组分木质素的合成机制。
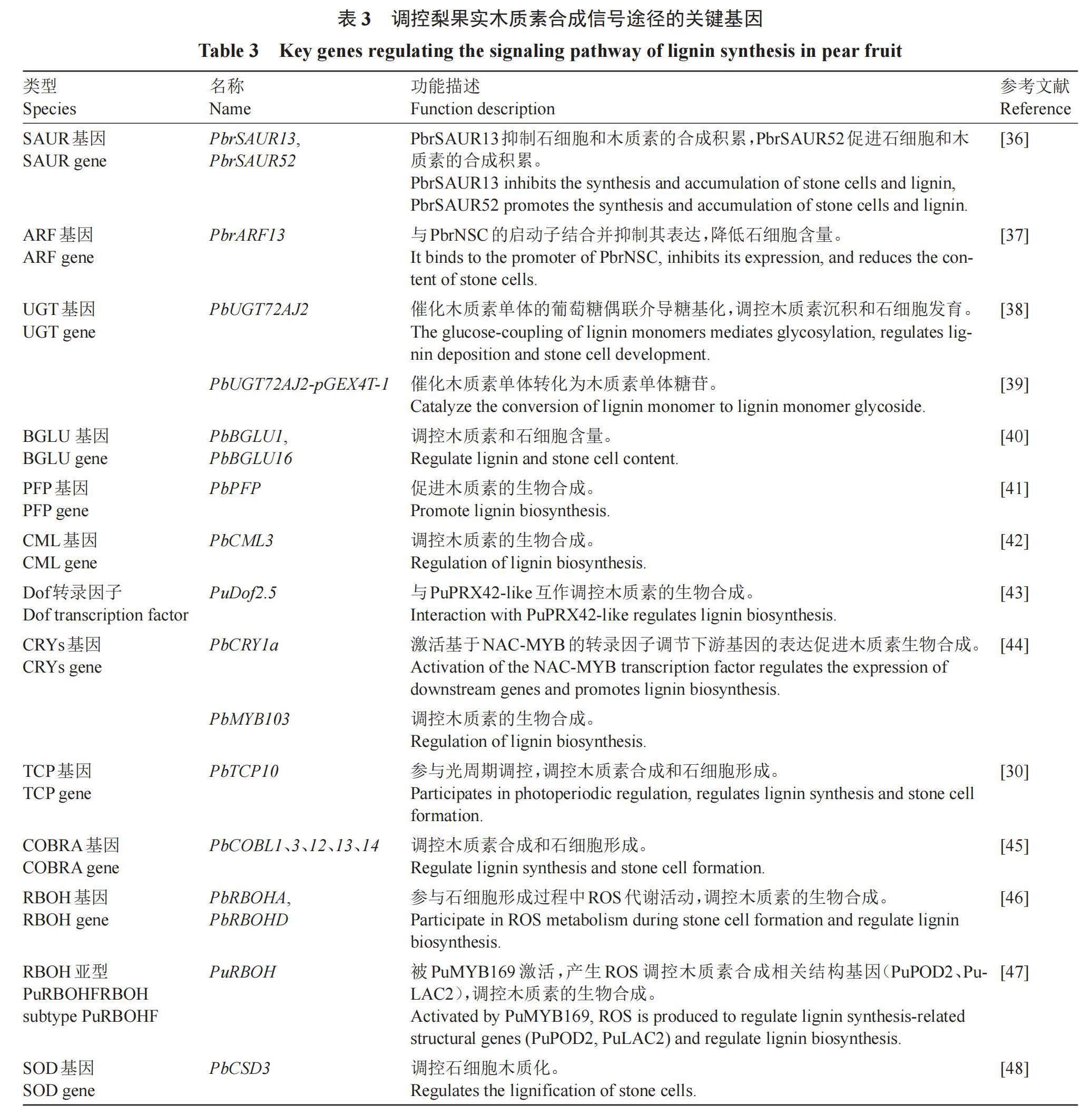
3 调控梨果实木质素合成的信号途径
梨果实木质素的合成调控是一个复杂的生物学过程,不仅直接受酶基因的影响,调节基因所编码的转录因子也可通过激活或抑制结构基因的表达,间接调控木质素的积累水平。同时与激素、糖、钙、光质等信号途径和环境因素密切相關(表3)[36-48]。
3.1 激素
3.1.1 生长素 生长素上调小RNA(small auxin-up RNA,SAUR)是生长素快速响应的基因,促进形成层活性,导致富含木质素的维管组织发育,进而调控木质素的积累[49]。砀山酥梨中共鉴定出116个SAUR基因,筛选出PbrSAUR13和PbrSAUR52两个关键基因。通过对39 d砀山酥梨果实的瞬时转化和草莓的稳定转化体系,发现PbrSAUR52主要促进石细胞和木质素的合成积累,而PbrSAUR13抑制石细胞和木质素的合成积累[36]。近期研究发现,外施200 μmol·L-1萘乙酸(NAA)可降低梨果实石细胞的含量,同时降低转录调节因子PbrNSC的表达量。PbrNSC能够直接结合生长素响应因子PbrARF13,过表达PbrARF13可显著降低梨果实中的石细胞含量;而利用病毒诱导的基因沉默(VIGS)抑制PbrARF13的表达,表型则相反。从而明确了PbrARF13可直接与PbrNSC的启动子结合并抑制其表达,降低石细胞含量。此外,通过石细胞形成相关基因的共表达网络分析,PbrNSC被确定为PbrMYB132的上游调控因子,PbrMYB132能够与纤维素合成酶基因(PbrCESA4b/7a/8a)和木质素合成基因(PbrLAC5)的启动子结合,激活其表达,促进纤维素和木质素的合成。由此,PbrARF13-PbrNSC-PbrMYB132调控级联能够响应生长素信号,调控梨果实石细胞中纤维素和木质素的生物合成[37]。
3.1.2 其他激素 脱落酸(abscisic acid,ABA)、茉莉酸甲酯(methyl jasmonate,MeJA)和水杨酸(salicylic acid,SA)等外源激素可通过影响木质素生物合成途径关键基因表达和转录因子,显著抑制梨果肉石细胞的木质化。Li等[7]在PbPALs家族成员的上游调控序列中发现了多个响应激素的元件,其中PbPAL1只含有ABA反应元件(ABA response element,ABRE),PbPAL3含有ABA响应元件ABRE、MeJA响应元件(MeJA response element,JARE)和SA响应元件(SA response,TCA),并且在PbPAL1和PbPAL3上游2000 bp启动子序列中发现了AC元件,故推测ABA、SA和MeJA可以通过调节PbPAL1和PbPAL3基因表达,进而影响梨果实石细胞的形成。
PbKNOX启动子含有TCA、CGTCA基序和ABRE等激素响应元件,SA通过诱导microRNA的表达可以调节梨果实木质素合成和石细胞发育[31]。由此推断,SA、ABA和MeJA可能直接或间接调节PbKNOXs的表达,从而影响石细胞的形成。大多数PbRBOH启动子含有大量与激素相关的元件,研究表明3种激素(SA、ABA和MeJA)处理梨果实后,大多数PbRBOHs在短时间内被转录诱导,对PbRBOHs的表达有显著影响。值得注意的是,10种PbRBOH启动子中并未发现SA应答元件。然而,在SA处理的果实中,PbRBOHI的表达水平仍然发生了变化。这可能是各种植物激素之间相互作用、相互诱导的结果[50]。
此外,研究发现ABA处理后梨果实中PbC4Hs的表达量先升高后降低,而MeJA处理后PbC4Hs的表达水平显著升高。经SA处理后,梨果实中PbC4H1和PbC4H2表达水平升高,PbC4H3表达水平下降[8]。因此,适当的生长素和细胞分裂素配比可以抑制木质素的合成。这些结果为梨果生产提供了重要的指导,可以通过喷施外源激素来调节梨果实木质素代谢的强度,从而不同程度地抑制石细胞的形成,改善果实品质。
3.2 糖
木质素的代谢过程主要包括木质素单体的生物合成、运输和聚合。木质素单体合成过程中通常会发生糖基化修饰,糖基化可以提升木质素单体的溶解度和稳定性,有利于木质素单体的运输和储存[51]。从木质素单体到木质素单体糖苷的转化需要尿苷二磷酸糖基转移酶(uridine diphosphate glycosyltransferase,UGT)的参与。PbUGT72AJ2基因参与木质素单体糖基化[38],重组蛋白PbUGT72AJ2-pGEX4T-1能够催化梨果实中木质素单体转化为木质素单体糖苷。Wang等[39]研究了PbUGT72A2转录在梨果实中的定位,并分析了PbUGT72AJ2的酶促反應动力。通过梨果实瞬时转化验证,在梨果实中过表达PbUGT72AJ2,木质素和石细胞含量几乎没有变化,而沉默该基因则显著增加木质素和石细胞含量。由此,PbUGT72AJ2通过催化木质素单体的葡萄糖偶联介导糖基化,影响下游基因的表达以及木质素单体的含量,从而影响梨果实木质素沉积和石细胞发育。
木质素单体糖苷被运送到细胞壁的特定部位,之后木质素开始去糖基化,在β-葡萄糖苷酶的作用下水解为木质素单木质素醇,最终形成木质素[51]。β-葡萄糖苷酶(β-glucosidases,BGLU)去糖基化功能在木质素单体转运中起着重要作用。Wang等[40]筛选到3个可能参与木质素合成的候选基因PbBGLU1、PbBGLU15和PbBGLU16,其中PbBGLU1和PbBGLU16转录主要位于梨果实木质素沉积区和石细胞区,过表达PbBGLU1和PbBGLU16后,梨果实中木质素和石细胞含量显著增加,而沉默PbBGLU1和PbBGLU16后,与对照组之间木质素和石细胞的含量没有显著差异。
葡萄糖作为木质素生物合成的重要碳源,其代谢与苯丙烷代谢途径密切相关。外源葡萄糖可以激活葡萄糖信号通路并调节其下游代谢活动,通过上调苯丙烷通路相关基因(PbPAL、PbHCT、PbCOMT、PbPRX)的表达诱导苯丙烷化合物的产生[52]。Jiao等[41]研究发现,砀山酥梨果实发育早期果糖-6-磷酸-1-磷酸转移酶(fructose-6-phosphate-1-phosphoric acid,PFP)基因的表达模式与木质素含量的变化趋势一致,通过验证,PbPFP正调控梨果实木质素生物合成。此外还观察到葡萄糖信号与激素信号之间的广泛串扰,特别是与ABA、GA和SA信号的串接。Pbr016851.1、Pbr002006.1和Pbr035515.1作为AtMYB52的同源基因,在葡萄糖处理的愈伤组织中表达上调,说明葡萄糖反应性转录因子的调控在葡萄糖介导木质素生物合成中发挥着重要作用。因此,研究葡萄糖反应转录因子如何感知葡萄糖信号并调控下游苯丙烷通路基因是非常有前景和意义的。
3.3 钙
钙离子(Ca2+)作为细胞内信号转导的第二信使,在调节细胞壁形成、木质素合成和活性氧代谢等方面发挥着重要的生理作用[53],这暗示木质素代谢和石细胞的形成与钙有关。
外源钙通过促进钙信号转导和上调转录因子调控木质素生物合成,从而增加梨果实中石细胞含量[54]。研究发现,0.1%的硝酸钙能够减少木质素含量,0.5%的硝酸钙处理后,梨愈伤组织中木质素含量和次生代谢物的积累显著增加,愈伤组织呈现褐色和暗沉。同时研究证明,大量差异表达基因(differentially expressed genes,DEGs)的次级代谢途径基因(CML、CAM、CDPK、CBL和CIPK)和转录因子可提供外源钙调控网络,引起代谢途径的改变,导致梨愈伤组织中木质素的积累。其中类钙调蛋白(calmodulin-like protein,CML)作为Ca2+的主要感受器,可通过调节下游靶蛋白的活性来调节多种细胞功能[55]。根据转录组数据结合qRT-PCR分析发现,PbrCML表达状况与钙处理后的愈伤组织变化趋势一致,过表达PbCML3可以促进梨果实中木质素代谢,显著增加其石细胞含量,验证发现PbCML3基因极可能作为转运蛋白参与调控某些转录因子从而间接调控木质素的生物合成[42],但CML对木质素含量的具体调控机制有待进一步研究。果面喷施5% CaCl2可以降低新高梨果实中PRX的活性,从而抑制果实木质素的合成[56]。外施CaCl2可显著抑制黄金梨中PAL、CAD和PRX的活性,且PpCAD1和PpCAD2基因的表达下调,影响石细胞形成[57]。
Dof(DNA Binding With One Zinc Finger)转录因子在木质素沉积中是一个正向调节因子[58]。研究发现,CaCl2处理南果梨果实后,其内源Ca2+水平升高,PuDof2.5表达量下降。PuDof2.5作为转录激活因子,与木质素生物合成基因PuPRX42-like启动子结合并抑制其转录;因此,PRX活性降低,木質素的生物合成受到抑制,石细胞含量和密度降低。而过表达PuDof2.5可上调PuPRX42-like表达水平,增强PRX酶活性,显著加速木质素积累和石细胞形成[43]。梨果实发育早期叶面喷施2%的CaCl2,不仅可以提高果实贮藏期间Ca/N和Ca/K的比值,而且可以降低梨果实的硬度和木质素含量,抑制“铁头病”的发生[59]。
3.4 活性氧
活性氧(reactive oxygen species,ROS)作为信号分子在植物细胞木质化过程中发挥着不可或缺的作用。ROS迸发主要由呼吸爆发氧化酶(respiratory burst oxidase homologue,RBOH,又称NADPH氧化酶)产生,参与梨果实石细胞的木质化过程[60]。前人研究表明,PbRBOHA、PbRBOHB和PbRBOHD在梨果实中转录本丰度较高,其中PbRBOHA和PbRBOHD的表达趋势与梨果实石细胞含量一致,且亚细胞定位显示PbRBOHA和PbRBOHD分布在质膜上,结合超氧化物含量的变化和时空表达分析,推定PbRBOHA和PbRBOHD参与了石细胞形成过程中ROS代谢活动[46]。梨果实中的木质素单体由PbUGT72AJ1、PbUGT72AJ2和PbUGT72AK1催化形成木质素糖苷,然后通过膜转运到细胞壁沉积,在PbDIR4的催化下形成木质素低聚物。最后,POD和LAC利用RBOH(PbRBOHA和PbRBOHD)和SOD产生的ROS和O2催化木质素聚合物链的伸长[16]。
随着研究的深入,人们发现RBOH亚型PuRBOHF在梨果实石细胞次生壁形成中起重要作用。Wang等[47]利用共聚焦显微镜和透射电镜观察发现ROS与细胞壁木质化之间的空间一致性。此外,石细胞木质化可被RBOH抑制剂二苯基氯化碘盐(diphenyleneiodonium chloride,DPI)所抑制,在生理水平上证明了RBOH介导的ROS对梨果木质素生物合成的关键作用。PuRBOHF过表达的梨果实和愈伤组织的木质素含量显著高于对照,而PuRBOHF沉默则抑制了木质素的积累。在此基础上,PuRBOHF可被PuMYB169激活,并通过产生ROS调控木质素合成相关结构基因(PuPOD2、PuLAC2)的转录。这是首次揭示PuRBOHF基因参与梨果实木质素代谢的分子机制。
Li等[48]研究了11种PbSODs在梨不同发育时期、不同品种和不同激素处理下的表达模式,发现PbCSD3的转录水平与木质素含量变化一致,其表达量在成熟期先升高后降低,结合时空表达结果表明,PbCSD3是梨果实石细胞木质化过程中活性氧代谢的候选基因。研究证明,石细胞形成是一个活性氧诱导的程序性细胞死亡(programmed cell death,PCD)的过程,石细胞分化时期、ROS的积累与PCD重叠[61]。高表达的细胞死亡相关基因和蛋白(AED3-like、MC1-like、ACD11-like)表达模式与ROS相关基因一致,其早期表达更丰富,后期下调[62]。
3.5 光质
光质对木质素沉积具有重要影响,作为木质素生物合成途径中的第一个限速酶,PAL受蓝光、紫外线和远红光调控[63]。蓝光对梨果实木质化具有正向调控作用,有研究表明其主要通过调控MYB、NAC等转录因子发挥作用。在蓝光处理下PbMYB103在不同梨果实发育时期、不同梨品种呈现高表达,PbMYB103可能是响应蓝光的转录因子[44]。
隐花色素(cryptochromes,CRYs)介导的蓝光信号已被证明参与了次生细胞壁增厚与光周期调控开花[64-65]。Wang等[44]研究发现蓝光增强了梨愈伤组织中4CL1、4CL2、COMT、CES、LAC7等木质素合成相关基因的表达,促进了木质素的积累。此外,在白梨中鉴定出4种蓝光受体CRY,分别为PbCRY1a、PbCRY1b、PbCRY2a和PbCRY2b,其中PbCRY1a、PbCRY1b与AtCRY1密切相关。同时证实了PbCRY1a作为木质素沉积的上游信号,通过激活基于NAC-MYB的转录因子调节下游基因的表达促进石细胞木质化。由此,CRY介导的蓝光信号在梨果实细胞壁木质化过程中起重要作用,并通过调控下游基因促进梨果实石细胞的形成。
迄今为止,关于光调控梨果实中木质素生物合成作用的研究主要集中在生理方面,如不同颜色果袋对果实品质的影响。研究表明光质可以通过影响PAL、C4H、4CL等光诱导酶调控木质素的生物合成,绿色透光果袋抑制PbNAC83a的表达,促进PbNAC83f和PbNAC91a的表达。橙色透光果袋促进PbNAC83a、PbNAC83f和PbNAC91a的表达。PbNAC83a表达量受光质影响与木质素含量的变化趋势一致,其基因表达可能是受红光促进、蓝光抑制[66]。近期研究发现,TCP基因和COBRA基因参与了次生细胞壁的形成,其中PbTCP10,PbCOBL1、3、12、13、14在果实中高表达,其启动子的顺式作用元件含有光响应元件,参与了光周期调控,进而调节石细胞的形成[30,45]。此外,光响应元件如Box4、GATA-motif、GT1-motif也存在于RBOH启动子中,说明套袋处理改变梨果实木质素和石细胞含量可能与RBOH的转录有关[50]。Wang等[67]研究发现无纺布套袋梨果实,通过下调苯丙烷生物合成相关DEGs抑制茌梨梨果皮木质素合成,进而减小果实皮孔直径,并推断果皮木质素含量可能与套袋透光性、光合速率及波长有关,而造成这些影响的具体机制需要进一步探索。
4 花粉直感
不同品种授粉后,梨树花粉能直接影响当年内母本果实品质,果实硬度、可溶性固形物含量、可滴定酸含量和石细胞数量等与木质素含量相关的性状,也表现出明显的花粉直感现象[68]。Yan等[69]以鸭梨和圆黄对砀山酥梨授粉,果实石细胞含量表现出显著的花粉直感现象,以鸭梨授粉的果实石细胞团的数量和聚合度较高,细胞壁的致密性更强,但兩者授粉的梨果实石细胞团形成过程是一致的,说明梨果实的父本花粉直感现象表现具有阶段性。此外,一些初级代谢物和木质素合成中间代谢物也参与了花粉调控木质素代谢和石细胞形成,使用OPIS-DA模型分析授粉后47 d和64 d的砀山酥梨果实的初级代谢物和木质素合成中间代谢物,发现授粉通过影响糖、氨基酸、脂肪酸和香豆酸含量调控梨果实木质素合成[70]。由此可推断,花粉影响了梨果实中苯丙氨酸的合成,进而调控了木质素的合成。不同授粉品种对梨果实发育过程中果肉、种子内源激素含量及相关酶活性都具有重要影响[71]。
microRNA通过糖酸代谢和激素信号传导等途径广泛参与梨果实发育和果实品质的调控,其中microRNA受花粉直感现象的影响,参与调控梨果实木质素代谢和石细胞形成[72]。已有研究报道,不同授粉品种显著影响梨果实木质素代谢相关基因和蛋白质的表达,例如授粉影响果实漆酶基因microRNA的表达[73],过氧化物酶47(PER47)、β-葡萄糖苷酶(BGLU15)以及漆酶4(LAC4)的表达[74],间接调控木质素沉积,最终影响梨果实石细胞的形成,但花粉直感以何种方式调节microRNA表达进而影响果实石细胞形成需进一步探索。
5 总结与展望
梨果实木质素生物合成的调控过程复杂,涉及多层面、多因子的协同作用。目前,MYB、bZIP、NAC及KNOX类转录因子都参与了梨果实木质素生物合成的调控,Trihelix[75]、LIM[76]和LTF1[77]类转录因子同样在木质素合成调控中起到了关键作用。单独研究某个或少数几个转录因子不足以揭示木质素生物合成的巨大调控网络,基于NAC-MYB的基因调控网络(NAC-MYB-GRN)模型被广泛认为是支撑木质素生物合成的基础[78],并且已被证明参与梨石细胞的木质化过程。另外,研究发现,PbMYB308-PbMYB61-PbLAC1模块参与梨石细胞中木质素的生物合成[23];PbrARF13-PbrNSC-PbrMYB132级联能够响应生长素信号,调控梨果实石细胞中的纤维素和木质素生物合成[37],其他复合调控家族参与木质素合成的报道较少。
当前对梨果实木质素合成的单一信号途径的调控研究较多,多信号调控的交叉调控网络的研究较少。激素、糖、钙和活性氧等是影响木质素合成的重要信号调控因子。此外,可能存在其他信号因子参与调控梨果实木质素的生物合成,譬如短肽信号[79]、受体激酶[80]、泛素化和其他响应对木质素的沉积等。这些信号因子是否存在时空特异性与相互影响,对梨果实木质素生物合成的调控作用及分子机制,仍有待进一步阐明。据报道,区域生态、树形等外部因素及梨树的脱萼生物学功能与木质素沉积也具有相关性[81]。随着DNA/RNA甲基化测序、MicroRNA测序和CHIP-Seq等前沿技术的引入,以及多组学联合应用和CRISPR-Cas9技术的不断优化[82],得以构建完整的木质素合成调控网络来阐明调控梨果实中木质素生物合成的分子机制,实现梨果实木质素合成和石细胞形成的精准调控,提高梨果实品质。
参考文献 References:
[1] CAO Y P,HAN Y H,LI D H,LIN Y,CAI Y P. Systematic analysis of the 4-coumarate:Coenzyme a ligase (4CL) related genes and expression profiling during fruit development in the Chinese pear[J]. Genes,2016,7(10):89.
[2] 徐钰清,田路明,曹玉芬,董星光,张莹,霍宏亮,齐丹,徐家玉,刘超. 梨果肉质地及香气研究进展[J]. 果树学报,2023,40(4):757-770.
XU Yuqing,TIAN Luming,CAO Yufen,DONG Xingguang,ZHANG Ying,HUO Hongliang,QI Dan,XU Jiayu,LIU Chao. Research progress in flesh texture and aroma of pears[J]. Journal of Fruit Science,2023,40(4):757-770.
[3] 王红宝,朱洁,王丹阳,陶书田. 梨果肉石细胞含量分析[J]. 江苏农业科学,2018,46(3):173-176.
WANG Hongbao,ZHU Jie,WANG Danyang,TAO Shutian. Analysis of flesh stone cell content in pear fruit[J]. Jiangsu Agricultural Sciences,2018,46(3):173-176.
[4] JIN Q,YAN C C,QIU J X,ZHANG N,LIN Y,CAI Y P. Structural characterization and deposition of stone cell lignin in Dangshan Su pear[J]. Scientia Horticulturae,2013,155:123-130.
[5] LI M T,CHENG C X,ZHANG X F,ZHOU S P,LI L X,YANG S L. Overexpression of pear (Pyrus pyrifolia) CAD2 in tomato affects lignin content[J]. Molecules,2019,24(14):2595.
[6] ZHANG J Y,LI J M,XUE C,WANG R Z,ZHANG M Y,QI K J,FAN J,HU H J,ZHANG S L,WU J. The variation of stone cell content in 236 germplasms of sand pear (Pyrus pyrifolia) and identification of related candidate genes[J]. Horticultural Plant Journal,2021,7(2):108-116.
[7] LI G H,WANG H,CHENG X,SU X Q,ZHAO Y,JIANG T S,JIN Q,LIN Y,CAI Y P. Comparative genomic analysis of the PAL genes in five Rosaceae species and functional identification of Chinese white pear[J]. PeerJ,2019,7:e8064.
[8] LI G H,LIU X,ZHANG Y,MUHAMMAD A,HAN W L,LI D H,CHENG X,CAI Y P. Cloning and functional characterization of two cinnamate 4-hydroxylase genes from Pyrus bretschneideri[J]. Plant Physiology and Biochemistry,2020,156:135-145.
[9] MA C,ZHANG H P,LI J M,TAO S T,QIAO X,KORBAN S S,ZHANG S L,WU J. Genome-wide analysis and characterization of molecular evolution of the HCT gene family in pear (Pyrus bretschneideri)[J]. Plant Systematics and Evolution,2017,303(1):71-90.
[10] XU J H,TAO X Y,XIE Z H,GONG X,QI K J,ZHANG S L,SHIRATAKE K,TAO S T. PbCSE1 promotes lignification during stone cell development in pear (Pyrus bretschneideri) fruit[J]. Scientific Reports,2021,11:9450.
[11] 張盼盼. “二十世纪” 梨果锈形成过程中木质素合成及调控基因的挖掘与表达分析[D]. 扬州:扬州大学,2018.
ZHANG Panpan. Mining and expression analysis of regulatory genes and lignin biosynthesis during pear fruit rust formation in “Nijisseiki”[D]. Yangzhou:Yangzhou University,2018.
[12] CHENG X,XIONG Y,LI D H,CHENG J,CAO Y P,YAN C C,JIN Q,SUN N,CAI Y P,LIN Y. Bioinformatic and expression analysis of the OMT gene family in Pyrus bretschneideri cv. Dangshan Su[J]. Genetics and Molecular Research,2016,15(3):gmr. 15038664.
[13] CHENG X,LI M L,LI D H,ZHANG J Y,JIN Q,SHENG L L,CAI Y P,LIN Y. Characterization and analysis of CCR and CAD gene families at the whole-genome level for lignin synthesis of stone cells in pear (Pyrus bretschneideri) fruit[J]. Biology Open,2017,6(11):1602-1613.
[14] SU X Q,ZHAO Y,WANG H,LI G H,CHENG X,JIN Q,CAI Y P. Transcriptomic analysis of early fruit development in Chinese white pear (Pyrus bretschneideri Rehd.) and functional identification of PbCCR1 in lignin biosynthesis[J]. BMC Plant Biology,2019,19(1):417.
[15] QI K J,SONG X F,YUAN Y Z,BAO J P,GONG X,HUANG X S,KHANIZADEH S,ZHANG S L,TAO S T. CAD genes:Genome-wide identification,evolution,and their contribution to lignin biosynthesis in pear (Pyrus bretschneideri)[J]. Plants,2021,10(7):1444.
[16] CHENG X,LI G H,MA C H,ABDULLAH M,ZHANG J Y,ZHAO H,JIN Q,CAI Y P,LIN Y. Comprehensive genome-wide analysis of the pear (Pyrus bretschneideri) laccase gene (PbLAC) family and functional identification of PbLAC1 involved in lignin biosynthesis[J]. PLoS One,2019,14(2):e0210892.
[17] YANG S L,ZHANG X N,LU G L,WANG C R,WANG R. Regulation of gibberellin on gene expressions related with the lignin biosynthesis in ‘Wangkumbae pear (Pyrus pyrifolia Nakai) fruit[J]. Plant Growth Regulation,2015,76(2):127-134.
[18] MA R F,LIU Q Z,XIAO Y,ZHANG L,LI Q,YIN J,CHEN W S. The phenylalanine ammonia-lyase gene family in Isatis indigotica Fort.:Molecular cloning,characterization,and expression analysis[J]. Chinese Journal of Natural Medicines,2016,14(11):801-812.
[19] CHENG X,ZHANG J Y,WANG H,CHEN T Z,LI G H,YAN C C,JIN Q,LIN Y,CAI Y P. Effects of metaxenia on stone cell formation in pear (Pyrus bretschneideri) based on transcriptomic analysis and functional characterization of the lignin-related gene PbC4H2[J]. Forests,2020,11(1):53.
[20] XUE Y S,SHAN Y F,YAO J L,WANG R Z,XU S Z,LIU D L,YE Z C,LIN J,LI X G,XUE C,WU J. The transcription factor PbrMYB24 regulates lignin and cellulose biosynthesis in stone cells of pear fruits[J]. Plant Physiology,2023,192(3):1997-2014.
[21] XUE C,YAO J L,XUE Y S,SU G Q,WANG L,LIN L K,ALLAN A C,ZHANG S L,WU J. PbrMYB169 positively regulates lignification of stone cells in pear fruit[J]. Journal of Experimental Botany,2019,70(6):1801-1814.
[22] 宋林艷. 梨花青苷和木质素合成调控相关SG4-R2R3-MYB基因的筛选及其功能解析[D]. 杨凌:西北农林科技大学,2021.
SONG Linyan. Identification and characterization of the SG4-R2R3-MYB genes that related with anthocyanin and lignin biosynthesis and regulation in pear[D]. Yangling:Northwest A & F University,2021.
[23] ZHU Y S,WANG Y C,JIANG H Y,LIU W J,ZHANG S H,HOU X K,ZHANG S S,WANG N,ZHANG R,ZHANG Z Y,CHEN X S. Transcriptome analysis reveals that PbMYB61 and PbMYB308 are involved in the regulation of lignin biosynthesis in pear fruit stone cells[J]. The Plant Journal,2023,116(1):217-233.
[24] CAO Y P,MENG D D,LI X X,WANG L H,CAI Y P,JIANG L. A Chinese white pear (Pyrus bretschneideri) BZR gene PbBZR1 act as a transcriptional repressor of lignin biosynthetic genes in fruits[J]. Frontiers in Plant Science,2020,11:1087.
[25] WANG R Z,XUE Y S,FAN J,YAO J L,QIN M F,LIN T,LIAN Q,ZHANG M Y,LI X L,LI J M,SUN M Y,SONG B B,ZHANG J Y,ZHAO K J,CHEN X,HU H J,FEI Z J,XUE C,WU J. A systems genetics approach reveals PbrNSC as a regulator of lignin and cellulose biosynthesis in stone cells of pear fruit[J]. Genome Biology,2021,22(1):313.
[26] GONG X,QI K J,CHEN J L,ZHAO L Y,XIE Z H,YAN X,KHANIZADEH S,ZHANG S L,TAO S T. Multi-omics analyses reveal stone cell distribution pattern in pear fruit[J]. The Plant Journal,2023,113(3):626-642.
[27] CHENG X,LI M L,ABDULLAH M,LI G H,ZHANG J Y,MANZOOR M A,WANG H,JIN Q,JIANG T S,CAI Y P,LI D H,LIN Y. In silico genome-wide analysis of the pear (Pyrus bretschneideri) KNOX family and the functional characterization of PbKNOX1,an Arabidopsis BREVIPEDICELLUS orthologue gene,involved in cell wall and lignin biosynthesis[J]. Frontiers in Genetics,2019,10:632.
[28] CAO Y P,HAN Y H,MENG D D,ABDULLAH M,LI D H,JIN Q,LIN Y,CAI Y P. Systematic analysis and comparison of the PHD-Finger gene family in Chinese pear (Pyrus bretschneideri) and its role in fruit development[J]. Functional & Integrative Genomics,2018,18(5):519-531.
[29] SU X Q,MENG T K,ZHAO Y,LI G H,CHENG X,ABDULLAH M,SUN X,CAI Y P,LIN Y. Comparative genomic analysis of the IDD genes in five Rosaceae species and expression analysis in Chinese white pear (Pyrus bretschneideri)[J]. PeerJ,2019,7:e6628.
[30] ZHAO Y,SU X Q,WANG X Y,WANG M N,CHI X J,MANZOOR M A,LI G H,CAI Y P. Comparative genomic analysis of TCP genes in six rosaceae species and expression pattern analysis in Pyrus bretschneideri[J]. Frontiers in Genetics,2021,12:669959.
[31] XUE C,YAO J L,QIN M F,ZHANG M Y,ALLAN A C,WANG D F,WU J. PbrmiR397a regulates lignification during stone cell development in pear fruit[J]. Plant Biotechnology Journal,2019,17(1):103-117.
[32] ZHANG M Y,XUE C,HU H J,LI J M,XUE Y S,WANG R Z,FAN J,ZOU C,TAO S T,QIN M F,BAI B,LI X L,GU C,WU S,CHEN X,YANG G Y,LIU Y Y,SUN M Y,FEI Z J,ZHANG S L,WU J. Genome-wide association studies provide insights into the genetic determination of fruit traits of pear[J]. Nature Communications,2021,12:1144.
[33] CAO Y P,HAN Y H,LI D H,LIN Y,CAI Y P. MYB transcription factors in Chinese pear (Pyrus bretschneideri Rehd.):Genome-wide identification,classification,and expression profiling during fruit development[J]. Frontiers in Plant Science,2016,7:577.
[34] LI X L,XUE C,LI J M,QIAO X,LI L T,YU L A,HUANG Y H,WU J. Genome-wide identification,evolution and functional divergence of MYB transcription factors in Chinese white pear (Pyrus bretschneideri)[J]. Plant and Cell Physiology,2016,57(4):824-847.
[35] CAO J F,ZHAO B,HUANG C C,CHEN Z W,ZHAO T,LIU H R,HU G J,SHANGGUAN X X,SHAN C M,WANG L J,ZHANG T Z,WENDEL J F,GUAN X Y,CHEN X Y. The miR319-targeted GhTCP4 promotes the transition from cell elongation to wall thickening in cotton fiber[J]. Molecular Plant,2020,13(7):1063-1077.
[36] WANG M N,MANZOOR M A,WANG X Y,FENG X F,ZHAO Y,HE J L,CAI Y P. Comparative genomic analysis of SAUR gene family,cloning and functional characterization of two genes (PbrSAUR13 and PbrSAUR52) in Pyrus bretschneideri[J]. International Journal of Molecular Sciences,2022,23(13):7054.
[37] XU S Z,SUN M Y,YAO J L,LIU X X,XUE Y S,YANG G Y,ZHU R X,JIANG W T,WANG R Z,XUE C,MAO Z Q,WU J. Auxin inhibits lignin and cellulose biosynthesis in stone cells of pear fruit via the PbrARF13-PbrNSC-PbrMYB132 transcriptional regulatory cascade[J]. Plant Biotechnology Journal,2023,21(7):1408-1425.
[38] CHENG X,MUHAMMAD A,LI G H,ZHANG J Y,CHENG J,QIU J X,JIANG T S,JIN Q,CAI Y P,LIN Y. Family-1 UDP glycosyltransferases in pear (Pyrus bretschneideri):Molecular identification,phylogenomic characterization and expression profiling during stone cell formation[J]. Molecular Biology Reports,2019,46(2):2153-2175.
[39] WANG H,FENG X F,ZHANG Y J,WEI D Y,ZHANG Y,JIN Q,CAI Y P. PbUGT72AJ2-mediated glycosylation plays an important role in lignin formation and stone cell development in pears (Pyrus bretschneideri)[J]. International Journal of Molecular Sciences,2022,23(14):7893.
[40] WANG H,ZHANG Y J,FENG X F,PENG F L,MAZOOR M A,ZHANG Y,ZHAO Y,HAN W L,LU J J,CAO Y P,CAI Y P. Analysis of the β-glucosidase family reveals genes involved in the lignification of stone cells in Chinese white pear (Pyrus bretschneideri Rehd.)[J]. Frontiers in Plant Science,2022,13:852001.
[41] JIAO Y R,GONG X,QI K J,XIE Z H,WANG Y L,YUAN K L,PAN Q,ZHANG S L,SHIRATAKE K,KHANIZADEH S,TAO S T. Transcriptome analysis provides new ideas for studying the regulation of glucose-induced lignin biosynthesis in pear calli[J]. BMC Plant Biology,2022,22(1):310.
[42] TAO X Y,LIU M,YUAN Y Z,LIU R N,QI K J,XIE Z H,BAO J P,ZHANG S L,SHIRATAKE K,TAO S T. Transcriptome provides potential insights into how calcium affects the formation of stone cell in Pyrus[J]. BMC Genomics,2021,22(1):831.
[43] ZHANG H,GAO S Y,WANG T Y,XU M Y,LI X Y,DU G D. Ca2+ mediates transcription factor PuDof2.5 and suppresses stone cell production in pear fruits[J]. Frontiers in Plant Science,2022,13:976977.
[44] WANG Q,GONG X,XIE Z H,QI K J,YUAN K L,JIAO Y R,PAN Q,ZHANG S L,SHIRATAKE K,TAO S T. Cryptochrome-mediated blue-light signal contributes to lignin biosynthesis in stone cells in pear fruit[J]. Plant Science,2022,318:111211.
[45] ZHAO Y,SU X Q,WANG X Y,WANG M N,FENG X F,AAMIR MANZOOR M,CAI Y P. Comparative genomic analysis of the COBRA genes in six Rosaceae species and expression analysis in Chinese white pear (Pyrus bretschneideri)[J]. PeerJ,2022,10:e13723.
[46] CHENG X,LI G H,MANZOOR M A,WANG H,ABDULLAH M,SU X Q,ZHANG J Y,JIANG T S,JIN Q,CAI Y P,LIN Y. In silico genome-wide analysis of respiratory burst oxidase homolog (RBOH) family genes in five fruit-producing trees,and potential functional analysis on lignification of stone cells in Chinese white pear[J]. Cells,2019,8(6):520.
[47] WANG X Q,LIU S Q,SUN H L,LIU C Y,LI X Y,LIU Y,LYU D G,DU G D. Production of reactive oxygen species by PuRBOHF is critical for stone cell development in pear fruit[J]. Horticulture Research,2021,8:249.
[48] LI G H,HU F,ZHANG Y,ZHAO Y,WANG H,CHEN T Z,CHENG X,CAI Y P. Comparative genomic analysis of superoxide dismutase (SOD) genes in three Rosaceae species and expression analysis in Pyrus bretschneideri[J]. Physiology and Molecular Biology of Plants,2021,27(1):39-52.
[49] ZHANG L M,KAMITAKAHARA H,SASAKI R,OIKAWA A,SAITO K,MURAYAMA H,OHSAKO T,ITAI A. Effect of exogenous GA4 + 7 and BA + CPPU treatments on fruit lignin and primary metabolites in Japanese pear “Gold Nijisseiki”[J]. Scientia Horticulturae,2020,272:109593.
[50] TAO S T,WANG D Y,JIN C,SUN W,LIU X,ZHANG S L,GAO F Y,KHANIZADEH S. Cinnamate-4-hydroxylase gene is involved in the step of lignin biosynthesis in Chinese white pear[J]. Journal of the American Society for Horticultural Science,2015,140(6):573-579.
[51] LIU C J. Deciphering the enigma of lignification:Precursor transport,oxidation,and the topochemistry of lignin assembly[J]. Molecular Plant,2012,5(2):304-317.
[52] LIU J,HUANG Q H,KANG P Z,LIANG L,CHEN J J. Lignin accumulation in three pumelo cultivars in association with sucrose and energy depletion[J]. Biomolecules,2019,9(11):701.
[53] MICHAILIDIS M,KARAGIANNIS E,TANOU G,KARAMANOLI K,LAZARIDOU A,MATSI T,MOLASSIOTIS A. Metabolomic and physico-chemical approach unravel dynamic regulation of calcium in sweet cherry fruit physiology[J]. Plant Physiology and Biochemistry,2017,116:68-79.
[54] ZHAO D Q,TANG Y H,XIA X,SUN J,MENG J S,SHANG J L,TAO J. Integration of transcriptome,proteome,and metabolome provides insights into how calcium enhances the mechanical strength of herbaceous peony inflorescence stems[J]. Cells,2019,8(2):102.
[55] LIN W D,LIAO Y Y,YANG T J W,PAN C Y,BUCKHOUT T J,SCHMIDT W. Coexpression-based clustering of Arabidopsis root genes predicts functional modules in early phosphate deficiency signaling[J]. Plant Physiology,2011,155(3):1383-1402.
[56] LEE S H,CHOI J H,KIM W S,PARK Y S,GEMMA H. Effects of calcium chloride spray on peroxidase activity and stone cell development in pear fruit (Pyrus pyrifolia ‘Niitaka)[J]. Journal of the Japanese Society for Horticultural Science,2007,76(3):191-196.
[57] LU G L,LI Z J,ZHANG X F,WANG R,YANG S L. Expression analysis of lignin-associated genes in hard end pear (Pyrus pyrifolia Whangkeumbae) and its response to calcium chloride treatment conditions[J]. Journal of Plant Growth Regulation,2015,34(2):251-262.
[58] RAMACHANDRAN V,TOBIMATSU Y,MASAOMI Y,SANO R,UMEZAWA T,DEMURA T,OHTANI M. Plant-specific Dof transcription factors VASCULAR-RELATED DOF1 and VASCULAR-RELATED DOF2 regulate vascular cell differentiation and lignin biosynthesis in Arabidopsis[J]. Plant Molecular Biology,2020,104(3):263-281.
[59] WANG Y L,ZHANG X F,WANG Y Z,YANG S L,QU H Y. The changes of intracellular calcium concentration and distribution in the hard end pear (Pyrus pyrifolia cv. ‘Whangkeumbae) fruit[J]. Cell Calcium,2018,71:15-23.
[60] BERTHET S,DEMONT-CAULET N,POLLET B,BIDZINSKI P,C?ZARD L,LE BRIS P,BORREGA N,HERV? J,BLONDET E,BALZERGUE S,LAPIERRE C,JOUANIN L. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification of Arabidopsis thaliana stems[J]. The Plant Cell,2011,23(3):1124-1137.
[61] 艾沙江·买买提,张校立,梅闯,马凯,闫鹏,韩立群,王继勋. ‘库尔勒香梨果实发育过程中石细胞形成与细胞凋亡的关系研究[J]. 果树学报,2020,37(1):59-67.
Aisajiang·Mamat,ZHANG Xiaoli,MEI Chuang,MA Kai,YAN Peng,HAN Liqun,WANG Jixun. Study on the relationship between the stone cell formation and apoptosis during the fruit development of ‘Kuerlexiangli pear[J]. Journal of Fruit Science,2020,37(1):59-67.
[62] MAMAT A,TUSONG K,XU J,YAN P,MEI C,WANG J X. Integrated transcriptomic and proteomic analysis reveals the complex molecular mechanisms underlying stone cell formation in Korla pear[J]. Scientific Reports,2021,11(1):7688.
[63] SRIVASTAVA S,VISHWAKARMA R K,ARAFAT Y A,GUPTA S K,KHAN B M. Abiotic stress induces change in Cinnamoyl CoA Reductase (CCR) protein abundance and lignin deposition in developing seedlings of Leucaena leucocephala[J]. Physiology and Molecular Biology of Plants,2015,21(2):197-205.
[64] ZHANG Q,XIE Z,ZHANG R,XU P,LIU H T,YANG H Q,DOBLIN M S,BACIC A,LI L G. Blue light regulates secondary cell wall thickening via MYC2/MYC4 activation of the NST1-directed transcriptional network in Arabidopsis[J]. The Plant Cell,2018,30(10):2512-2528.
[65] LIU H T,LIU B,ZHAO C X,PEPPER M,LIN C T. The action mechanisms of plant cryptochromes[J]. Trends in Plant Science,2011,16(12):684-691.
[66] 張齐彦. PbNAC83a、PbNAC83f和PbNAC91a调控梨果实木质素合成机制研究[D]. 南京:南京农业大学,2019.
ZHANG Qiyan. The regulating mechanism of PbNAC83a,PbNAC83f and PbNAC91a about lignin synthesis on pear fruits[D]. Nanjing:Nanjing Agricultural University,2019.
[67] WANG Y L,ZHANG X F,WANG R,BAI Y X,LIU C L,YUAN Y B,YANG Y J,YANG S L. Differential gene expression analysis of ‘Chili (Pyrus bretschneideri) fruit pericarp with two types of bagging treatments[J]. Horticulture Research,2017,4:17005.
[68] ROSIANSKI Y,FREIMAN Z E,COCHAVI S M,YABLOVITZ Z,KEREM Z,FLAISHMAN M A. Advanced analysis of developmental and ripening characteristics of pollinated common-type fig (Ficus carica L.)[J]. Scientia Horticulturae,2016,198:98-106.
[69] YAN C C,ZHANG N,XU C,JIN Q,QI Y J,CAI Y P. Effects on stone cell development and lignin deposition in pears by different pollinators[J]. Frontiers in Plant Science,2023,14:1093661.
[70] LI S M,SU X Q,ABDULLAH M,SUN Y M,LI G H,CHENG X,LIN Y,CAI Y P,JIN Q. Effects of different pollens on primary metabolism and lignin biosynthesis in pear[J]. International Journal of Molecular Sciences,2018,19(8):2273.
[71] LI X T,BAO J P. Effects of different pollination combinations on stone cells,lignin,and related enzyme activities in fragrant pear fruit[J]. HortScience,2022,57(5):652-656.
[72] WU J,WANG D F,LIU Y F,WANG L,QIAO X,ZHANG S L. Identification of miRNAs involved in pear fruit development and quality[J]. BMC Genomics,2014,15(1):953.
[73] CHENG X,YAN C C,ZHANG J Y,MA C H,LI S M,JIN Q,ZHANG N,CAO Y P,LIN Y,CAI Y P. The effect of different pollination on the expression of Dangshan Su pear microRNA[J]. BioMed Research International,2017,2017:2794040.
[74] LI S M,SU X Q,JIN Q,LI G H,SUN Y M,ABDULLAH M,CAI Y P,LIN Y. iTRAQ-based identification of proteins related to lignin synthesis in the pear pollinated with pollen from different varieties[J]. Molecules,2018,23(3):548.
[75] GAO H Y,HUANG R,LIU J,GAO Z M,ZHAO H S,LI X P. Genome-wide identification of Trihelix genes in moso bamboo (Phyllostachys edulis) and their expression in response to abiotic stress[J]. Journal of Plant Growth Regulation,2019,38(3):1127-1140.
[76] CHENG X,LI G H,MUHAMMAD A,ZHANG J Y,JIANG T S,JIN Q,ZHAO H,CAI Y P,LIN Y. Molecular identification,phylogenomic characterization and expression patterns analysis of the LIM (LIN-11,Isl1 and MEC-3 domains) gene family in pear (Pyrus bretschneideri) reveal its potential role in lignin metabolism[J]. Gene,2019,686:237-249.
[77] GUI J S,LUO L F,ZHONG Y,SUN J Y,UMEZAWA T,LI L G. Phosphorylation of LTF1,an MYB transcription factor in Populus,acts as a sensory switch regulating lignin biosynthesis in wood cells[J]. Molecular Plant,2019,12(10):1325-1337.
[78] OHTANI M,DEMURA T. The quest for transcriptional hubs of lignin biosynthesis:Beyond the NAC-MYB-gene regulatory network model[J]. Current Opinion in Biotechnology,2019,56:82-87.
[79] JOHNSSON C,JIN X,XUE W Y,DUBREUIL C,LEZHNEVA L,FISCHER U. The plant hormone auxin directs timing of xylem development by inhibition of secondary cell wall deposition through repression of secondary wall NAC-domain transcription factors[J]. Physiologia Plantarum,2019,165(4):673-689.
[80] LIU C,YU H S,VOXEUR A,RAO X L,DIXON R A. FERONIA and wall-associated kinases coordinate defense induced by lignin modification in plant cell walls[J]. Science Advances,2023,9(10):eadf7714.
[81] 汪曉谦,商叶,刘维成,刘畅,杜国栋,吕德国. 生长调节剂及钙、硼肥对南果梨萼片脱落、果实品质及木质素代谢的影响[J]. 沈阳农业大学学报,2019,50(4):399-405.
WANG Xiaoqian,SHANG Ye,LIU Weicheng,LIU Chang,DU Guodong,L? Deguo. Effect of growth regulators,Ca and B fertilizers on calyx abscission,fruit quality,and lignin metabolism in Nanguo Pear[J]. Journal of Shenyang Agricultural University,2019,50(4):399-405.
[82] CHEN Y T,MAO W W,LIU T,FENG Q Q,LI L,LI B B. Genome editing as a versatile tool to improve horticultural crop qualities[J]. Horticultural Plant Journal,2020,6(6):372-384.
猜你喜欢
纺织科学研究(2023年12期)2023-04-20
造纸信息(2022年8期)2022-11-10
轮胎工业(2021年4期)2021-12-25
上海包装(2019年8期)2019-11-11
中国造纸(2019年6期)2019-09-10
天然产物研究与开发(2018年10期)2018-11-06
天津造纸(2016年1期)2017-01-15
中国塑料(2016年6期)2016-06-27
中国造纸学报(2015年1期)2015-12-16
中国塑料(2014年4期)2014-10-17
