
浙江省常见15个树种的光合特性
2012-11-24 07:08刘欣欣张明如温国胜邹伶俐
浙江农林大学学报 2012年2期
刘欣欣,张明如,温国胜,张 俊,邹伶俐,吴 刚
(1.浙江农林大学 林业与生物技术学院,浙江 临安 311300;2.浙江农林大学 园林与建筑学院,浙江临安 311300;3.浙江农林大学 旅游与健康学院,浙江 临安311300)
马尾松Pinus massoniana群落是浙江省森林旅游区低山丘陵地段分布面积较大的一类森林群落类型。调控现有马尾松单优群落的结构组成,是恢复亚热带低山丘陵区地带性森林植被的重要任务之一。丁圣彦等[1]研究结果认为:在亚热带森林植被进展演替过程中,马尾松的衰退在于木荷Schima superba,栲Castanopsis fargesii等耐荫性阔叶树种的抑制作用,结果降低了马尾松的种间竞争强度,引起马尾松逐渐从原有植物群落中消失。不可否认的是,在马尾松退化群落中,存在光能资源未被充分利用的现象。为了促进马尾松群落的进展演替和提高光资源的利用效率,需要对马尾松退化群落进行阔叶化改造,在马尾松退化森林群落下层引植阔叶树种,合理配置形成复层结构以加速地带性森林植被的恢复进程。前人已经就亚热带地区树种的光能利用特征进行了卓有成效的研究,如在天童国家森林公园[2]、台州地区[3]和重庆地区[4]等,从光合生理生态角度探讨了亚热带地区群落演替各个阶段木荷,栲,东南石栎Lithocarpus harlandii和马尾松等优势种的特性。但是还需对上述树种在不同季节的光合特性作进一步研究。因此,本研究通过测定来研究比较浙江省常见15个树种苗期的光能利用能力以及耐荫性的特点,解释不同树种利用光能的差异,并对结果进行排序,以此来确定树种在退化群落中的配置,以期为浙江省低山丘陵区合理恢复退化植被,优化群落结构提供理论依据。
1 试验材料和方法
1.1 试验材料
试验材料为15个树种2~3年生盆栽苗,分别为苦槠Castanopsis sclerophylla,木荷,杜英Elaeocarpus decipiens,乌桕Sapium sebiferum,东南石栎,石栎Lithocarpus glaber,浙江楠Phoebe chekiangensis,乌冈Quercus phillyraeoides,竹柏Podocarpus nagi,马尾松,水杉Metasequoia glyptostroboide,红豆杉Taxus chinensis,红叶石楠Photinia×fraseri,美丽胡枝子Lespedeza formosa,红花木Loropetalum chinense,各个树种为3个盆栽苗,植株健壮。
1.2 光合参数日变化测定
于2010年11月7-8日(晴天),利用Licor-6400便携式光合仪,采用透明叶室测定各树种盆栽苗中上部第3~5片成熟功能叶,从8:30-16:30隔2 h同步测定净光合速率、光合有效辐射、气温、胞间二氧化碳摩尔分数、气孔导度、蒸腾速率等参数,重复3次°树种-1,测定1盆°次-1,取测定结果的平均值。
1.3 光响应过程测定
于2010年11月4-6日(晴天),采用开放式气路,利用Licor-6400便携式光合仪的LED红蓝光源依次设定光量子通量密度为 2 000,1 500,1 200,1 000,800,500,300,100,50,30,10 和 0 μmol°m-2°s-1,选择各树种盆栽苗中上部第3~5片成熟功能叶进行测定,重复3盆°树种-1,测1片°盆-1,重复3次°叶片-1。用SPSS 13.0拟合求得最大净光合速率、光补偿点和光饱和点。
1.4 数据处理
运用Excel 2003软件进行数据处理,Origin8作图,采用SPSS13.0进行统计分析。光能利用效率(LUE)计算公式[5]为: LUE=Pn/PAR。其中: Pn为净光合速率(μmol°m-2°s-1),PAR为光合有效辐射(μmol°m-2°s-1)。
2 结果与分析
2.1 光合作用日变化
2.1.1 阔叶树种的光合参数日变化 分析图1可知:9个树种的净光合速率Pn在测定时段的变化均为单峰型,峰值出现在12:30,而乌桕的峰值出现10:30。阔叶树种Pn峰值大小顺序为乌桕>杜英>木荷>东南>石栎>竹柏>浙江楠>苦槠>乌冈。石栎的气孔导度在测定时段的变化属于双峰型,其他8个树种的气孔导度呈单峰型。9个树种气孔导度峰值大小顺序为乌桕>杜英>竹柏>东南石栎>乌冈>石栎>苦槠>木荷>浙江楠。东南石栎、苦槠、乌冈、浙江楠的蒸腾速率日变化的2个峰值分别出现在10:30和14:30;乌桕、石栎、木荷、竹柏的蒸腾速率日变化呈先上升后下降的变化趋势,蒸腾速率峰值大小顺序为:乌桕>石栎>竹柏>木荷;杜英的蒸腾速率变化较为平缓。9个树种的胞间二氧化碳摩尔分数与气孔导度呈相反变化趋势。东南石栎、杜英、木荷、浙江楠、竹柏的胞间二氧化碳摩尔分数日变化呈先下降后上升的特征;乌冈胞间二氧化碳摩尔分数于10:30出现峰值;石栎的胞间二氧化碳摩尔分数在14:30出现峰值;苦槠的胞间二氧化碳摩尔分数变化相对平缓。9个树种的胞间二氧化碳摩尔分数峰值大小排序为:东南石栎>浙江楠>木荷>杜英>石栎>竹柏>乌桕>乌冈>苦槠。
2.1.2 针叶树种的光合参数日变化 红豆杉、马尾松、水杉的净光合速率日变化均呈单峰型,其大小顺序为红豆杉>马尾松>水杉(图2),且红豆杉的净光合速率高于马尾松和水杉;马尾松的气孔导度呈典型的单峰型,峰值出现在12:30,红豆杉和水杉的气孔导度在10:30出现峰值;蒸腾速率峰值大小顺序为马尾松>红豆杉>水杉;胞间二氧化碳摩尔分数日变化均为双峰型,峰值分别出现在10:30和14:30。
2.1.3 灌木树种的光合参数日变化 灌木树种的净光合速率、气孔导度、蒸腾速率日变化均属于单峰型(图3)。净光合速率和气孔导度峰值均出现在12:30,净光合速率峰值大小顺序为美丽胡枝子>红叶石楠>红花木;气孔导度峰值大小顺序为:红叶石楠>美丽胡枝子>红花木;蒸腾速率峰值出现在10:30,其大小排序为美丽胡枝子>红叶石楠>红花木;胞间二氧化碳摩尔分数与气孔导度变化趋势相反,谷值出现在12:30,谷值大小顺序为美丽胡枝子>红花木>红叶石楠。所测15个树种的净光合速率日变化均呈单峰型,未出现 “光合午休”现象。东南石栎、苦槠、乌冈、浙江楠、马尾松、红花木的蒸腾速率均呈双峰型,其余树种呈单峰型。针叶和灌木树种的胞间二氧化碳摩尔分数总体上要比阔叶树种的高。
2.2 光响应过程测定
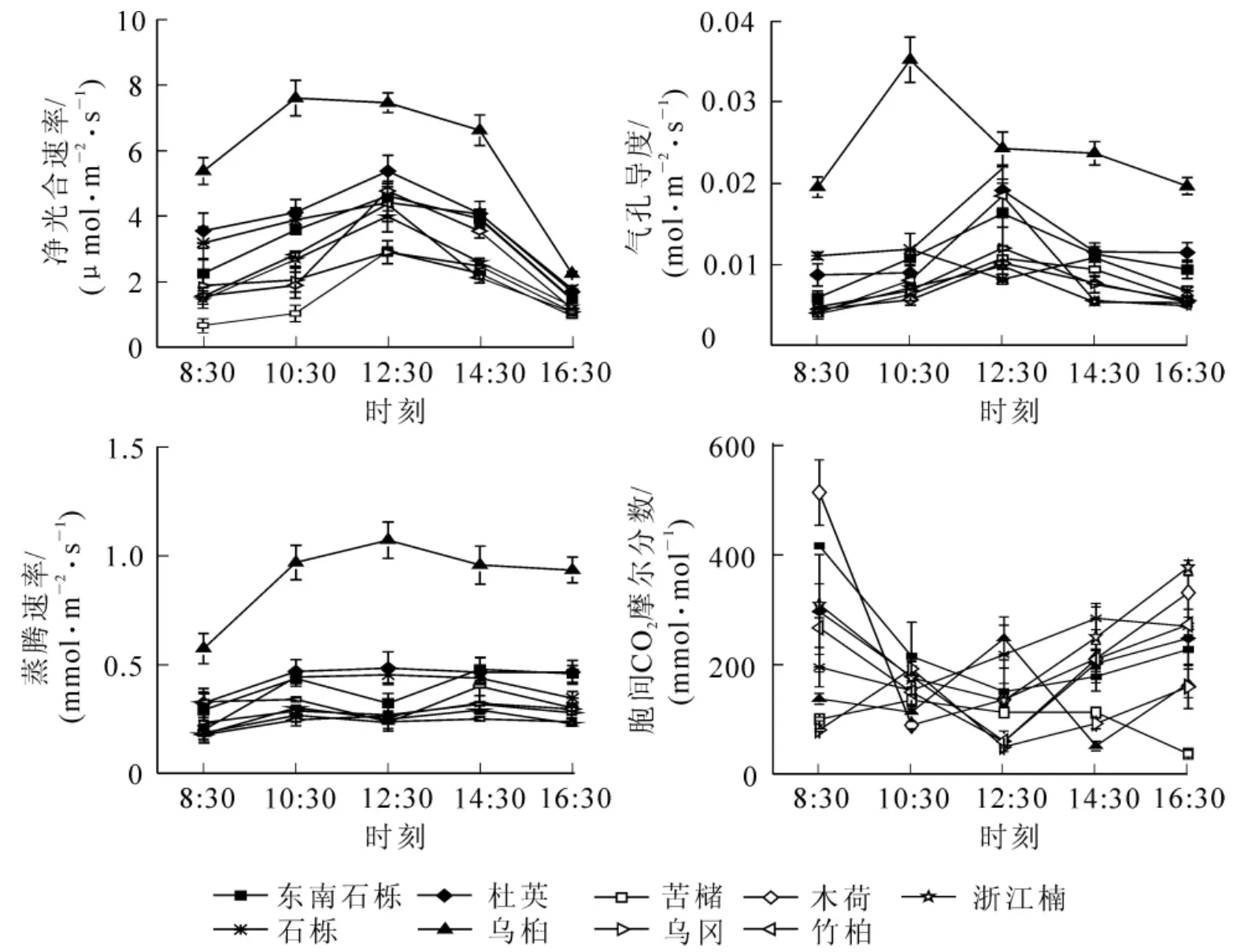
图1 9种阔叶树种光合参数日变化Figure 1 Diurnal changes of photosynthetic parameters of nine broadleaf tree species
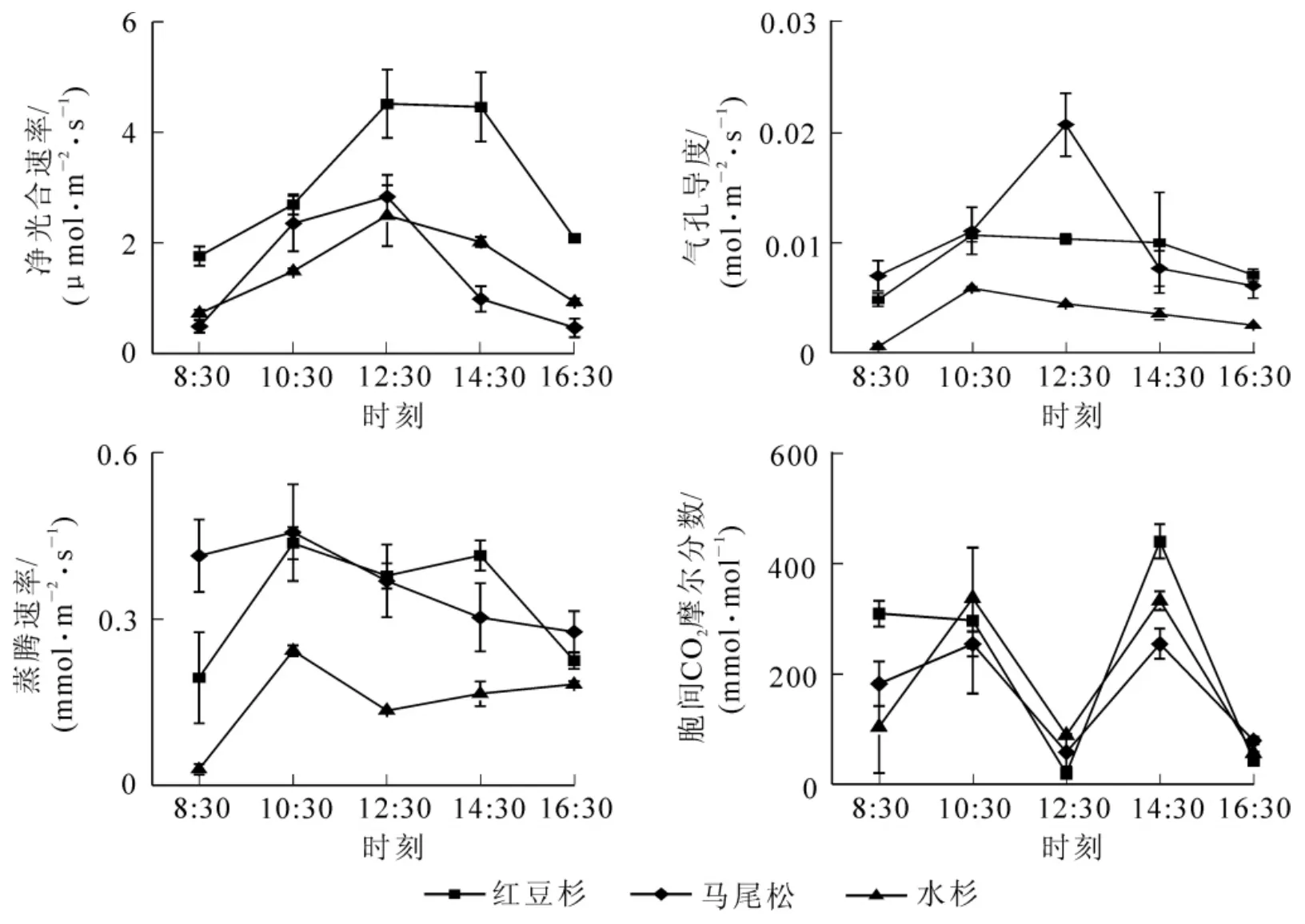
图2 3种针叶树种光合参数日变化Figure 2 Diurnal changes of photosynthetic parameters of three coniferous species
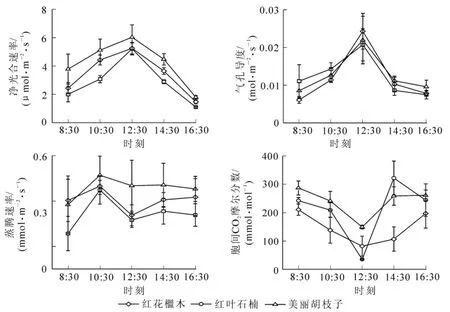
图3 3种灌木树种光合参数日变化Figure 3 Diurnal changes of photosynthetic parameters of three shrub species
2.2.1 阔叶、针叶及灌木树种的光响应过程测定 9个树种的光响应特征参数如表1所示。通过SPSS 13.0 拟合后认为苦槠的光饱和点最大,达 1 780.9 μmol°m-2°s-1,浙江楠最小 1 310.4 μmol°m-2°s-1,9 个阔叶树种光饱和点排序为苦槠>东南石栎>竹柏>乌桕>杜英>石栎>木荷>乌冈>浙江楠;光补偿点由大到小排序为乌冈>东南石栎>苦槠>浙江楠>杜英>木荷>乌桕>竹柏>石栎,其中乌冈光补偿点达87.9 μmol°m-2°s-1,而石栎仅为 4.1 μmol°m-2°s-1。光补偿点的高低直接反映植物对弱光的利用能力大小。一般认为杜英是较耐荫树种,但是本研究中杜英的光补偿点偏高,可能是幼苗和成树之间光合生理特性有所区别。最大净光合速率的排序为苦槠>乌桕>东南石栎>木荷>竹柏>杜英>乌冈>石栎>浙江楠。3个针叶树种的光饱和点大小排序为水杉>红豆杉>马尾松;光补偿点大小排序为马尾松>水杉>红豆杉;红豆杉的最大净光合速率最高,为65.9 μmol°m-2°s-1,马尾松和水杉最大净光合速率比较接近,分别为 34.4 μmol°m-2°s-1和 38.9 μmol°m-2°s-1(表1)。由表1 可知: 3 种灌木的光饱和点和最大净光合速率大小顺序为美丽胡枝子>红花木>红叶石楠;光补偿点为美丽胡枝子>红叶石楠>红花木。
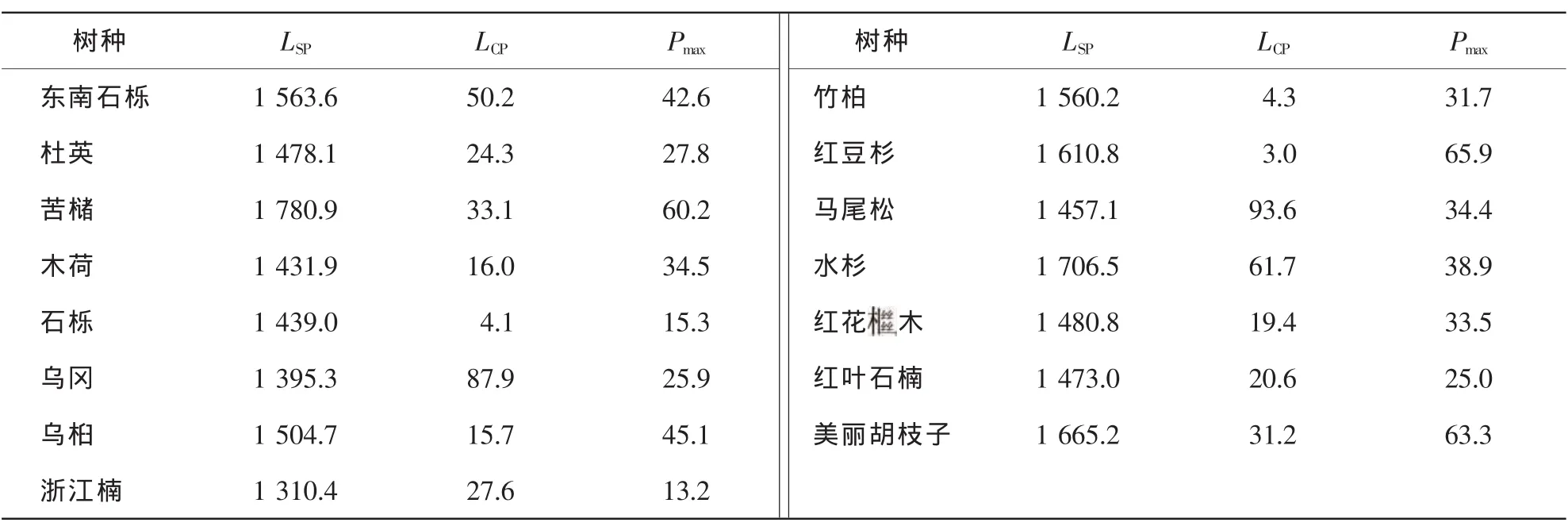
表1 15个树种光饱和点LSP,光补偿点(LCP)及最大净光合速率(Pmax)Table 1 Light saturation point (LSP),light compensation point (LCP) and maximum net photosynthetic rate (Pmax) of fifteen tree species
2.2.2 测定树种耐荫性聚类分析 利用SPSS13.0对15个树种的光补偿点和光饱和点进行聚类分析,结果如图4所示。乌冈、马尾松属于强阳性树种;苦槠、美丽胡枝子、水杉、东南石栎为阳性树种;红花木、红叶石楠、杜英、乌桕、木荷、石栎、竹柏、红豆杉、浙江楠归类为耐荫树种,对弱光的利用能力较强。
2.3 光能利用效率分析

图4 15个树种耐荫性聚类分析结果Figure 4 Cluster analysis of fifteen tree species shade tolerance
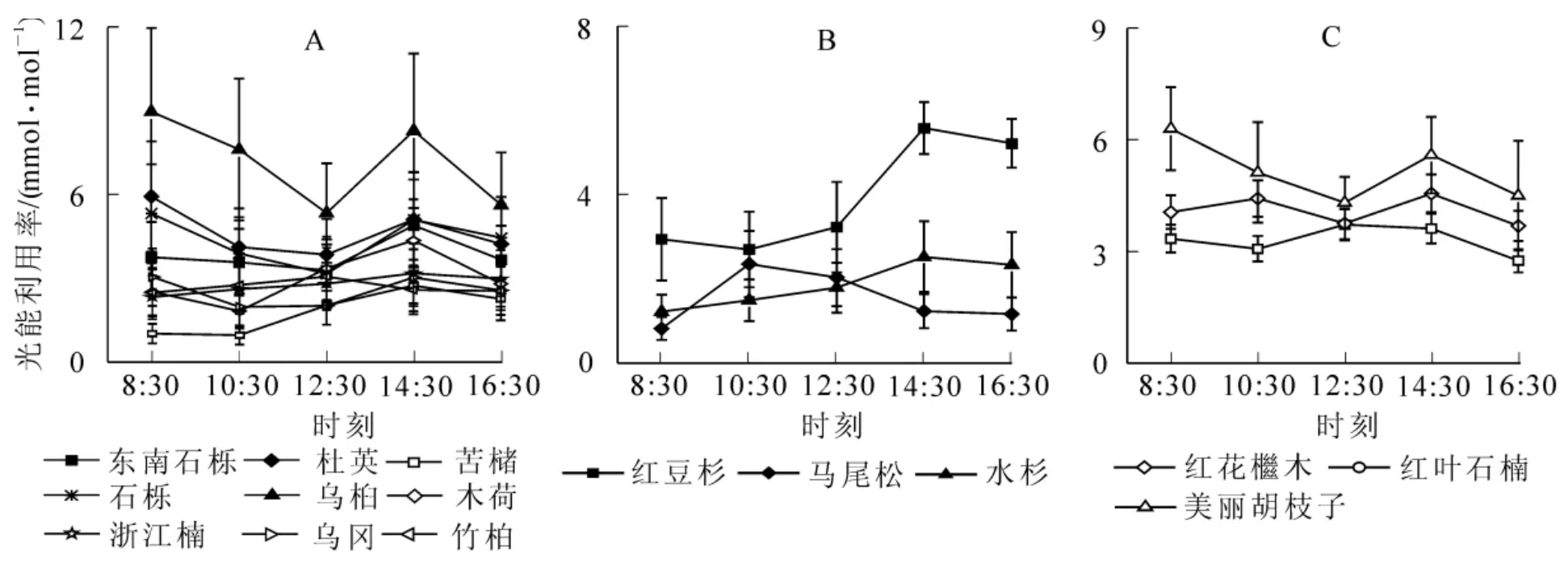
图5 9种阔(A),3种针叶(B)和3种灌木(C)树种光能利用率日变化Figure 5 Diurnal variation of broadleaf,coniferous and shrub species instantaneous light use efficiency
由图5-A可知:东南石栎、苦槠、木荷、浙江楠的光能利用效率在14:30出现峰值;杜英、石栎、乌桕、乌冈光能利用率的峰值出现在8:30,竹柏则出现在12:30。9个树种光能利用效率峰值大小排序为东南石栎>乌桕>杜英>石栎>木荷>浙江楠>竹柏>乌冈>苦槠。分析图5-B可知:马尾松和水杉的光能利用率的峰值分别为出现在10:30和14:30;红豆杉的光能利用率在测定时段均高于马尾松和水杉,峰值为5.6 mmol°mol-1。由图5-C可知,美丽胡枝子的光能利用率高于红花木和红叶石楠,8:30为测定时段的最高值,达6.3 mmol°mol-1,在12:30出现谷值;红花木的光能利用率谷值为 3.1 mmol°mol-1,峰值为3.6 mmol°mol-1;红叶石楠的光能利用率属于双峰型,峰值分别出现在10:30和14:30。灌木树种的光能利用率峰值由大到小为:美丽胡枝子>红叶石楠>红花木。
3 讨论
光能资源利用是植被稳定分布的重要问题之一。马尾松退化生境的光资源剩余现象显著,耐荫性树种因无法适应退化生境光环境而逐渐消失,阳性树种如马尾松种子因其喜光特性和较高的氮营养转移效率[6]成功入侵成林[7]并形成单优群落。江波等[8]认为应将马尾松林退化群落改造成具有良好结构功能和高效稳定多树种结构的阔松混交林,考虑到森林群落是乔木、灌木和草本植物的复合体,对马尾松退化群落的改造应强调乔、灌、草复层结构的优化配置、树种的生态互补关系以及芒萁Dicranopteris dichotoma单优层片的调控措施[9],因此,树种选择是低山丘陵地区马尾松退化群落恢复的关键技术之一[10-11],所选树种应适应马尾松退化群落下层光环境并能加速森林群落的进展演替[12]。
不同的树种对光的适应、利用显然是不同的。光照强度的增加导致常绿阔叶树种比叶面积降低,最大净光合速率和叶片氮含量增加[13]。常绿阔叶林在演替的次生灌丛阶段,日平均光合作用速率以木荷为最大,石栎其次,马尾松幼树的日平均光合作用速率低于前两者[14]。本研究苗期阶段秋季晴天测定的结果证实: 日平均光合速率为石栎(3.5 μmol°m-2°s-1)>木荷(2.5 μmol°m-2°s-1)>马尾松(1.4 μmol°m-2°s-1)。这可能与测定树种生长的阶段及季节有关,并且木荷幼苗早期具有很强的耐荫性,生长速率慢,可长时间忍耐低光照环境[15],而石栎是中性偏阳性树种,生态幅较大[14]。柏明娥等[16]研究了美丽胡枝子夏季光合作用特性,认为美丽胡枝子为强阳性速生植物,与本研究的结果一致,但本研究得出的光补偿点和光饱和点高于柏明娥等的结果;柯世省等[3]认为东南石栎的光补偿点较低、光饱和点较高。本研究中,东南石栎的光补偿点较高。
基于15个树种光补偿点和光饱和点的聚类结果认为:乌冈、马尾松为强阳性树种,可以种植在全光环境下;东南石栎、苦槠、水杉和美丽胡枝子归属于阳性树种,应配置于疏林,灌丛下;杜英、木荷、石栎、乌桕、竹柏、浙江楠、红豆杉、红花木和红叶石楠归类为耐荫树种,适合配置在林冠下。然而树种的光能利用特征受树种本身特性、发育阶段、季节变化、小气候和土壤肥力状况等多种因素的影响。我们测定分析的结果仅仅是针对测定树种苗期于秋季的光响应特征,所以研究尺度应该拓宽,如需要测定冬、春、夏3个季节的幼苗、幼树乃至成熟植株的光响应特征。此外,测定树种具有雌雄异株的特性时,光合响应特征亦会发生一定的变化。另外,本研究仅针对光能利用进行讨论,为了进一步利用退化群落中的资源,调整群落树种组成,植物的水分利用能力及植物对土壤养分利用能力是今后需深化研究的问题。
[1]丁圣彦,宋永昌.常绿阔叶林演替过程中马尾松消退的原因[J].植物学报,1998,40(8):755-760.DING Shengyan,SONG Yongchang.Declining causes of Pinus massoniana in the progresses of succession of evergreen broad leaved forest[J].Acta Bot Sin,1998,40 (8): 755-760.
[2]丁圣彦.浙江天童常绿阔叶林演替系列栲树和木荷成为优势种的原因[J].河南大学学报,2001,31(1):79-83.DING Shengyan.The causes of Castanopsis fargesii and Schima superba being dominant species of series of evergreen broad-leaved forest in Zhejiang Tiantong [J].J Henan Univ Nat Sci,2001,31 (1): 79-83.
[3]柯世省,金则新,林恒琴,等.天台山东南石栎光合生理生态特性[J].生态学杂志,2004,23(3):1-5.KE Shisheng,JIN Zexin,LIN Hengqin,et al.Photosynthetic ecophysiological characteristics of Lithocarpus harlandii in Tiantai Mountain of Zhejiang Province,China [J].Chin J Ecol,2004,23 (3): 1-5.
[4]王周平,李旭光,石胜友,等.重庆缙云山针阔混交林林隙树木更替规律研究[J].植物生态学报,2001,25(4):399-404.WANG Zhouping,LI Xuguang,SHI Shengyou,et al.Gap phase tree replacement in a conifer-broadleaved mixed forest in Jinyun Mountain,Chongqing [J].Acta Phytoecol Sin,2001,25 (4): 399-404.
[5]JENKINS J P,RICHARDSON A D,BRASWELL B H,et al.Refining light-use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radiometric measurements [J].Agric For Meteorol,2007,143: 64-79.
[6]王希华,黄建军,闫恩荣.天童常绿阔叶林若干树种的叶片营养转移研究[J].广西植物,2004,24(1):81-85.WANG Xihua,HUANG Jianjun,YAN Enrong.A study on leaf nutrient resorption of some trees in Tiantong National Forest Park [J].Guihaia,2004,24 (1): 81-85.
[7]洪利兴,王泳,杜国坚,等.我国南方马尾松林生态系统的退化特征和改造对策研究[J].浙江林业科技,2000,20 (2): 1-9.HONG Lixing,WANG Yong,DU Guojian,et al.Retrograde characteristics of ecosystem of Pinus massoniana forest and its transformation countermeasures [J].J Zhejiang For Sci Technol,2000,20 (2): 1-9.
[8]江波,袁位高,朱锦茹,等.森林生态体系快速构建理论与技术研究[M].北京:中国林业出版社,2010:182-185.
[9]张明如,温国胜,张汝民,等.千岛湖森林群落下层芒萁层片发育机理初步判断 [J].内蒙古农业大学学报:自然科学版,2010,31(3): 28-34.ZHANG Mingru,WEN Guosheng,ZHANG Rumin,et al.A study of the development mechanisms of Dicranopteris dichotoma synusium of the forest communities in Thousands Lake [J].J Inner Mongolia Agric Univ Nat Sci Ed,2010,31(3): 28-34.
[10]王希华,宋永昌.马尾松林恢复为常绿阔叶林的研究[J].生态学杂志,2001,20(1):30-32.WANG Xihua,SONG Yongchang.Study on restoration of evergreen broad-leaved forest from Pinus massoniana forest[J].Chin J Ecol,2001,20 (1): 30-32.
[11]赵平.退化生态系统植被恢复的生理生态学研究进展[J].应用生态学报,2003,14(11):2031-2036.ZHAO Ping.Advances in plant ecophysiological studies on re-vegetation of degraded ecosystems [J].Chin J Appl Ecol,2003,14 (11): 2031-2036.
[12]达良俊,宋坤.浙江天童受损常绿阔叶林实验生态学研究(Ⅰ)生态恢复实验与长期定位[J].华东师范大学学报:自然科学版,2008(4):1-11.DA Liangjun,SONG Kun.Experimental ecology research on destroyed evergreen broad-leaved forests in TNFP,Zhejiang (Ⅰ) Ecological restoration experiments and long-term ecological study [J].J East China Norm Univ Nat Sci,2008 (4): 1-11.
[13]康明敏,张奇平,达良俊,等.浙江天童受损常绿阔叶林实验生态学研究(Ⅵ)不同干扰下植被恢复初期主要优势种叶性状及其生态适应性[J].华东师范大学学报:自然科学版,2010(3):26-38.KANG Minming,ZHANG Qiping,DA Lianjun,et al.Experimental ecology research on destroyed evergreen broadleaved forest in TNFP,Zhejiang(Ⅵ)leaf traits and ecologcal adaptation of five main dominant species during the early stage restoration after different disturbances [J].J Eastchina Norm Univ Nat Sci,2010 (3): 26-38.
[14]丁圣彦.常绿阔叶林演替系列比较生态学[M].开封:河南大学出版社,1999:30.
[15]宋坤,杨徐烽,达良俊,等.浙江天童受损常绿阔叶林实验生态学研究 (Ⅱ)主要常绿树种的生长格局[J].华东师范大学学报:自然科学版,2008(4):12-24.SONG Kun,YANG Xufeng,DA Liangjun,et al.Experimental ecology research on destroyed evergreen broad-leaved forests in TNFP,Zhejiang (Ⅱ) The growth patterns of dominant evergreen trees determined by tree ring analysis[J].J East China Norm Univ Nat Sci,2008 (4): 12-24.
[16]柏明娥,洪利兴,朱汤军,等.美丽胡枝子光合作用特性及其影响因子分析 [J].浙江林业科技,2007,27(6):1-5.BO Minge,HONG Lixing,ZHU Tangjun,et al.Photosynthetic characteristics of Lespedeza formosa and influencing factors [J].J Zhejiang For Sci Technol,2007,27 (6): 1-5.
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
林业科学(2022年5期)2022-08-20
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
花卉(2021年2期)2021-01-21
农家科技中旬版(2020年10期)2020-07-12
农业与技术(2020年12期)2020-07-04
农家科技中旬版(2020年2期)2020-03-18
绿色科技(2019年5期)2019-11-29
农学学报(2019年1期)2019-02-21
