
小麦谷胱甘肽S-转移酶基因的克隆及原核表达
2016-07-28 01:36于永昂杨天佑
华北农学报 2016年3期
张 蕾,于永昂,2,杨天佑
(1.河南科技学院生命科技学院,现代生物育种河南省协同创新中心,河南新乡 453003;2.西北农林科技大学农学院,陕西杨凌 712100)
小麦谷胱甘肽S-转移酶基因的克隆及原核表达
张 蕾1,于永昂1,2,杨天佑1
(1.河南科技学院生命科技学院,现代生物育种河南省协同创新中心,河南新乡 453003;2.西北农林科技大学农学院,陕西杨凌 712100)
为了进一步研究小麦谷胱甘肽-S-转移酶基因(TaGST)的功能,采用RT-PCR方法分离了小麦谷胱甘肽-S-转移酶基因(TaGST)的ORF全长cDNA,并进行了生物信息学分析。结果表明:小麦TaGST基因的ORF全长690 bp,编码229个氨基酸;TaGST蛋白分子质量为25.81 kDa,p I为5.29。系统进化分析表明,该基因编码蛋白与水稻OsGST蛋白的氨基酸同源性最高,与已知植物GST家族成员的氨基酸序列聚类分析将TaGST聚为Phi类GST。构建原核表达载体pET32-TaGST,对TaGST基因进行原核表达,SDS-PAGE结果表明,其所表达蛋白与预期蛋白大小一致。为进一步研究该基因的特性和功能奠定了理论基础。
小麦;谷胱甘肽过S-转移酶;基因克隆;原核表达
谷胱甘肽-S-转移酶(Glutathione-S-transferase,GSTs)是普遍存在于动植物中的一种多基因编码的多功能蛋白酶,在清除生物和非生物胁迫产生的氧化损伤中起着重要的作用[1-2],其一般是由同源或异源二聚体蛋白组成的2个亚基来催化还原型谷胱甘肽与亲电的化合物进行结合反应,降低其亲电活性,最终达到脱毒作用[3-4]。目前,已经在拟南芥、水稻、杨树、玉米和大豆等植物中发现了多种GST蛋白,其中在拟南芥、水稻和杨树中就分别发现了53,59,81个GST蛋白[5-7]。根据蛋白的相似性,将GST主要分为7类:Phi(F)、Tau(U)、Theta(T)、Zeta(Z)、Lambda(L)、DHAR(Dehydroascorbate reductase)和TCHQD(Tetrachlorohydroquinone dehalopgenase),其中,Phi、Tau、Lambda和DHAR类是植物所特有的[8-10]。
GSTs在不同组织、不同发育时期的表达量有差异,并且其表达能够受多种环境因子的诱导,如低
温[11]、脱水[12]、伤害[13]和病原菌的侵染[14]等,在植物的生长发育、次生代谢和胁迫耐受中起着重要作用。许多研究表明,GST在植物抗性中起着重要的作用[15-18]。烟草中过量表达烟草GST基因提高了转基因烟草的耐盐和抗低温能力[15],而将棉花GST基因转入烟草增强了转基因烟草的抗氧化胁迫能力[16]。在水稻中过量表达盐地碱蓬GST基因可以提高转基因水稻的抗低温能力[17],而在拟南芥中过量表达盐地碱蓬GST基因可以提高转基因拟南芥的抗旱能力[18]。小麦是我国重要的粮食作物,但由于干旱、低温和盐碱等非生物胁迫严重影响了小麦产量。克隆小麦抗逆基因TaGST对于进一步利用该基因提高小麦抗逆性具有重要意义。为此,从小麦叶片中克隆了TaGST基因全长序列,并利用生物信息学技术对该基因编码蛋白的理化性质进行了分析。同时,构建了pET32-TaGST原核表达载体,在宿主菌E.coli BL21(DE3)中成功进行了诱导表达,为小麦TaGST蛋白的进一步分离纯化以及结构和功能研究奠定理论基础。
1 材料和方法
1.1 试验材料
供试小麦品种为新麦26,将小麦种子置于铺有滤纸的培养皿中,置于培养箱中培养,三叶期时,取新鲜幼嫩叶片用液氮速冻待用;原核表达载体pET-32a购自德国Novagen公司;E.coli DH5α感受态细胞为河南科技学院发酵工程实验室保存;限制性内切酶Bam HⅠ和Hin dⅢ、T4DNA连接酶购自大连TaKaRa公司;表达菌株E.coli BL21(DE3)购自北京全式金生物技术有限公司;其他试剂均为国产分析纯。
1.2 试验方法
1.2.1 TaGST基因的克隆 采用TRIzol-A+(Tiangen,北京)试剂提取样品总RNA,按说明书使用Prim Script RT Reagent Kit(TaTaRa,大连)反转录合成第一链cDNA。根据小麦TaGST(GenBank登录号:X56012)基因序列,用Primer Premier 5.0设计特异性引物GST-F1:5′-ATGTCTCCGGTGAAGGTG-3′,GST-R1:5′-CTAGTACTGCGCACCTAG-3′。以合成的cDNA第一链为模板进行PCR扩增。PCR反应条件为:94℃预变性3 min;94℃30 s,56℃30 s,72℃40 s,35个循环;72℃延伸5 min。PCR反应结束后产物经电泳检测,回收与目标片段大小一致的条带,将其连接到pGM-T载体(Tiangen,北京),转化E.coli DH5α感受态细胞,挑取白色菌斑摇菌,进行菌液PCR扩增,将鉴定正确的阳性菌株进行测序。
1.2.2 小麦TaGST序列的生物信息学分析 通过Protparam分析其编码蛋白的理化性质;将TaGST基因序列和推导的氨基酸序列在NCBI(http://www. ncbi.nlm.nih.gov)上用BLASTX进行同源性搜索和比对;用DNAMAN 6.0进行多重序列比对;用TMHMM预测蛋白质跨膜区结构;用SingnalP4.1软件进行信号肽预测;用Mega 5.0软件构建系统发育树。
1.2.3 TaGST基因原核表达载体的构建 根据原核表达载体pET-32a和TaGST基因序列设计带有Bam HⅠ和Hin dⅢ酶切位点的特异性引物用于TaGST基因原核表达载体的构建,引物序列如下:GST-F2:5′-CGGGATCCATGTCTCCGGTGAAGGTG-3′(下划线为Bam HⅠ酶切位点),GST-R2:5′-CCCAAGCTTGTACTGCGCACCTAGCTT-3′(下划线为Hin dⅢ酶切位点)。
PCR产物经回收后连接到pGM-T载体,转化E.coli DH5α感受态细胞,涂布于含有Amp的LB固体培养基上,37℃培养过夜,挑取单克隆进行PCR扩增和双酶切鉴定,并进行测序鉴定,以获得重组质粒pGM-TaGST。将测序鉴定正确的pGM-TaGST质粒与原核表达载体pET-32a分别用Bam HⅠ和Hin dⅢ进行双酶切,酶切产物经T4DNA连接酶连接后热击转化E.coli DH5α感受态细胞,在LB(Amp+)固体培养基上培养12 h后,挑取阳性克隆提取质粒,进行PCR扩增和双酶切鉴定,以获得原核表达载体pET32-TaGST。
1.2.4 TaGST基因的原核表达 将原核表达载体pET32-TaGST转化E.coli BL21(DE3)感受态细胞,挑取单克隆至10 m L LB(Amp+)液体培养基中,200 r/min 37℃振荡培养12 h。然后按照1∶100(V/V)转接到新鲜LB液体培养基中培养2~3 h至OD600达到0.6~0.8,取1 mL重组质粒菌液加IPTG至终浓度为0.5 mmol/L,37℃、200 r/min诱导培养1,2,3,4 h。诱导表达完成后,取1 m L菌液,12 000 r/min离心10 min收集菌体,进行SDS-PAGE电泳,检测融合蛋白的表达情况。凝胶用考马斯亮蓝R-250染色并脱色至条带清晰。
2 结果与分析
2.1 TaGPX基因的克隆
根据NCBI中小麦GST基因的序列设计特异性扩增引物,采用RT-PCR技术扩增得到1条大约700 bp的条带(图1),测序结果显示该序列长690 bp,与预期片段大小一致,编码229个氨基酸,起始密码子为ATG,终止密码子为TAG(图2)。
2.2 小麦TaGST序列的生物信息学分析
利用Protparam分析TaGST的理化性质,推测TaGST蛋白的分子式为C1188H1816N296O326S11,分子质量为25.81 kDa,理论等电点(p I)为5.29。TaGST蛋白不稳定系数为46.10(高于域值40),属于不稳定蛋白。总的带负电荷的残基(Asp+Glu)为28,总的带正电荷的残基(Arg+Lys)为21。TMHMM和SignalP 4.1在线分析结果表明,该蛋白没有跨膜结构域和信号肽,属于非分泌蛋白。TaGST蛋白的结构域预测发现其保守结构域分为N端(位于3-78氨基酸残基)和C端(位于95-216氨基酸残基)两部分(图3)。

图1 TaGST基因的PCR扩增Fig.1 PCR am p lification of TaGST gene
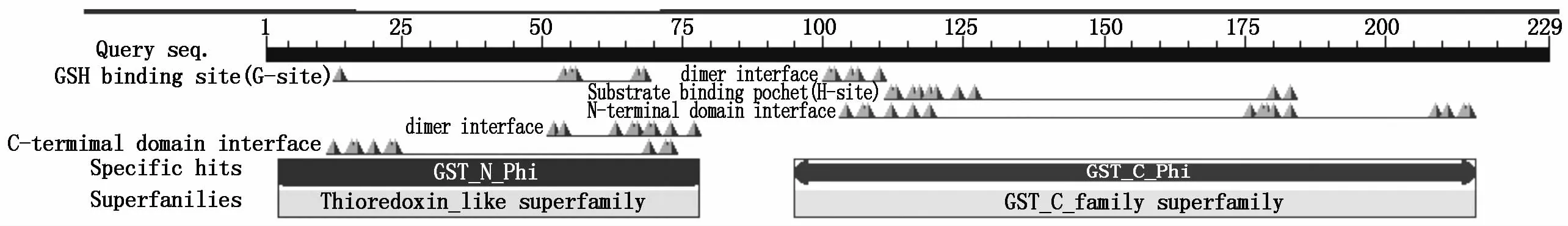
图3 TaGST保守结构域分析Fig.3 Conserved dom ain analysis of TaGST
利用DNAMAN 6.0软件,将克隆的TaGST基因编码的氨基酸序列与GenBank中其他植物的GST氨基酸序列进行比对,发现该基因编码蛋白与已知的水稻(BAB39939)、大麦(AAL733940)、玉米(AAA33469)、拟南芥(CAB83126)的Phi类GST具有较高的同源性,氨基酸相似性分别达到61.57%,61.14%,49.78%,43.67%(图4)。进一步利用Mega 5.0软件对小麦TaGST蛋白与水稻、拟南芥的Phi、Theta、Zeta、Lambda、Tau类GST蛋白序列构建系统发育树,结果表明,本研究所克隆的TaGST基因编码的蛋白和水稻、拟南芥的Phi类GST的亲缘 关系最近(图5)。

图4 TaGST基因编码的氨基酸序列与其他作物GST氨基酸序列的比对Fig.4 M u ltip le alignm ent of the deduced am ino acid sequences of TaGST and GST p roteins from other p lants
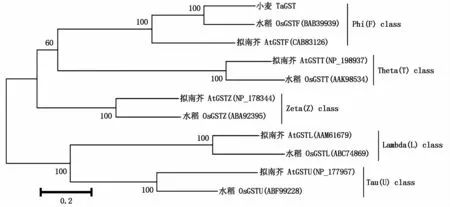
图5 五类GSTs进化树分析Fig.5 A phy logenetic tree constructed based on the am ino acid sequences of five classes of GSTs
2.3 TaGST基因原核表达载体的构建及酶切鉴定
通过Bam HⅠ和Hin dⅢ双酶切重组质粒pGMTaGST和pET-32a空载体,回收目的基因片段和线性载体进行连接,得到重组质粒pET32-TaGST,转化E.coli DH5α感受态细胞,抽提质粒进行双酶切鉴定,结果显示,TaGST基因成功连接到pET-32a载体上(图6)。重组质粒再次测序,结果表明,插入片段与克隆序列完全一致。由此表明pET32a-TaGST原核表达载体构建成功。

图6 小麦TaGST基因重组质粒pET32a-TaGST的双酶切鉴定Fig.6 Identification of the recom binan t vector pET32a-TaGST by Bam HⅠand H in dⅢdouble digestion
2.4 TaGST基因重组蛋白的诱导表达
由图7可知,经IPTG诱导的pET32a-TaGST重组质粒在E.coli BL21(DE3)中表达,分子质量约45 kDa,与预期的目标融合蛋白大小一致,目的蛋白约25.81 kDa,标签蛋白20.4 kDa,且随着诱导表达时间的增加融合蛋白表达量增多。
3 结论与讨论
植物在不利的非生物逆境胁迫条件下如高温、低温、干旱、盐渍等,会造成细胞代谢过程异常,积累大量的活性氧(ROS),影响植株的生长发育。在正常情况下,植物体内ROS的产生和清除处于一种动态的平衡状态,而当植物受到逆境胁迫时,大量积累的ROS能够对植物体内的脂质、蛋白质以及核酸等生物大分子产生伤害[19-21]。GST利用还原型谷胱甘肽的还原作用来清除逆境胁迫下细胞中产生的ROS,同时也能够对受到氧化损伤的细胞进行修复,从而维持植物体内的正常代谢过程。在瞿麦(Dianthus superbus L.)中过量表达烟草GST基因能够增加转基因瞿麦在干旱和不同光照条件下的光合作用[22]。火把梨的GST基因能够参与维持火把梨果实发育过程中的氧化还原平衡和应答逆境胁迫[23]。在模拟酸雨胁迫下,青花菜GST基因的表达量在胁迫初期显著增加,随着时间延长开始下降,表明GST基因参与了青花菜抗酸雨的应答反应[24]。这一系列研究表明,植物GSTs对抵抗逆境胁迫具有重要作用。
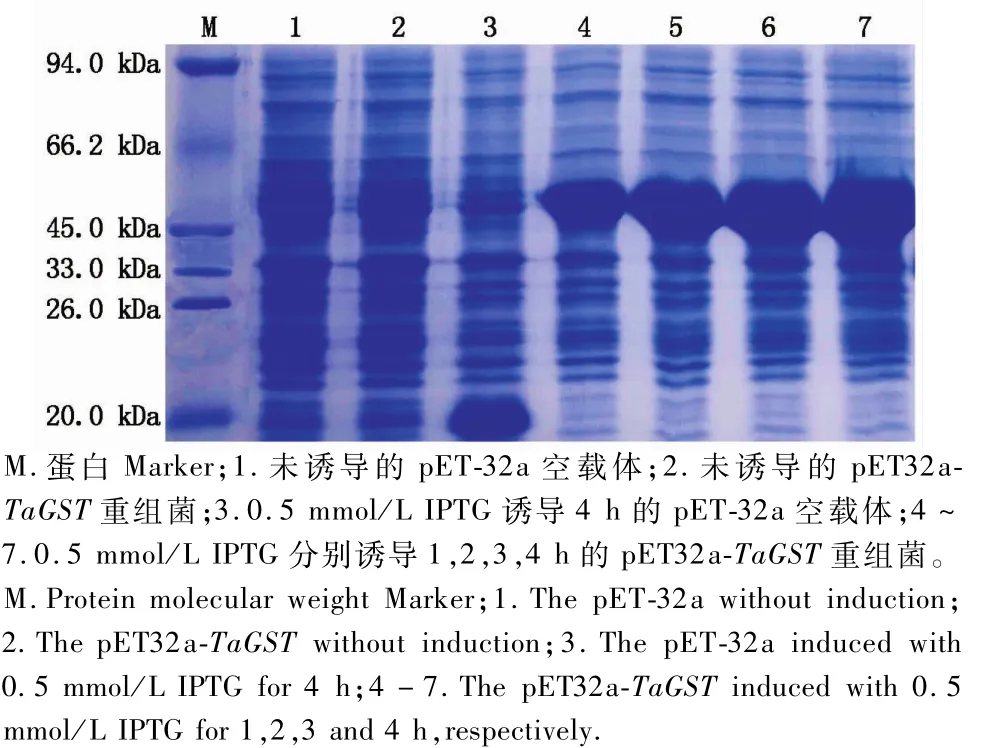
图7 pET32a-TaGST融合蛋白的表达分析Fig.7 Analysis of pET32a-TaGST p rotein exp ression in by SDS-PAGE
本试验根据NCBI中小麦TaGST基因序列,设计特异性引物,采用RT-PCR方法从小麦叶片中成功克隆得到具有完整ORF的TaGST基因序列;生物信息学分析表明,该基因所推导的氨基酸序列与已知的水稻、大麦、玉米和拟南芥的GST蛋白序列具有较高的相似性;保守结构域分析结果表明,所克隆的TaGST蛋白序列含有GST_N和GST_C,属于Phi类,同时聚类分析也将本试验中的TaGST归于Phi类GST。
原核表达技术常用于一些重要蛋白质的结构和功能研究。目前,在原核表达技术中应用最为广泛的表达宿主菌是大肠杆菌,其具有结构简单、操作方便以及对外源目的蛋白表达量高等多种优点[25-26]。本试验采用目前应用最广泛的pET系列原核表达载体进行目的蛋白的表达分析,成功构建了TaGST基因的原核表达载体pET32a-TaGST,并进一步将其转化E.coli BL21(DE3)感受态细胞进行目的蛋白的诱导表达。SDS-PAGE凝胶电泳分析表明,成功诱导出1条分子质量约为45 kDa的蛋白条带,与预测分子质量大小一致,为进一步纯化蛋白质以及进行生物学功能分析提供了基础资料。
综上所述,本试验从小麦叶片中成功地克隆了TaGST基因,利用原核表达技术成功诱导表达TaGST蛋白,为进一步研究TaGST蛋白的结构和功能以及通过遗传转化提高植物的抗逆性提供试验理论基础。
[1] Frova C.The plant glutathione transferase gene family:genomic structure,functions,expression and evolution[J]. Physiologia Plantarum,2003,119(4):469-479.
[2] George S,Venkataraman G,Parida A.A chlorop last-localized and auxin-induced glutathione S-transferase from phreatophyte Prosopis juliflora confer drought tolerance on tobacco[J].Journal of Plant Physiology,2010,167(4):311-318.
[3] Diao G P,Wang Y C,Wang C,et al.Cloning and functional characterization of a novel glutathione S-Transferase gene from limonium bicolor[J].Plant Molecular Biology Reporter,2011,29(1):77-87.
[4] Kumar S,Asif M H,Chakrabarty D,et al.Expression of a rice Lambda class of glutathione S-transferase,OsGSTL2,in Arabidopsis provides tolerance to heavy metal and other abiotic stresses[J].Journal of Hazardous Materials,2013,248-249,228-237.
[5] 范玉洁,林飞鹏,安泽伟,等.一个橡胶树谷胱甘肽-S-转移酶基因的克隆和表达特性分析[J].中国农业科学,2011,44(20):4150-4158.
[6] Soranzo N,Gorla A M,M izzi L,et al.Organisation and structural evolution of the rice glutathione S-transferase gene family[J].Molecular Genetics and Genomics,2004,271:511-521.
[7] Lan T,Yang Z L,Yang X,et al.Extensive functional diversification of the Populus glutathione S-transferase supergene family[J].The Plant Cell,2009,21:3749-3766.
[8] Edwards R,Dixon D P.Plant glutathione transferases[J]. Methods Enzymol,2005,401:169-186.
[9] 陈秀华,王臻昱,李先平,等.谷胱甘肽S-转移酶的研究进展[J].东北农业大学学报,2013,44(1):149-153.
[10] 易乐飞,郝 伟,李信书,等.条斑紫菜Mu型谷胱甘肽S-转移酶基因克隆与分析[J].水产科学,2016,35(1):67-71.
[11] Lo Piero A R,Puglisi I,Rapisarda P,et al.Anthocyanins accumulation and related gene expression in red orange fruit induced by low temperature storage[J].Journal of Agricultural and Food Chem istry,2005,53(23):9083-9088.
[12] Bianchi M W,Roux C,Vartanina N,et al.Drought regulation of GST8,ncodes the Arabidopsis homologue of ParC/Nt107 glutathione transferase/peroxidase[J]. Physiol Plant,2002,116:96-105.
[13] Vollenweider S,Weber H,Stolz S,et al.Fatty acid ketodienes and fatty acid ketotrienes:M ichael addition acceptors that accumulate in wounded and diseased Arabidopsis leaves[J].Plant Journal,2000,24(4):467-476.
[14] Mauch F,Dudler R.Differential induction of distinct glutathione S-transferases of wheat by xenobiotics and by pathogen attack[J].Plant Physiol,1993,102:1193-1201.
[15] Roxas V P,Roger K,Smith JR,et al.Over expression of glutathione S-transferase/glutathione peroxidase enhances the growth of transgenic tobacco seedlings during stress[J].Nat Biotech,1997,15:988-991.
[16] 於丙军,刘友良.盐胁迫对一年生盐生野大豆幼苗活性氧代谢的影响[J].西北植物学报,2003,23(1):18-22.
[17] 赵凤云,王晓云,赵彦修,等.转入盐地碱蓬谷胱甘肽转移酶和过氧化氢酶基因增强水稻幼苗对低温胁迫的抗性[J].植物生理与分子生物学学报,2006,32(2):231-238.
[18] 戚元成,张小强,刘卫群.过量表达谷胱甘肽转移酶基因对转基因拟南芥抗旱能力的影响[J].植物生理学通讯,2008,44(2):268-270.
[19] Chi Y H,Chen Y S,Vanitha J,et al.Expansion mechanisms and functional divergence of the glutathione stransferase family in Sorghum and other higher plants[J].DNA Research,2011,18:1-16.
[20] 毛小辉,魏毅东,张建福,等.粳稻品种云引谷胱甘肽S-转移酶基因OsGST的克隆及序列分析[J].福建农业学报,2014,29(3):197-203.
[21] Mohsenzadeh S,Esmaeili M,Moosavi F A,et al.Plant glutathione S-transferase classification,structure and evolution[J].African Journal of Biotechnology,2011,10(42):8160-8165.
[22] Lim JD,Hang S J,Yu C Y,et al.Expression of the glutathione S-transferase gene(NT107)in transgenic Dianthus superbus[J].Plant Cell Tissue Organ Cult,2005,80:277-286.
[23] 刘迪秋,王光勇,王继磊,等.火把梨谷胱甘肽S-转移酶基因的克隆与表达[J].西北植物学报,2012,32(1):29-34.
[24] 高世超,林义章,钟凤林,等.青花菜谷胱甘肽-S-转移酶基因克隆及其表达分析[J].西北植物学报,2014,34(4):651-657.
[25] Chen R.Bacterial expression systems for recombinant protein production:E.coli and beyond[J].Biotechnology Advances,2012,30(5,SI):1102-1107.
[26] 张晓东,李彩霞,王连春,等.滇龙胆GrSLS1基因的克隆与原核表达[J].西北植物学报,2014,34(7):1311-1317.
Cloning and Prokaryotic Expression of TaGST from Triticum aestivum L.
ZHANG Lei1,YU Yongang1,2,YANG Tianyou1
(1.School of Life Science and Technology,Henan Institute of Science and Technology,Collaborative Innovation Center of Modern Biological Breeding of Henan Province,Xinxiang 453003,China;2.College of Agronomy,Northwest A&F University,Yangling 712100,China)
To investigate the function of TaGST gene,RT-PCR was used to obtain TaGST gene open reading frame sequence from wheat and analyzed by bioinformatics method.The sequence analysis results showed that the ORF of TaGST gene had a length of 690 bp coding for 229 amino acid,and the relative molecular weight of TaGST protein was 25.81 kDa and its theoretical isoelectric pointwas 5.29.Homology analysis showed that the amino acid sequence of TaGST was highly homologous with Oryza sativa,and phylogenetic analysis of the relationship of the newly identified TaGST with some known plant GSTs grouped the TaGST into the class of PhiGSTs.The prokaryotic expression of TaGST gene was done after construction of its prokaryotic expression vector pET32-TaGST,and the SDS-PAGE results displayed that the expressed protein was consistentwith the anticipated size.The resultswere expected to lay a foundation for further studies on the properities and function of this gene.
Wheat;Glutathione-S-transferase;Gene clone;Porkaryotic expression
Q78;S512.03 文献标识码:A 文章编号:1000-7091(2016)03-0038-06
10.7668/hbnxb.2016.03.006
2015-11-26
国家自然科学基金项目(1305047);河南省教育厅科学技术研究重点项目(12A180011);河南省高等学校青年骨干教师资助计划项目(2014GGJS-100)
张 蕾(1987-),女,河南新乡人,助理实验师,硕士,主要从事植物分子生物学研究。
猜你喜欢
中国医学影像学杂志(2021年6期)2021-08-13
国际放射医学核医学杂志(2020年4期)2020-07-27
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
水产科学(2016年1期)2016-12-19
天然产物研究与开发(2016年6期)2016-06-05
中国继续医学教育(2015年3期)2016-01-06
安徽医科大学学报(2015年9期)2015-12-16
动物医学进展(2015年10期)2015-12-07
动物医学进展(2015年10期)2015-12-07
