
野扁桃的繁育系统研究
2017-03-23 01:41罗淑萍
西北农业学报 2017年2期
程 靖,李 疆,罗淑萍,陆 婷
(新疆农业大学 林学与园艺学院,乌鲁木齐 830052)
野扁桃的繁育系统研究
程 靖,李 疆,罗淑萍,陆 婷
(新疆农业大学 林学与园艺学院,乌鲁木齐 830052)
为研究孑遗植物野扁桃的繁育系统,采用野外观测、荧光显微观察及统计分析方法对野扁桃的繁育系统进行研究。结果表明,该种单花开花持续时间为8 d,花冠直径为(24.79 ± 0.40) mm;花粉活力在刚散粉时最高达96.88%,在第17天半数花粉失去活性,第28天花粉完全失去活性;柱头在开花第1~3天可授性最强,可持续7 d;杂交指数为3;自花花粉和异花花粉均能在柱头上萌发,授异花花粉59 h后,花粉管进入子房。人工控制授粉试验表明,自花、异花授粉均能结实,需要昆虫传粉媒介。以上结果说明,野扁桃属于混合交配繁育系统类型,需要传粉者。
野扁桃;杂交指数;授粉试验;繁育系统
野扁桃(AmygdalusledebourianaSchlechet)也称矮扁桃或野生巴旦杏,属蔷薇科李亚科桃属扁桃亚属植物,原产于欧洲东南和亚洲中西部,是珍贵的第三纪孑遗植物种类[1],目前仅分布于哈萨克斯坦和新疆[2]。野扁桃不仅是多年生速生、抗旱、耐寒的油料兼药用经济树种[3],并且其花色艳丽、芳香,可作为早春观花灌木在园林中应用,然而近年来,野扁桃自然居群分布面积在逐渐缩小[4],已被列入国家重点保护野生植物名录[5],深入了解野扁桃居群退化的原因是保护和开发利用野扁桃种质资源的必要途径。
植物繁育系统的生殖力、存活力、适应力低下等内在因素是植物走向濒危的根本原因[6],所以,要找出野扁桃居群的退化原因,了解其繁育系统是必要的途径。植物的繁育系统是探讨物种多样性发生历史、维持机理和保护策略的综合研究,是所有影响后代遗传组成有性特征的总和,主要包括花形态特征、传粉特性和交配系统等[7],曾斌[8]对新疆野扁桃繁殖生物学中的花芽形态分化、实生繁殖及组织培养繁殖进行研究,并进行人工授粉试验,报道授粉15 d后的幼果坐果率,而对野扁桃成熟果坐果率高低、是否需要传粉媒介以及限制结实的因素等还有待于进一步研究。为此,本试验对野扁桃的繁育系统进行研究,以期为深入了解野扁桃的结实特性、揭示其种群生态适应性机制和促进种群恢复提供科学依据。
1 材料与方法
1.1 试验材料与试验地概况
试验材料采自新疆塔城地区裕民县境内的野扁桃自然保护区,该地区海拔900~1 200 m,地理位置为北纬45°51′~45°55′,东经82°26′~82°33′,气温较高,年平均温度5.3 ℃,最冷月为1月,平均气温-6.7 ℃;最热月为7月,平均气温19 ℃,极端最高气温37.4 ℃,极端最低气温-32.0 ℃;降水丰富,土质松软肥沃,土壤多为亚高山腐质土、草甸土、钙质土,土层较厚,主要是残积风化物、堆积物、洪积物等构成的母质,有机质含量丰富[8]。
1.2 研究方法
1.2.1 花部特征 开花前随机标记30个生长一致的花蕾,观测记录花部形态、单花花期、花药散粉所用的时间、雌雄蕊的空间位置,并测量花冠直径。1.2.2 花粉活力与柱头可授性的检测 参考郭红超等[9]的方法,采用MTT(噻唑蓝)染色法对野扁桃的花粉活力及柱头可授性进行测定。花粉活力测定具体方法:将即将开放的花蕾取回置于室内通风处,散粉后用棉签将花粉弹于载玻片上,将1滴MTT溶液(100 mg MTT溶解在5 mL质量分数为5%的蔗糖溶液中)置于花粉样品中,充分混匀,风干后再重复滴加1次MTT溶液,干后在显微镜下观察,若花粉变蓝黑色则表明有活力,若无变化或黄褐色则表明为无活力,统计着色花粉粒和未着色的花粉粒数目,然后计算花粉的活力百分率,每天定时观察1次,每次制片3张,每片计数3个视野。
柱头可授性测定的具体方法为:从10个植株上选取发育一致,即将开花的花蕾套袋以防止昆虫拜访,并做标记,从开花之日到花柱萎蔫为止,每天12:00随机摘取10个标记花朵,将其柱头浸入MTT溶液10 min,若柱头颜色变为深蓝色则表明其具有可授性,染色越深表示其柱头表面酶活性越高,可授性也越强,无颜色变化则不具可授性。1.2.3 杂交指数(Outcrossing index, OCI) 依照Dafini[10]的方法计算OCI值,具体方法为:(1)花朵或花序直径 <1 mm记为0,1~2 mm记为1,2~6 mm记为2,>6 mm记为3。(2)花药开裂时间与柱头可授粉期之间的时间间隔,同时或雌蕊先熟记为0,雄蕊先熟记为1。(3)柱头与花药的空间位置,同一高度记为0,空间分离记为1。三者之和为OCI值。当OCI=0时,繁育系统为闭花受精(Cleistogamy);OCI=1时,繁育系统为专性自交(Obligate autogamy);OCI=2时,繁育系统为兼性自交(Facultative autogamy);OCI=3时,繁育系统为自交亲和,有时需要传粉者,OCI=4 时,繁育系统为部分自交亲和,异交,需要传粉者。
1.2.4 野扁桃人工授粉的荧光显微观察 开花前标记即将同日开放的300个花蕾,去雄套袋,分成2组,一组开花后人工授自花花粉,另一组授异花花粉,授粉后每隔3 h(22:00至次日10:00停止采样)随机采15朵花,将雌蕊用FAA(由体积分数70%的酒精、甲醛、冰醋酸按照体积比为89∶6∶5的方法配制)固定并带回实验室,用苯胺蓝溶液(质量分数0.1%的苯胺蓝+质量分数2.6 %的磷酸氢二钠)染色并整体压片,Olympus BH 22RFL型荧光显微镜下观察并拍照。
1.2.5 控制授粉试验 2015年4月在观测点随机标记植株大小相对一致且未开放的花近2 000朵,并将其分成5组进行如下处理:(1)对照,不套袋、不去雄、自由传粉,用于检测自然条件下的传粉情况;(2)开花前去雄,套纸袋,检测野扁桃是否无融合生殖;(3)开花前套纸袋、不去雄,检测是否需要传粉者;(4)人工自花传粉,不去雄、套袋,检测自交是否亲和;(5)人工异株异花传粉,套纸袋、去雄,检测异交结实情况。
2 结果与分析
2.1 花部特征
野扁桃花两性,辐射对称,有花蜜和香气,单花花期8 d;花瓣近圆形,粉红色,花冠直径(24.79±0.40)mm;雄蕊为20~31枚,平均24枚,花丝下部与萼筒贴生,花朵初开时为白色,开花后第4天基部渐变为玫红色;花药黄色,花瓣打开后花药从外轮向内轮依次开裂散粉,全部花药开裂历时2 d;雌蕊子房上位,1室,着生2枚胚珠,花柱细长,柱头绿色,为湿型柱头,与雄蕊等高。
2.2 花粉活力与柱头可授性的检测
从图1看出,野扁桃花药散粉第1 天花粉活力最强,达到96.88%,此后逐渐降低,第17天半数花粉失去活力,直到第28天花粉活力完全丧失。柱头可授性检测结果表明,花初开时柱头就能被染成深蓝色,根据柱头染色的速度及颜色,发现第1~3天柱头染色最快、最深,表明柱头表面酶有较高活性,可授性最强。随后可授性下降,到第7天时柱头虽仍能被染为蓝色,但柱头表面已明显萎蔫褐变,不具备接受花粉的能力。
2.3 杂交指数
杂交指数(Outcrossing index, OCI)常用以判断显花植物的繁育系统类型,OCI值越高,繁育系统越倾向于异交并依赖传粉媒介,相反则倾向于自交。根据野外观测,野扁桃花直径>6 mm,记为3;花粉活力和柱头可授性检测的结果表明花粉活力较强时,柱头也具有可授性,同时成熟,记为0;雌雄蕊等高,记为0,故其杂交指数值为3,根据Dafini[10]杂交指数的判定标准,野扁桃的繁育系统为自交亲和,有时需要传粉者。
2.4 野扁桃人工授粉的荧光显微观察
通过荧光显微观察发现,自花花粉和异花花粉均能在柱头萌发,图2-a、2-b显示授粉7 h后自花花粉和异花花粉在柱头上萌发的情况;从图2-c上可以看到,授自花花粉32 h后,花粉管到达花柱的近1/2处,未观察到花柱中的胼胝质塞;从图2-d可以看出,授异花花粉59 h后,花粉管进入子房。
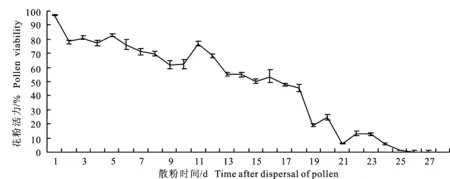
图1 野扁桃的花粉活力变化曲线Fig.1 Survivorship curve of pollens viability in A.ledebouriana
2.5 控制授粉对野扁桃坐果率的影响
从表1可以看出,去雄套袋处理没有结实,表明野扁桃没有无融合生殖现象;套纸袋防止昆虫拜访的处理也没有结实,表明其没有自发的自花传粉现象,其结实需要传粉媒介;人工辅助授粉处理的结果率均高于自然条件结果率,说明可能存在传粉者不足的结实限制性因素;异花授粉高于自花授粉的结实率表明其繁育系统倾向于异交。
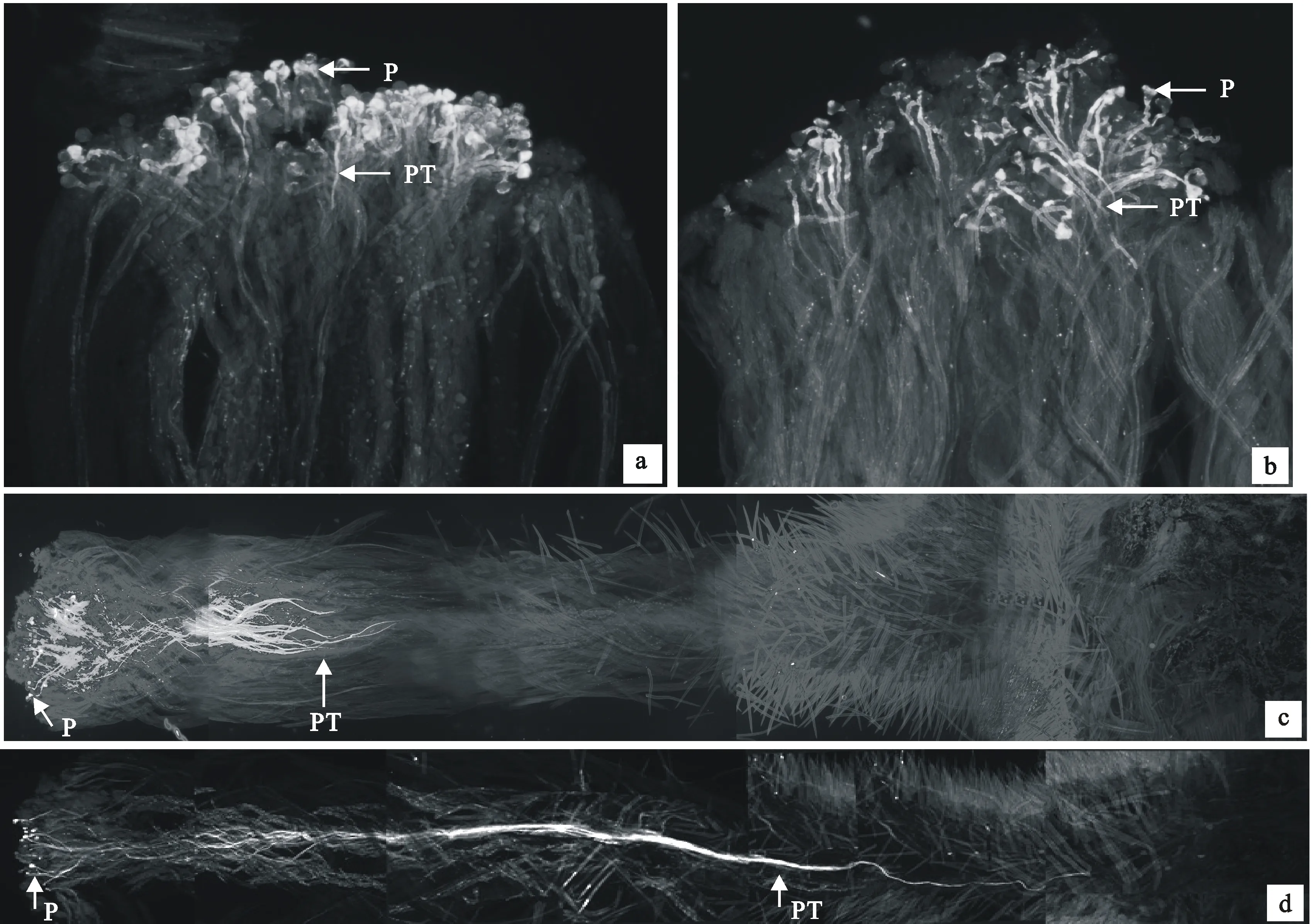
P.花粉粒 Pollen; PT.花粉管 Pollen tube;a.花粉在柱头萌发(自花花粉) Pollen’s germination on the stigma (self combination); b.花粉在柱头萌发(异花花粉) Pollen’s germination on the stigma (cross combination); c.32 h花粉管伸长到花柱的接近1/2处(自花花粉) Pollen tubes grow nearly to 1/2 of style (self combination); d.花粉管生长至子房(异花花粉) Pollen tube growth to ovary (cross combination)

图2 野扁桃授粉的荧光显微观察Fig.2 Study on pollination experiment and growth of pollen tube under fluorescent microscope on A.ledebouriana
3 讨 论
自交和异交是植物繁育系统研究的一对中心问题[11]。在自然界中,纯粹的自交和异交种是很少见的,大多数植物都属于混合交配系统[12-14],虽然异交可以维持种群内高水平的遗传变异,但自交率高的植物其花药和柱头的空间距离近,导致花粉更容易落在自己的柱头上,而外来粉量却几乎不受影响[15],因而能克服传粉媒介的短缺,实现繁殖保障。与栽培扁桃和蒙古扁桃自交不亲和的繁育系统不同[16-17],野扁桃的杂交指数OCI值、自花粉和异花粉受精过程荧光显微观察以及田间授粉试验等结果均表明新疆野扁桃的繁育系统为自交亲和,传粉过程需要传粉者,属于混合交配系统,与曾斌[8]的结论相符。植物的繁育系统与环境因子存在着密切的联系[18],面临恶劣的环境条件,植物的繁育系统一般具有多样性,且多为混合交配方式[19],野扁桃属于早春开花植物,常伴随着频繁的雨雪、多风天气,因此,野扁桃在繁育系统上所表现出的特点,可以有效保障其在不利条件影响下的成功繁殖,也是对恶劣环境的生态适应性的表现。
Burd[20]提出在被子植物中花粉限制是一个普遍现象,研究结果显示野扁桃上也存在花粉限制,首先,野扁桃花粉和柱头同时呈现,这暗示着野扁桃受到传粉者限制,因为两性花植物雌雄功能在时间上和空间上分离能阻止花内自花传粉及雌雄功能干扰[21],而在缺少传粉者的情况下,花粉和柱头同时呈现则有利于在昆虫的一次拜访中两性功能同时受益;此外,人工授粉能提高结实率的结果也表明其受到花粉限制。近年来野扁桃保护区虫害严重[22],喷洒药物防治害虫的同时也可能会对传粉昆虫造成威胁,导致传粉昆虫数量不足,进而影响到野扁桃等依赖于传粉昆虫的野生植物的生殖成功,因此在防治害虫时还应重视传粉昆虫的保护。
野扁桃成熟果实的自然结实率很低,仅有2.64%,即使人工异花授粉,野扁桃结实率也才达到4.09%,相对于69%的幼果结实率较低[8],说明果实在成熟过程中落果严重。除了花粉限制以外,资源限制也是导致花多果少的重要因素,而且花粉和资源限制往往同时发生[23],那么野扁桃是否受到资源限制从而导致大量落果?其他环境因素,如晚霜冻、大风、越来越严重的虫害[22],以及放牧是否导致幼果脱落,并在多大程度上影响野扁桃生殖成功?这些问题还有待于进一步研究,这样才能全面了解野扁桃的濒危机制。建议提高自然保护区野扁桃病虫害综合防治水平,注重传粉昆虫的保护,并保护其生境不受破坏,以有效地保护野扁桃这一珍贵物种资源。
Reference:
[1] 朱京琳.新疆巴旦杏[M].乌鲁木齐:新疆人民出版社,1983:137-155.
ZHU J L.Xinjiang Almond [M].Urumqi:Xinjiang People’s Publishing House,1983:137-155 (in Chinese).
[2] 曹青爽,姬玉英.新疆特有植物--野扁桃资源价值及保护利用[J].中国野生植物资源,2007(4):41-42,46.
CAO Q SH,JI Y Y.Values and protection of wide almonds resources grown in Xinjiang [J].ChineseWildPlantResources,2007(4):41-42,46 (in Chinese with English abstract).
[3] 吾买尔夏提·塔汉,艾尔买克·南蓬.新疆野扁桃种仁油脂肪酸成分的GC-MS分析[J].中国油脂,2009(11):77-79.
Wumaierxiati·Tahan,Aiermaike·Nanpeng.Fatty acid composition analysis ofAmygdalusledebourianaSchlecht seed oil with GC-MS [J].ChinaOilsandFats,2009(11):77-79 (in Chinese with English abstract).
[4] 李 疆,曾 斌,罗淑萍,等.我国野扁桃资源的保护及引种繁育[J].新疆农业科学,2006,43(1):61-62.
LI J,ZENG B,LUO SH P,etal.Protection and propagation ofAmygdalusledebourianaSchleche in China [J].XinjiangAgriculturalSciences,2006,43(1):61-62 (in Chinese with English abstract).
[5] 伊林克.新疆珍稀濒危特有高等植物[M].乌鲁木齐:新疆科学技术出版社,2006:9.
YI L K.Rare Endangered Endemic Higher Plants in Xinjiang of China [M].Urumqi:Xinjiang Science and Technology Press,2006:9(in Chinese).
[6] 张文辉,祖元刚,刘国彬.十种濒危植物的种群生态学特征及致危因素分析[J].生态学报,2002,22(9):1512-1520.
ZHANG W H,ZU Y G,LIU G B.Population ecological characteristics and analysis on endangered cause of ten endangered plant species [J].ActaEcologicaSinica,2002,22(9):1512-1520 (in Chinese with English abstract).
[7] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
HE Y P,LIU J Q.A review on recent advances in the studies of plant breeding system [J].ActaPhytoecologicaSinica,2003,27(2):151-163(in Chinese with English abstract).
[8] 曾 斌.新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究[D].乌鲁木齐:新疆农业大学,2008.
ZENG B.A study on propagate biological characteristics and genetic diversity of germplasm resources ofAmygdalusledebourianaSchlecht [D].Urumqi:Xinjing Agricultural University,2008 (in Chinese with English abstract).
[9] 郭红超,严 成,魏 岩.木地肤的开花动态与花粉活力及柱头可授性研究[J].草业学报,2014,23(4):87-93.
GUO H CH,YAN CH,WEI Y.Study on the flowering dynamic,pollen viability and stigma receptivity ofKochiaprostrate[J].ActaPrataculturaeSinica,2014,23(4):87-93 (in Chinese with English abstract).
[10] DAFINI A.Pollination Ecology:A practical Approach [M].New York:Oxford Univercity Press,1992:1-57.
[11] WYATT R.Pollinator Plant Interactions and the Evolution of Breeding Systems[M]//REAL L.Pollination Biology.New York:Academy Press,1983:51-95.
[12] LLOYD D G.Some reproductive factors affecting the selection of self-fertilization in plants [J].AmericanNaturalist,1979,113(1):67-79.
[13] BARRETT S C H,ECKERT C G.Variation and Evolution of Plant Mating Systems[M]//KAWANO S.Biological Approaches and Evolutionary Trends in Plants.New York:Academic Press,1990:229-254.
[14] VOGLER D W,KALISZ S.Sex among the flowers:the distribution of plant mating systems [J].Evolution,2001,55(1):202-204.
[15] 张大勇,姜新华.植物交配系统的进化、资源分配对策与遗传多样性[J].植物生态学报,2001,25(2):130-143.
ZHANG D Y,JIANG X H.Mating system evolution,resource allocation,and genetic diversity in plants [J].ActaPhytoecologicaSinica,2001,25(2):130-143 (in Chinese with English abstract).
[16] 高启明.新疆栽培扁桃自交不亲和性的研究[D].乌鲁木齐:新疆农业大学,2011.
GAO Q M.Study of self-incompatibility from almond in Xinjiang [D].Urumqi:Xinjing Agricultural University,2011 (in Chinese with English abstract).
[17] 马 骥,倪细炉,史宏勇,等.蒙古扁桃的开花生物学研究[J].西北植物学报,2010,30(6):1134-1141.
MA J,NI X L,SHI H Y,etal.Flowering biology ofAmygdalusmongolica[J].ActaBotanicaBoreali-occidentaliaSinica,2010,30(6):1134-1141 (in Chinese with English abstract).
[18] 方炎明.森林植被的显花、传粉与繁育系统[J].南京林业大学学报(自然科学版),2012,36(6):1-7.
FANG Y M.Florescence,pollination and breeding systems of forest vegetation [J].JournalofNanjingForestryUniversity(NaturalScienceEdition),2012,36(6):1-7 (in Chinese with English abstract).
[19] 何亚平,费世民,刘建全,等.高山植物繁育系统研究进展初探[J].四川林业科技,2005,26(4):43-49.
HE Y P,FEI SH M,LIU J Q,etal.A preliminary review of studies of alpine plant breeding system [J].JournalofSichuanForestryScienceandTechnology,2005,26(4):43-49 (in Chinese with English abstract).
[20] BURD M.Ovule packaging in stochastic pollination and fertilization environments [J].Evolution,1995,49(1):100-109.
[21] 白伟宁,张大勇.雌雄同体植物的性别干扰及其进化意义[J].植物生态学报,2005,29(4):672-679.
BAI W N,ZHANG D Y.Sexual interference in cosexual plants and its evolutionary implications [J].ActaPhytoecologicaSinica,2005,29(4):672-679 (in Chinese with English abstract).
[22] 孙谊祥.野巴旦自然保护区病虫害调查及综合治理技术简报[J].新疆林业,2001(2):43.
SUN Y X.A briefing on pest survey and comprehensive management technology ofAmygdalusledebourianaschleche in nature reserve [J].XinjiangForestry,2001(2):43(in Chinese).
[23] CAMPBELL D R,HALAMA K J.Resource and pollen limitations to lifetime seed production in a natural plant population [J].Ecology,1993,74(4):1043-1051.
(责任编辑:潘学燕 Responsible editor:PAN Xueyan)
Study on Breeding System ofAmygdalusledebouriana
CHENG Jing, LI Jiang, LUO Shuping and LU Ting
(College of Forestry and Horticulture, Xinjiang Agricultural University, Urumqi 830052, China)
A.ledebourianais a relict species in Xinjiang.Its breeding system was studied by field observation, fluorescence microscopy observation and statistical analysis methods.The results showed that the flowering process of single flower lasted for 8 days and the corolla diameter was (24.79±0.40) mm;the pollen viability was 96.88% at beginning, half pollen lost activity after 17 days and all pollens lost activity in the 28 th day;stigma had highest receptivity from 1 to 3 days at blossoming stage, lasting 7 days;the outcrossing index was 3;pollens in both self-pollination and cross-pollination germinated on the stigma, and pollen tubes could reach the ovary after 59 hours in cross-pollination.The study of artificial pollination showed that both self-pollination and cross-pollination were fertile, which it needed insects to be the pollination media.The results suggested that the breeding system is self-compatible and pollinators is needed.
Amygdalusledebouriana; Hybrid index; Artificial pollination; Breeding system
CHENG Jing, female, master student.Research area:ornamental plants and horticulture.E-mail:893116635@qq.com
LU Ting,female,Ph.D, associate professor, master supervisor.Research area:plant reproductive biology.E-mail:luting0909@126.com
日期:2016-12-29
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20161229.1008.024.html
2016-01-29
2016-04-10
国家自然科学基金(31260186)。
程 靖,女,硕士研究生,研究方向为园林植物与观赏园艺。E-mail:893116635@qq.com
陆 婷,女,博士,副教授,硕士生导师,研究方向为植物繁殖生态学。E-mail:luting0909@126.com
S662.9
A
1004-1389(2017)02-0242-06
Received 2016-01-29 Returned 2016-04-10
Foundation item National Natural Sciecne Foundation of China(No.31260186).
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
铜仁学院学报(2018年7期)2018-09-08
中成药(2018年6期)2018-07-11
现代园艺(2017年22期)2018-01-19
中国稻米(2017年6期)2017-12-28
甘肃林业(2016年3期)2016-11-07
国外医药(抗生素分册)(2016年2期)2016-07-12
作物研究(2015年4期)2015-03-24
