
利用功能标记揭示新疆小麦改良品种与地方品种的遗传变异
2018-05-14 08:07简大为刘宏伟买春艳于立强韩新年张宏军李洪杰
作物学报 2018年5期
简大为 周 阳 刘宏伟 杨 丽 买春艳 于立强 韩新年 张宏军,* 李洪杰,*
利用功能标记揭示新疆小麦改良品种与地方品种的遗传变异
简大为1,2周 阳2刘宏伟2杨 丽2买春艳3于立强4韩新年5张宏军2,*李洪杰2,*
1新疆生产建设兵团第四师农业科学研究所, 新疆伊宁 835000;2中国农业科学院作物科学研究所 / 农作物基因资源与基因改良国家重大科学工程, 北京 100081;3新乡矮败小麦育种技术创新中心, 河南新乡 453731;4石家庄市农林科学研究院赵县试验基地, 河北赵县 051530;5谷物品质与遗传改良兵团重点实验室, 新疆石河子 832000
揭示新疆小麦改良品种与地方品种在主要农艺性状相关基因上的遗传变异对进一步改良和利用新疆育成品种具有重要意义。本研究利用52个功能标记对136份新疆小麦改良品种和地方品种分析发现, 与适应性相关的矮秆等位变异和、半冬性生长习性相关等位变异、T1BL·1RS易位系, 与品质相关的高脂肪氧化酶活性等位变异、低多酚氧化酶活性等位变异、低黄色素含量等位变异以及与高粒重等位变异()仅分布在改良品种中, 而且光周期不敏感等位变异(77.6%)、优质麦谷蛋白亚基Dx5+Dy10 (35.4%)和硬质等位变异(25.0%), 以及高千粒重等位变异(63.3%)、() (33.8%)、() (93.7%)、() (77.9%)、(78.5%)、(50.0%)和(92.1%)在改良品种中分布频率明显高于地方品种。大部分优异等位变异分布频率随着育种时期的推进呈现不连续性上升趋势。在适应性与品质相关基因方面, 春性改良品种的优异等位变异频率高于冬性改良品种。功能标记分析显示改良品种的遗传多样性高于地方品种。136份新疆小麦资源被聚为改良品种和地方品种两类, 改良品种被进一步聚为冬性和春性两类, 说明新疆改良品种与地方品种间存在明显的遗传差异。本研究鉴定的优异等位基因和等位基因组合为进一步改良新疆小麦品种提供了重要信息。
小麦; 新疆; 功能标记; 农艺性状; 遗传变异
小麦是新疆的主要粮食作物, 约占全省主要粮食作物种植面积的40%~60%[1]。经过几十年的努力, 新疆小麦的产量和品质在近七、八十年取得了突破性进展, 单产水平由1949年的870 kg hm–2 [2]提高到2016年的6488 kg hm–2(http://www.feedtrade.com.cn/ yumi/yumi_china), 目前主推小麦品种大部分达到中筋水平[3]。品种推广也经历了几轮更换, 从20世纪50年代以地方品种为主[2], 到21世纪一大批优质、高产、广适性改良品种的推广, 小麦优良品种在不同时期对新疆小麦生产都发挥了重要的作用。育种是一个优异基因不断聚合和选择的过程, 这也是导致优异基因频率不断提高的原因[4]。随着育种和生产的发展, 新疆地方品种逐渐被改良品种取代, 这种品种演替是否与改良品种在主要农艺性状相关的基因上所经历的强选择压有关, 有待研究。因此, 有必要全面了解新疆改良品种与地方品种间在与适应性、品质和产量等性状相关基因上的遗传变异。
功能标记(functional marker)是依据基因序列的多态性开发的, 这些基因的不同等位变异与表型直接相关[5]。截至2012年, 已开发了96个小麦功能标记, 涉及30多个基因座93个等位变异[6]。抽穗期是小麦的重要农艺性状之一, 对小麦适应不同生态环境具有重要的作用。它主要受春化基因(vernalization,)、光周期基因(photoperiod,)和早熟性本身基因(earliness,)[7]控制, 其中,、、、、、和是决定春化和光周期反应的主要基因座[8-11]。“绿色革命”基因“”主要包括和, 其中, 矮秆等位变异和被广泛用于降低小麦株高, 提高抗倒伏能力和产量[12]。小麦-黑麦T1BL·1RS易位系广泛用于产量、适应性、抗病性和其他农艺性状改良, 在小麦生产上发挥了重要作用[5]。小麦的加工品质主要与高、低分子量麦谷蛋白亚基(high- and low-molecular weight glutenin subunits)、脂肪氧化酶(lipoxygenase, LOX)活性、籽粒硬度(puroindoline b,)基因、多酚氧化酶(polyphenol oxidase, PPO)活性和黄色素含量相关基因八氢番茄红素合酶(phytoene synthase, PSY)密切相关[13-19]。粒重是小麦籽粒产量重要构成因素之一, 由多基因控制, 细胞壁转化酶(cell wall invertase,)基因、谷氨酰胺合酶(glutamine synthetase,)基因、籽粒大小(grain size,)基因、蔗糖合酶(sucrose synthase,)基因和粒重相关(IAA-glucose hydrolase gene,)基因是控制粒重和籽粒大小的重要基因, 目前已经从小麦上克隆了这些基因的同源基因, 而且已有相应的功能标记[20-26]。
本研究比较了与适应性、品质和粒重相关基因等位变异在新疆改良品种和地方品种中的频率变化, 揭示了相关基因优异等位变异在不同育种时期频率变化趋势, 并且进行了改良品种和地方品种的遗传多样性和群体结构分析。研究结果不仅可以解释改良品种取代地方品种的原因, 而且可以为育种者提供理想的育种材料和聚合优异等位基因提供重要的参考信息。
1 材料与方法
1.1 供试品种
包括新疆小麦资源136份, 其中79份改良品种(附表1), 57份地方品种(附表2)。前者包括35份春性品种和44份冬性品种, 都是1966年至2015年育成或审定的新疆主推品种。后者包括50份春性地方品种和7份冬性地方品种。
1.2 分子标记检测
取每份供试材料10粒种子放在带有湿润试纸的培养皿中发芽, 室温下生长5 d, 然后从每份材料各个植株上取等量叶片混合用于DNA提取[27]。
利用52个与适应性、品质和粒重相关的功能标记对供试材料进行等位变异分析。适应性相关的基因包括(和)[12]、(、、和)[8-9,28]、(、和)[10-11]和T1BL·1RS易位系()[29]; 与品质相关的基因包括(Dx5+Dy10)[13]、()[14]、()[15]、()[17]和(和)[18-19]; 粒重相关基因主要包括(、和)[20-21]、()[22]、()[23]和()[24]、()[25]和()[26]。
PCR在Biometra Thermocycler (Germany)上进行, 总反应体系为20 μL, 包括2 ×PCR Master Mix 10 μL, 正反引物(10 μmol L–1)各1 μL, DNA模板(50~100 ng μL–1) 2 μL。PCR程序为94°C预变性5 min, 94°C变性30 s, 57~60°C退火30 s, 72°C延伸1~3 min, 72°C后延伸10 min。PCR扩增产物经2%琼脂糖凝胶电泳检测, 溴化乙锭染色后用凝胶成像系统观察电泳结果, 读取基因型。需要说明的是对于粒重相关基因座、、、和在PCR扩增的基础上, 分别利用限制性内切酶I、Y I、R I、I和II进行酶切, 用2%琼脂糖凝胶电泳检测酶切产物。
1.3 统计分析
采用PowerMarker v3.25软件[30]分析每个基因座等位基因数目、遗传多样性指数和多态性信息含量(PIC)。利用Arlequin v3.5软件[31]对亚组间(改良品种和地方品种)、亚组内个体间以及个体间进行分子方差分析(analysis of molecular variance, AMOVA)。通过NTSYS-pc v2.1软件[32]对供试材料进行基于主成分(PCA)的群体结构分析。
为了比较不同时期新疆小麦品种基因频率变化趋势, 按照品种的育成或审定时间将136个小麦资源分成6组, 即地方品种(57份)、1966—1990年(13份)、1991—2000年(18份)、2001—2005年(19份)、2006—2010年(17份)和2011—2015年(12份); 冬性品种和春性品种也分别按上述原则分为6组, 冬性品种各组依次为7、8、11、6、3和7份, 春性品种各组依次为50、5、7、13、14和5份。通过简单线性回归方程=+分析优异等位变异频率分别在所有供试品种、冬性和春性品种不同时期的变化趋势, 其中,表示因变量,表示自变量,表示回归截距,表示回归系数。用回归系数的大小来反映变化趋势, 具体参考Meng等[4]的方法进行回归分析。
2 结果与分析
2.1 适应性、品质和粒重性状基因不同等位变异分布频率
利用52个功能标记对136份新疆小麦资源进行等位变异分型, 部分引物扩增结果见图1。从这些基因不同等位变异分布频率看, 无论是冬性或春性资源, 与适应性相关的矮秆等位变异和、光周期不敏感等位变异以及T1BL·1RS易位系在改良品种中的分布频率均高于地方品种(表1)。与冬性生长习性密切相关的隐性等位变异、和主要分布在改良品种中; 与春性生长习性相关的显性等位变异在改良品种中分布频率也明显高于地方品种。矮秆等位基因组合(5.1%)仅在改良品种中出现且在春性改良品种中分布频率更高。相反, 高秆等位变异组合主要分布在地方品种中, 频率高达98.2%, 且这一等位变异组合在冬性地方品种频率高于春性地方品种(图2-A)。春性改良品种的和等位变异以及T1BL·1RS易位系的分布频率高于冬性改良品种。相反, 冬性改良品种等位变异分布频率高于春性改良品种。
与品质相关的优质麦谷蛋白亚基Dx5+Dy10、面粉色泽相关的高脂肪氧化酶活性等位变异、硬质等位变异、低多酚氧化酶活性等位变异和低黄色素含量等位变异在全部、冬性和春性改良品种中的分布频率均高于相应的地方品种(表1), 而且只在改良品种中检测到优异等位变异组合+、+、+和Dx5+Dy10。冬性改良品种和的频率高于春性改良品种; 相反, 春性改良品种Dx5+Dy10、和等位变异的频率高于冬性改良品种, 而且只在春性改良品种中检测到和等位变异。值得注意的是, 除了Dx5+Dy10外, 其他3个优异等位变异组合仅分布于冬性改良品种(图2-B)。

图1 部分适应性(A)、品质(B)和粒重(C)性状基因在12个新疆小麦品种中的扩增结果
1: 新春3号; 2: 新春4号; 3: 新春5号; 4: 新春6号; 5: 新春7号; 6: 新春8号; 7: 新春9号; 8: 新春10号; 9: 新春11; 10: 新春12; 11: 新春13; 12: 新春14。
1: Xinchun 3; 2: Xinchun 4; 3: Xinchun 5; 4: Xinchun 6; 5: Xinchun 7; 6: Xinchun 8; 7: Xinchun 9; 8: Xinchun 10; 9: Xinchun 11; 10: Xinchun 12; 11: Xinchun 13; 12: Xinchun 14.

表1 新疆改良品种和地方品种与适应性、品质和粒重性状相关基因等位变异分布频率比较

(续表1)
在粒重相关的8个基因座中, 高粒重等位变异、()、()、()、、、()和在改良品种中的分布频率均高于地方品种(表1)。在14对高粒重等位变异组合中,+、+、+和+在改良品种中频率明显高于地方品种, 分别达到82.3%、72.2%、63.3%和70.9% (图2-C)。冬性改良品种高粒重等位变异组合多于春性改良品种。
2.2 适应性、品质和粒重性状基因优异等位变异在不同育种时期的频率变化
在与适应性、品质和粒重性状相关的17个优异等位变异中, 有11个分布频率随育种时期推进呈现不连续性上升的趋势(图3)。矮秆等位变异在地方品种1991—2000年期间呈现连续上升趋势, 2000年频率达到94.1%; 2001—2005年期间频率下降到73.9%, 2005年以后保持平稳(图3-A)。在冬性改良品种中,分布频率在2001—2010年期间呈现下降趋势, 2011年后又出现上升趋势; 而春性改良品种变化趋势与所有品种的变化趋势一致。除了在2006—2010期间, 等位变异分布频率随着育种时期推进呈明显上升趋势(图3-B)。T1BL·1RS易位系仅分布在改良品种中, 在地方品种1991—2000年期间无论是冬性或春性改良品种, 其易位系分布频率均随育种时期推进呈现上升趋势, 2000年以后呈现下降趋势(图3-C)。与面粉色泽相关的高脂肪氧化酶活性等位变异分布频率随育种时期推进显著提高, 特别是冬性改良品种呈直线上升趋势, 回归系数达到15.0 (< 0.01), 且2达到0.96 (< 0.01)(图3-D)。优质麦谷蛋白亚基Dx5+Dy10分布频率随育种时期推进呈现不连续的上升趋势, 但在2011年以后的改良品种中出现下降趋势(图3-E)。高千粒重等位变异(图3-F)、() (图3-G)、(图3-H)、(图3-I)、(图3-J)和(图3-K)分布频率在地方品种–2000年期间均呈现直线上升趋势, 随着育种时期的推进在2001年以后分布频率呈现不连续上升, 其中, 等位变异的分布频率在2006年以后的改良品种中达到100%。随着育种时期推进, 6个等位变异在冬、春性改良品种中的分布频率呈现不同的变化趋势, 特别是等位变异在春性改良品种中没有出现, 而在冬性改良品种中于地方品种–2010年期间呈现直线上升趋势(图3-G)。

图2 影响株高(A)、品质(B)和粒重(C)基因优异等位变异组合在新疆改良品种和地方品种中分布频率

图3 适应性、品质和粒重基因优异等位变异在不同育种时期频率变化
A:等位变异; B:等位变异; C: TlBL·1RS易位系; D:等位变异; E: Dx5+Dy10亚基; F:等位变异; G:(等位变异); H:等位变异; I:等位变异; J:等位变异; K:等位变异。*和**分别表示2在0.05和0.01概率水平显著。
A:allele; B:allele; C: TlBL·1RS translocation line; D:allele; E: Dx5+Dy10 subunits; F:allele; G:(allele); H:allele; I:allele; J:allele; K:allele.*and**indicate significance of2at the 0.05 and 0.01 probability levels, respectively.
2.3 改良品种和地方品种的遗传多样性比较
除了春性资源与粒重相关基因座的多态性信息含量(PIC)平均值外, 无论是全部、冬性还是春性资源, 改良品种所有基因座的平均等位基因数目和平均PIC均高于地方品种(表2)。春性改良品种在与适应性相关以及品质相关基因座的平均等位基因数目和平均PIC高于冬性改良品种。相反, 冬性改良品种与粒重相关基因座的平均PIC高于春性改良品种。在适应性相关基因的10个基因座中, 春性改良品种的平均等位基因数目(1.70)和PIC (0.20)均高于冬性改良品种(1.50和0.10)。在品质相关基因的8个基因座中, 春性改良品种平均等位基因数目(2.00)和平均PIC (0.23)亦高于冬性改良品种(1.63和0.21)。在粒重相关基因的8个基因座中, 春性和冬性改良品种的平均等位基因数目相等, 而春性改良品种的平均PIC高于冬性改良品种。
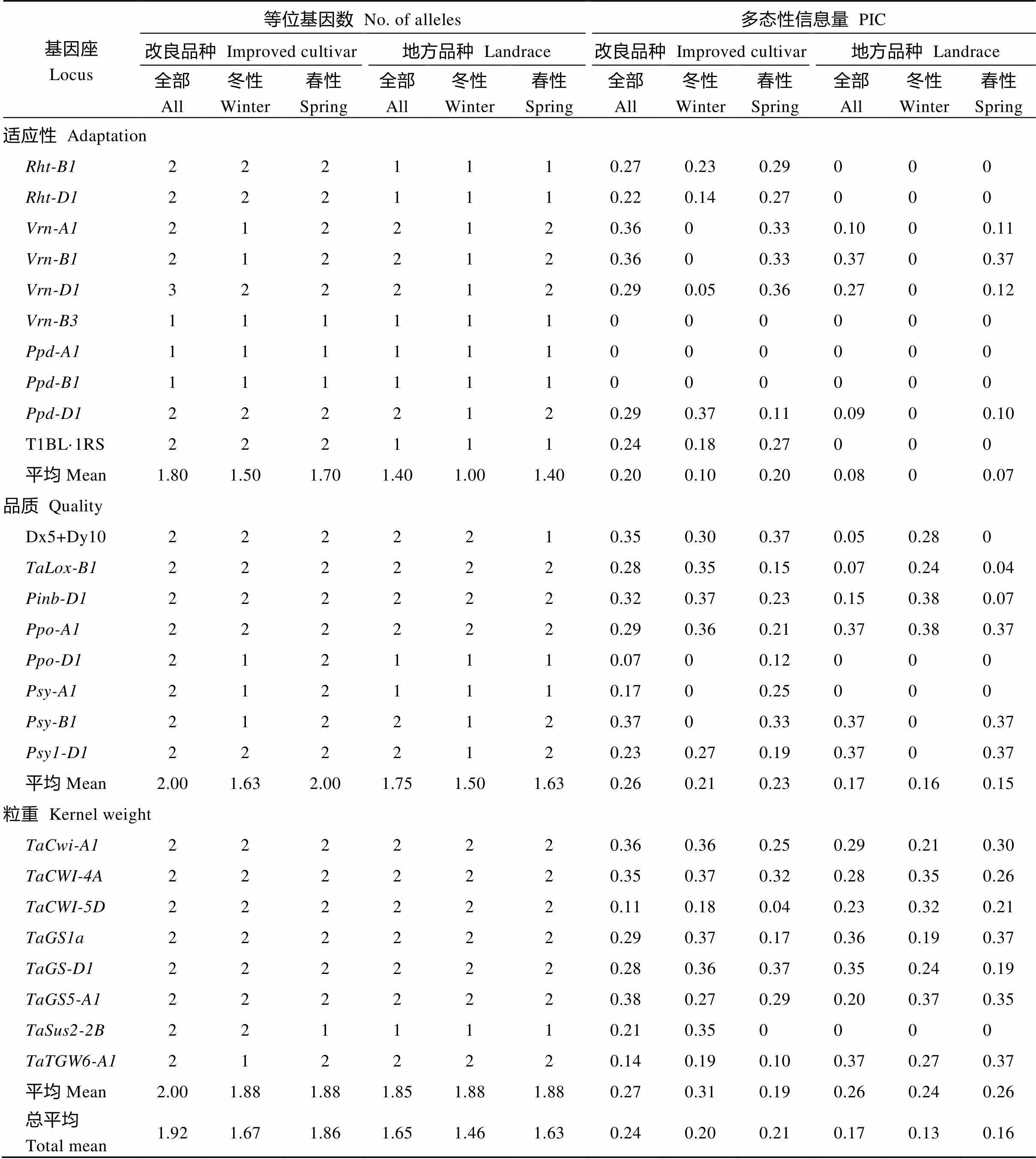
表2 新疆改良品种与地方品种适应性、品质和粒重性状相关基因座等位基因数目和多态性信息含量比较
2.4 群体结构分析
对供试材料52个等位基因遗传多样性的分子检测结果分析发现, 改良品种和地方品种两类群体间、群体内个体间以及个体间差异都极显著(< 0.01), 这三者分别解释总变异的23.9%、71.5%和4.7% (表3)。
基于主成分的群体结构分析发现, 通过第1主成分(PC1)和第2主成分(PC2)聚类能够将136份资源划分为改良品种和地方品种两类, 其中冬性改良品种和春性改良品种也各自聚为一类(图4)。

表3 遗传多样性分子方差分析
**表示在0.01水平上差异显著。**denotes significant difference at the 0.01 probability level.
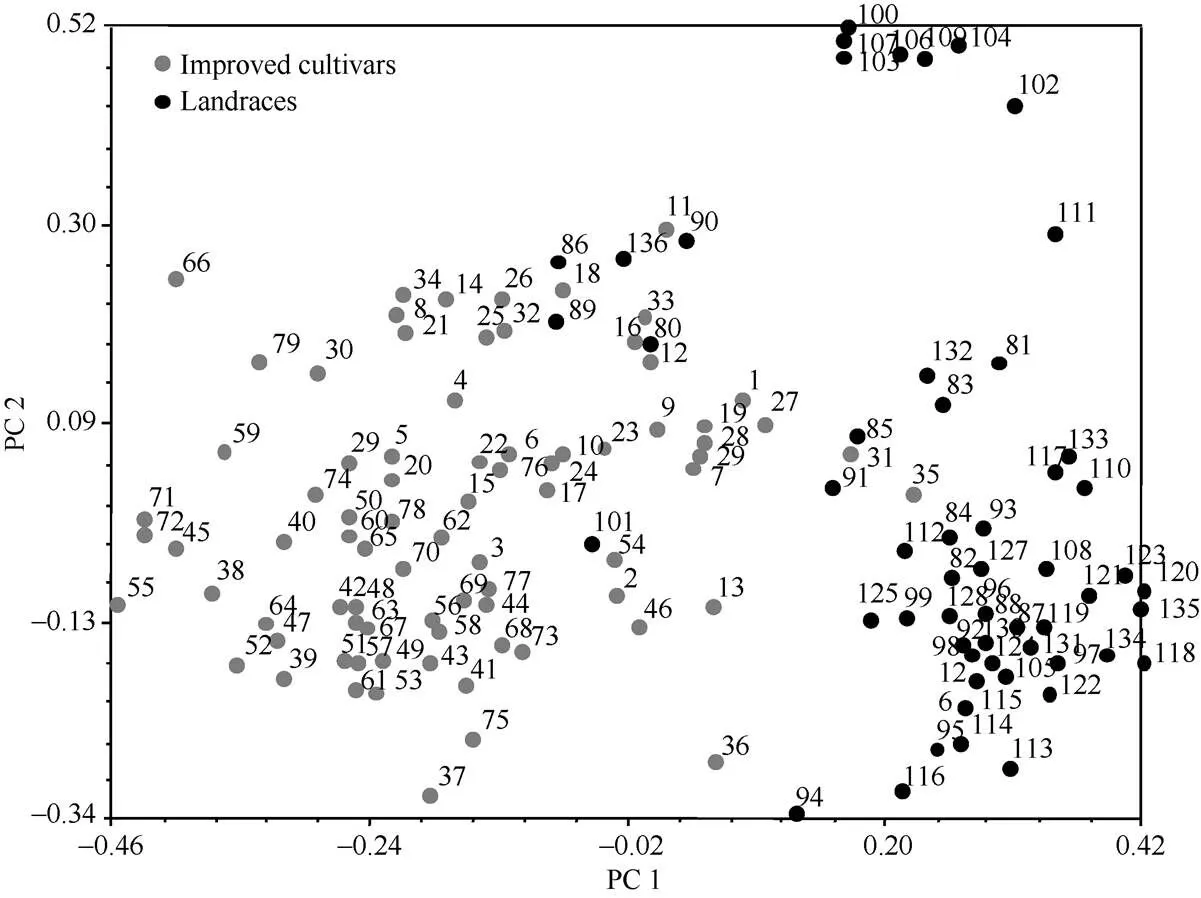
图4 136份新疆小麦资源主成分分析
1~79: 改良品种, 其中1~35为冬性品种, 36~79为春性品种; 80~136: 地方品种, 其中80~86为冬性地方品种, 87~136为春性地方品种。图中数字代表的资源名称参考附表1和附表2。
1–79: improved cultivars, including winter (1–35) and spring (36–79) accesions; 80–136: landraces, including winter (80–86) and spring (87–136) accesions. Numbers represent germplasm names given in Supplementary Tables 1 and S2.
3 讨论
新疆改良品种比地方品种含有更高的优异等位变异频率, 充分反映了育种是将优异等位基因不断选择和聚合的过程。矮秆等位变异和在我国小麦生产上发挥了重要的作用。和在新疆改良品种的分布频率分别为79.7%和15.2% (表1), 至少含有其中一个矮秆等位变异的频率达到85%, 表明这2个等位变异已经广泛应用于新疆小麦育成品种。杨松杰等[33]发现,等位变异在新疆冬春小麦(8份)中的分布频率为62.5%,等位基因分布频率为12.5%。Meng等[4]证明, 80%以上建国以来中国大面积推广品种(80份)至少含有两个矮秆等位变异中的一个。
光周期基因是影响抽穗期的一类重要基因[7]。在和基因座, 新疆改良品种和地方品种没有差异, 都只含有一种等位变异(表1)。究其原因, 一是这2个基因座的功能标记是基于日本小麦品种开发的, 对中国品种可能不适用[10]; 二是引起抽穗期变化的可能是拷贝数, 目前这种推测在基因座已经被证实[34]。
20世纪80年代后我国育成的小麦品种中38%~43%含有T1RL·1BS[35-36]。新疆小麦品种也不例外, 特别是1990—2010年间, 有20%的新疆小麦育成品种中含有这个易位系。T1RL·1BS易位系在提高抗病性和丰产性的同时, 也可能降低小麦面包加工品质, 这可能是2010年以后新疆小麦改良品种中并没有检测到T1BL·1RS片段的原因之一。与品质相关的优质麦谷蛋白亚基Dx5+Dy10在新疆育种品种中达到33.6%, 相比之下, 在地方品种中仅为1.8% (表1)。与对360份新疆地方品种、育成品种和国内外引进小麦品种分析结果一致, 33.9%的改良品种含有Dx5+Dy10亚基[37], 但这一数值低于黄淮冬麦区的41.1%[38]。与面粉色泽相关的高脂肪氧化酶活性等位变异、低多酚氧化酶活性等位变异和低黄色素含量等位变异在改良品种中分布频率均高于地方品种(表1)。尽管等位变异在改良品种的频率比地方品种高, 但明显低于王亮等[39]报道的86.9%。
现代育种实践使得重要农艺性状相关的优异等位变异逐渐累积。本研究中, 11个优异等位变异随着育种时期推进, 在新疆小麦改良品种中呈现不同程度的上升趋势(图3), 比如矮秆等位变异、优质相关的等位变异和高粒重等位变异, 这是不难理解的, 这些等位变异对新疆小麦生产是有利的, 因此被不断积累和保留下来。Meng等[4]通过对1949年以来80个大面积推广小麦品种研究同样发现, 与重要农艺性状相关的优异等位变异分布频率随育种时期推进呈现上升趋势。
新疆属于冬、春麦兼种区, 这一特殊生态区域要求北疆以春性品种为主, 而南疆以冬性或者抗寒性较好的弱冬性品种为主[40]。对春化基因的检测发现, 春性改良品种至少含有一个显性等位变异, 而冬性品种在所有春化基因座全部为隐性, 说明该地区小麦品种春化基因组成完全是为适应特定生态环境不断选择的结果[41]。光周期不敏感等位变异在新疆冬性地方品种和春性地方品种中分布频率很低, 而在春性和冬性改良品种中频率明显提高(表1), 表明可能经历了较强的选择压。值得注意的是, 该等位变异在春性改良品种中分布频率(95.1%)明显高于冬性改良品种(55.9%)。随着育种时期的推进分布频率在春性改良品种中呈现直线上升趋势, 而在冬性品种中2010年以前呈现直线上升, 2010年后呈现明显下降趋势, 进一步表明冬性和春性改良品种对该等位变异的选择是不一致的。其原因可能是春播春性品种要求生育期短, 光周期敏感品种会延长生育期, 不能满足生产需要, 这类品种在小麦生产实践过程中自然被淘汰; 而冬性品种在秋季播种需要经历漫长的冬季和春化过程, 这就需要冬性品种具有强的抗寒性, 光周期敏感品种能够延长营养生长过渡到生殖生长的时间, 在一定程度上避开冬季冻害, 增强抗寒性[42]。
本研究发现改良品种的平均等位基因数目、遗传多样性指数和PIC均高于地方品种(表2), 与Rasheed等[43]对巴基斯坦小麦改良品种和地方品种比较结果一致, 但与Hao等[44]对中国小麦的研究有所不同。本研究利用的所有功能标记都是基于改良品种开发的, 这样的标记对地方品种或者野生近缘种的检测效率可能会有一定的影响, 导致在地方品种中检测不到某些稀有或新的等位变异, 这是可能导致地方品种遗传多样性低的一个原因。另外, 与改良品种比较, 供试新疆地方品种没有发现矮秆等位变异和、半冬性等位变异以及T1BL·1RS易位染色体, 也没有检测到低黄色素含量等位变异和高千粒重等位变异(), 这与实际情况是一致的, 比如, 广泛应用的矮秆等位变异和主要来自于日本品种农林10号; T1BL·1RS易位系在20世纪70年代才引入我国; 含有等位变异的新冬20实际上是从河北引进的品种, 因此, 在新疆地方品种中不可能检测到这些等位变异。由于这些基因对于提高新疆小麦品种适应性、品质和产量具有重要作用, 因此在改良品种中被保留下来, 这可能是导致新疆改良品种比地方品种遗传多样性高的另一个原因。当然, 本研究仅是基于功能标记分析, 如果采用SNP等高通量分子标记, 相信地方品种在遗传多样性上会有变化。
Rasheed等[43]利用功能标记对巴基斯坦228份小麦资源进行聚类分析, 发现改良品种和地方品种被分别聚为一类。同样, 本研究利用52对功能标记就能很好地将136个新疆小麦资源聚为改良品种和地方品种两类(图4)。这充分说明地方品种和改良品种存在巨大的遗传差异。
4 结论
利用功能标记分析发现, 重要农艺性状相关的优异等位变异在新疆改良品种中频率明显高于地方品种, 而且随着育种时期的推进这些优异等位变异频率在改良品种中呈现上升趋势。在适应性和品质相关基因方面, 春性改良品种的优异等位变异频率比冬性改良品种更高。改良品种遗传多样性高于地方品种。136份新疆小麦资源被聚为改良品种和地方品种两类, 冬性和春性改良品种分别聚为一类。
[1] 吴新元.新疆优质小麦产业化进展. 新疆农业科学, 2010, 47(增刊2): 1–2 Wu X Y. The industrializing progress of high-quality wheat in Xinjiang., 2010, 47 (suppl-2): 1–2 (in Chinese)
[2] 曹连莆, 艾尼瓦尔. 新疆小麦高产优质育种的进展与策略. 石河子大学学报(自然科学版), 1998, 2(增刊): 1–7 Cao L P, Ainiwaer. The progress and strategy on high yield and superior quality about wheat breeding in Xinjiang.(Nat Sci Edn), 1998, 2 (suppl): 1–7 (in Chinese with English abstract)
[3] 马宏. 新疆小麦品质状况浅谈. 粮食加工, 2009, 34(5): 87–88 Ma H. Preliminary study on quality in Xjnjiang wheat., 2009, 34(5): 87–88 (in Chinese)
[4] Meng L, Xiang C, Liu H, Yang L, Mai C, Yu L, Wei Y, Li H, Zhang H, Zhou Y. The impact of modern plant breeding on dominant Chinese wheat cultivars (L.) revealed by SSR and functional markers., 2018, 65: 55–65
[5] Andersen J R, Lübberstedt T. Functional markers in plants., 2003, 8: 554–560
[6] Liu Y, He Z, Appels R, Xia X. Functional markers in wheat: current status and future prospects., 2012, 125: 1–10
[7] Worland A J, Börner A, Korzun V, Li W M, Petrovíc S, Sayers E J. The influence of photoperiod genes on the adaptability of European winter wheats., 1998, 100: 385–394
[8] Fu D, Szűcs P, Yan L, Helguera M, Skinner J S, Zitzewitz J V, Hayes P M, Dubcovsky J. Large deletions within the first intron inare associated with spring growth habit in barley and wheat., 2005, 273: 54–65
[9] Yan L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A, Valarik M, Yasuda S, Dubcovsky J. The wheat and barley vernalization geneis an orthologue of., 2006, 103: 19581–19586
[10] Beales J, Turner A, Griffiths S, Snape J W, Laurie D A. A-is misexpressed in the photoperiod insensitivemutant of wheat (L.)., 2007, 115: 721–733
[11] Seki M, Chono M, Nishimura T, Sato M, Yoshimura Y, Matsunaka H, Fujita M, Oda S, Kubo K, Kiribuchi-Otobe C, Kojima H, Nishida H, Kato K. Distribution of photoperiod-insensitive alleleand its effect on heading time in Japanese wheat cultivars., 2013, 63: 309–316
[12] Ellis M, Spielmeyer W, Gale K, Rebetzke G, Richards R. “Perfect” markers for theanddwarfing genes in wheat., 2002, 105: 1038–1042
[13] Ishikawa G, Nakamura T. A new co-dominant PCR-based marker to identify the high-molecular-weight glutenin subunit combination ‘‘5+10’’ of common wheat., 2007, 103: 1–4
[14] Geng H W, He Z H, Zhang L P, Qu Y Y, Xia X C. Cloning the lipoxygenase gene on chromosome 4BS and development of functional markers in common wheat., 2012, 52: 568–576
[15] Giroux M J, Morris C F. A glycine to serine change in puroindoline b is associated with wheat grain hardness and low levels of starch-surface friabilin., 1997, 95: 857–864
[16] Sun D J, He Z H, Xia X C, Zhang L P, Morris C F, Appels, R, Ma W J, Wang H. A novel STS marker for polyphenol oxidase activity in bread wheat., 2005, 16: 209–218
[17] He X Y, He Z H, Zhang L P, Sun D J, Morris C F, Fuerst E P, Xia X C. Allelic variation of polyphenol oxidase (PPO) genes located on chromosomes 2A and 2D and development offunctional markers for the PPO genes in common wheat., 2007, 115: 47–58
[18] He X Y, Zhang Y L, He Z H, Wu Y P, Xiao Y G, Ma C X, Xia X C. Characterization of phytoene synthase 1 gene () located on common wheat chromosome 7A and development of a functional marker., 2008, 116: 213–221
[19] He X Y, He Z H, Ma W, Appels R, Xia X C. Allelic variants of phytoene synthase 1 () genes in Chinese and CIMMYT wheat cultivars and development of functional markers for flour colour., 2009, 23: 553–563
[20] Ma D, Yan J, He Z, Wu L, Xia X. Characterization of a cell wall invertase geneon common wheat chromosome 2A and development of functional markers., 2012, 29: 43–52
[21] Jiang Y, Jiang Q, Hao C, Hou J, Wang L, Zhang H, Zhang S, Chen X, Zhang X. A yield-associated gene, in wheat: its function, selection and evolution in global breeding revealed by haplotype analysis., 2015, 128: 131–143
[22] Guo Y, Sun J, Zhang G, Wang Y, Kong F, Zhao Y, Li S. Haplotype, molecular marker and phenotype effects associated with mineral nutrient and grain size ofin wheat., 2013, 154: 119–125
[23] Zhang Y, Liu J, Xia X, He Z., an ortholog of rice, is associated with grain weight and grain length in common wheat., 2014, 34: 1097–1107
[24] Wang S, Zhang X, Cheng F, Cui D. A single-nucleotide polymorphism ofgene revealed its association with kernel weight in Chinese bread wheat., 2015, 6: 1166
[25] Jiang Q, Hou J, Hao C, Wang L, Ge H, Dong Y, Zhang X. The wheat () sucrose synthase 2 gene () active in endosperm development is associated withyield traits., 2011, 11: 49–61
[26] Hanif M, Gao F, Liu J, Wen W, Zhang Y, Rasheed A, Xia X, He Z, Cao S., an ortholog of rice, is associated with grain weight and yield in bread wheat., 2016, 36: 1. https://doi.org/10.1007/s11032-015-0425-z
[27] Guo X, Wang Y, Meng L, Liu H, Yang L, Zhou Y, Zhang H. Distribution of theallele associated with facultative growth habit in Chinese wheat accessions., 2015, 206: 1–10
[28] Zhang J, Wang Y, Wu S, Yang J, Liu H, Zhou Y. A single nucleotide polymorphism at thepromoter region in common wheat is associated with vernalization response., 2012, 125: 1697–1704
[29] Liu C, Yang Z J, Li G R, Zeng Z X, Zhang Y, Zhou J P, Liu Z H, Ren Z L. Isolation of a new repetitive DNA sequence fromenables targeting ofchromatin in wheat background., 2008, 159: 249–258
[30] Liu K, Muse S V. PowerMarker: An integrated analysis environment for genetic marker analysis., 2005, 21: 2128–2129
[31] Excoffier L, Lischer H E L. Arlequin suite ver3.5: a new series of programs to perform population genetics analyses under Linux and Windows., 2010, 10: 564–567
[32] Rohlf F J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, Version 2.1. Exeter Software. Setauket, New York, 2000
[33] 杨松杰, 张晓科, 何中虎, 夏先春, 周阳. 用STS标记检测矮秆基因和在中国小麦中的分布.中国农业科学, 2006, 39: 1680–1688 Yang S J, Zhang X K, He Z H, Xia X C, Zhou Y. Distribution of dwarfing genesandin Chinese bread wheats detected by STS marker., 2006, 39: 1680–1688 (in Chinese with English abstract)
[34] Díaz A, Zikhali M, Turner A S, Isaac P, Laurie D A. Copy number variation affecting theandgenes is associated with altered flowering time in wheat ()., 2012, 7: e33234
[35] 周阳, 何中虎, 张改生, 夏兰琴, 陈新民, 高永超, 井赵斌, 于广军. 1BL/1RS易位系在我国小麦育种中的应用. 作物学报, 2004, 30: 531–535 Zhou Y, He Z H, Zhang G S, Xia L Q, Chen X M, Gao Y C, Jing Z B, Yu G J. Utilization of 1BL/1RS translation in wheat breeding in China., 2004, 30: 531–535 (in Chinese with English abstract)
[36] 李洪杰, 王晓鸣, 宋凤景, 伍翠平, 武小菲, 张宁, 周阳, 张学勇. 中国小麦品种对白粉病的抗性反应与抗病基因检测. 作物学报, 2011, 37: 943–954 Li H J, Wang X M, Song F J, Wu C P, Wu X F, Zhang N, Zhou Y, Zhang X Y. Response to powdery mildew and detection of resistance genes in wheat cultivars from China., 2011, 37: 943–954 (in Chinese with English abstract)
[37] 王亮, 穆培源, 徐红军, 刘丽, 何中虎, 夏先春, 庄丽, 桑伟, 韩新年, 聂迎彬. 新疆小麦品种高分子量麦谷蛋白亚基组成分析. 麦类作物学报, 2008, 28: 430–435 Wang L, Mu P Y, Xu H J, Liu L, He Z H, Xia X C, Zhuang L, Sang W, Han X N, Nie Y B. Compositions of HMW-GS in wheat varieties and advanced lines from Xinjiang., 2008, 28: 430–435 (in Chinese with English abstract)
[38] Yang F P, Wang L H, Wang J W, He X Y, Zhang X K, Shang X W, Yang W X, Xia X C, He Z H. Characterisation of high- and low-molecular-weight glutenin subunit genes in Chinese winter wheat cultivars and advanced lines using allele-specific markers and SDS-PAGE., 2010, 61: 84–91
[39] 王亮, 穆培源, 徐红军, 庄丽, 桑伟, 韩新年, 聂迎彬, 夏先春,何中虎.新疆小麦品种中多酚氧化酶(PPO)活性基因等位变异的分布. 麦类作物学报, 2008, 28: 766–771 Wang L, Mu P Y, Xu H J, Zhuang L, Sang W, Han X N, Nie Y B, Xia X C, He Z H. Distribution of allelic variations of polyphenol oxidase genes in Xinjiang wheat cultivars., 2008, 28: 766–771 (in Chinese with English abstract)
[40] 王荣栋, 孔军, 陈荣毅, 张伟. 新疆小麦品质生态区划. 新疆农业科学, 2005, 42: 309–314 Wang R D, Kong J, Chen R Y, Zhang W. Classification on Xinjiang wheat regions based on ecological quality., 2005, 42: 309–314 (in Chinese with English abstract)
[41] 曹霞, 王亮, 冯毅, 徐红军, 穆培源, 张晓科, 张影全. 新疆小麦品种春化和光周期主要基因的组成分析. 麦类作物学报, 2010, 30: 601–606 Cao X, Wang L, Feng Y, Xu H J, Mu P Y, Zhang X K, Zhang Y Q. Combination of vernalization and photoperiod main genes in Xinjiang wheat., 2010, 30: 601–606 (in Chinese with English abstract)
[42] Mahfoozi S, Limin A E, Hayes P M, Hucl P, Fowler, D B. Influence of photoperiod response on the expression of cold hardiness in wheat and barley., 2000, 80: 721–724
[43] Rasheed A, Xia X, Mahmood T, Ouraishi U M, Aziz A, Bux H, Mahmood Z, Mirza J I, Mujeeb-Kazi A, He Z. Comparison of economically important loci in landraces and improved wheat cultivars from Pakistan., 2016, 56: 287–301
[44] Hao C, Wang L, Ge H, Dong Y, Zhang X. Genetic diversity and linkage disequilibrium in 407 Chinese bread wheat (L.) revealed by SSR markers., 2011, 6: e17279
附表1 79份新疆小麦育成品种基本信息
Supplementary table 1 General information on 79 wheat improved cultivars from Xinjiang

编号Code名称Name系谱Pedigree冬/春性Winter/ spring来源Origin审定或育成年份Released/ bred year 1新冬2号热依木夏/亥恩·亥德冬新疆农业科学院粮食作物研究所1966 2新冬5号巴克甫克/北京7号冬新疆农业科学院粮食作物研究所1969 3新冬7号新冬2号/敖萨德3号冬新疆农业科学院粮食作物研究所1969 4新冬14热衣木夏/乌克兰83冬新疆农垦科学院作物研究所1984 5新冬15 新冬2号/中引5号冬新疆农业科学院粮食作物研究所1989 6新冬16 (巴克甫克/亥恩·亥德)/70-4冬新疆生产建设兵团农七师农业科学研究所1988 7新冬17 新冬14/安选5号冬新疆农垦科学院作物研究所1994 8新冬18 N.S11-33/新冬3号冬新疆农业科学院粮食作物研究所1994 9新冬19 (170/阿夫乐尔)/H-矮82-6 F0种子辐照冬新疆农业科学院核技术生物技术研究所1995 10新冬20冀875018冬新疆农业科学院粮食作物研究所1995 11新冬21 318/88122冬新疆农业科学院核技术生物技术研究所1997 12新冬22 (诺斯塔/花春84-1)/(76-4/洛夫林13)冬新疆生产建设兵团农七师农业科学研究所1996 13新冬23 美国引进F1分离群体/花培品系88-136冬新疆农业科学院核技术生物技术研究所2000 14新冬24 9245/冀6159冬新疆农业科学院粮食作物研究所2003 15新冬27从中国农业大学引进高代品系系谱选择冬新疆农垦科学院作物研究所2005 16新冬28 92-45/新冬20冬新疆农业科学院粮食作物研究所2005 17新冬29 PH82-2-2/鲁植79-1冬新疆生产建设兵团农四师农业科学研究所2005 18新冬30不详冬新疆农垦科学院作物研究所2007 19新冬31 (4114/新冬14/丰收)/(新冬18/中引85)冬新疆农业科学院粮食作物研究所2007 20新冬3373-13-36/82-4009冬石河子农业科技开发研究中心粮食油料作物研究所2009 21新冬36 (89(813)/新冬18)/晋农207冬新疆农垦科学院作物研究所2011 22新冬37 京411/贵农15冬中国农业科学院作物科学研究所2012 23新冬38 (矮秆916/9133)/伊农16冬新疆生产建设兵团农四师农业科学研究所2011 24新冬41 石冬8号/新冬22冬石河子农业科学研究院2013 25新冬48 (新冬22/M844)/石冬8号冬石河子农业科学研究院2015 26新冬51 (藁城8901优/新冬18)/冀5473冬新疆农垦科学院作物研究所2015 27新冬52 新冬17/95-7-13-2冬石河子农业科学研究院粮油作物研究所2015 28伊农16(白壳欧柔/72-829/K2-13)/ (72-629-7141-64/K2-13)冬新疆生产建设兵团农四师农业科学研究所1994 29伊农18[(77224/R2-13)/(7113-9/76-629)]/[(运动1号/00089)/(工农10-3/72-原78)]冬新疆生产建设兵团农四师农业科学研究所1999 30奎花1号京花1号/奎冬3号冬新疆生产建设兵团农七师农业科学研究所1991 31石冬7号87-5048/昌冬5号(76-165)冬石河子农业科技开发研究中心1999 32石冬8号昌冬5号(76-165)/82-4009冬石河子农业科技开发研究中心2003 33石冬9号轮回选择冬新疆农垦科学院作物研究所2003 34喀冬1号巴克甫克/亥恩·亥德冬喀什地区农业科学研究所1972 35喀冬4号华北187系选冬喀什农业学校1973 36新春3号西特·赛洛斯/奇春4号 F0种子辐照春新疆农业科学院核技术生物技术研究所1985 37新春4号151/74-6//74-16/沃尔森春新疆农业科学院粮食作物研究所/昌吉农业科学研究所1990 38新春5号繁6/6038春新疆农垦科学院作物研究所1990 39新春6号中7906/新春2号春新疆农业科学院核技术生物技术研究所1993
(续附表1)

编号Code名称Name系谱Pedigree冬/春性Winter/ spring来源Origin审定或育成年份Released/ bred year 40新春7号中7906/新春2号春新疆农业科学院核技术生物技术研究所1995 41新春8号CO7/21-23春石河子大学农学院1997 42新春9号引进春麦品系NS-65春新疆农业科学院核技术生物技术研究所1999 43新春10号9-3-3/新春4号春新疆农业科学院粮食作物研究所2002 44新春11 新春2号/86-7春石河子大学农学院2002 45新春12 8021/77-13春新疆农业科学院粮食作物研究所2003 46新春13不详春不详2003 47新春14 不详春新疆农业科学院核技术生物技术研究所2004 48新春15 F5繁24/85307春新疆农业科学院粮食作物研究所2004 49新春16 86-6B/93鉴9春新疆农垦科学院作物研究所2004 50新春17 新春6号/NS64春新疆农业科学院核技术生物技术研究所2005 51新春18 不详春不详2005 52新春19 不详春石河子大学农学院2005 53新春20 昌春6号/墨西哥M85-30春新疆农业科学院粮食作物研究所2006 54新春21 NS-23-3/青海946春新疆生产建设兵团农五师农业科学研究所2006 55新春22 Tal/永1265春新疆农垦科学院作物研究所; 宁夏永宁县小麦育繁所2006 56新春23 CIMMYT引进F2/88-136 春新疆农业科学院核技术生物技术研究所2006 57新春24不详春不详2006 58新春25 不详春新疆农业科学院奇台春小麦试验场2006 59新春26 新春9号/新春6号春新疆农业科学院核技术生物技术研究所2007 60新春27 91I82299/21-4春新疆农业科学院粮食作物研究所2007 61新春28 CIMMYT引进, 原代号为01-25春新疆农垦科学院作物研究所2007 62新春29 85-56/25-3春新疆农业科学院粮食作物研究所2008 63新春30 新春9号/新春6号春新疆农业科学院核技术生物技术研究所2009 64新春31 12-25/96-5春石河子大学农学院2009 65新春32 97-18/永良11春新疆生产建设兵团农五师农业科学研究所2009 66新春33 新春9号/新春6号春新疆农业科学院核技术生物技术研究所2010 67新春34 88(13)/5×44春新疆农业科学院粮食作物研究所2011 68新春35 巴96-4870/93鉴29春新疆农垦科学院作物研究所2011 69新春36 21-6/黑小麦春新疆生产建设兵团农十三师农业科学研究所2011 70新春38 原212/97-46-3春新疆农垦科学院作物研究所与新疆九禾种业有限责任公司2012 71宁春23 (宁春4号/中7906)/陕农7855春宁夏农林科学院作物科学研究所1995 72宁春32 建三江- 6918/高代品系1658春宁夏农林科学院作物科学研究所2002 73宁春35 宁春16/永A71春宁夏农林科学院作物科学研究所2003 74宁春37 从南非引进春宁夏农林科学院作物科学研究所2005 75昌春6号(82A3/021)/(78A131/03312)春新疆农业科学院奇台春小麦试验场1994 76抗旱1号H101/C8501春新疆生产建设兵团农四师农业科学研究所2013 77青春5号阿勃/欧柔春青海省农业科学研究院1969 78新曙光1号阿勃/欧柔春黑龙江省农业科学研究院1971 79吐春9号(80B63/02126)/(77A7/834)春新疆农业科学院奇台春小麦试验场1998
附表2 57份新疆小麦地方品种基本信息
Supplementary table 2 General information on 57 wheat landraces from Xinjiang

编号Code名称Name冬/春性Winter/spring来源Origin编号Code名称Name冬/春性Winter/spring来源Origin 80库车白冬麦冬新疆库车109直芒春新疆焉耆 81白冬麦冬新疆呼图壁110佛手麦春新疆库车 82小白冬麦冬新疆呼图壁111稻麦子春新疆乌什 83阿克库孜盖冬新疆库尔勒112高拉山春麦春新疆疏附 84热衣木夏冬新疆新和113白芒红麦春新疆叶城 85长巴什曼冬新疆叶城114古玛尔汗春新疆墨玉 86纳瓦提然冬新疆新和115克拉黑麦春新疆墨玉 87阿克脱哈尔春新疆哈巴河116阿及麦春新疆墨玉 88其力克春新疆洛浦117白吐里克春新疆墨玉 89黑芒春新疆米泉118西藏稻麦子春新疆洛浦 90白光头春新疆昌吉119其力克春新疆洛浦 91红春麦春新疆昌吉120买甚春麦春新疆于田 92木锨棒春新疆昌吉121古尔汗满春新疆于田 93疙瘩头春新疆昌吉122阿克禾孜干春新疆于田 94金包银春新疆昌吉123吐里克春新疆和田 95兰麦春新疆昌吉124克兹买克春新疆和田 96黑芒春春新疆米泉125米麦米尔春新疆和田 97白春麦春新疆玛纳斯126大头郞春新疆哈密 98无芒麦春新疆鄯善127小白芒(白)春新疆青河 99分枝麦春新疆哈密128小白芒(红)春新疆青河 100大白麦春新疆哈密129小红芒1春新疆青河 101黄库班克春新疆伊宁130小红芒2春新疆青河 102比热巴沙尔春新疆霍城131小红麦春新疆巴里坤 103波兰麦春新疆吐鲁番132托力克春新疆和田 104黑头麦春新疆塔城133金包银(红)春新疆伊吾 105分枝黑芒春新疆塔城134金包银(白)春新疆伊吾 106良山麦子春新疆阿勒泰135红穗无芒春新疆阿勒泰 107黑库班克春新疆阿勒泰136阿克贾克春新疆和田 108兰壳兰芒春新疆阿勒泰
Functional Markers Reveal Genetic Variations in Wheat Improved Cultivars and Landraces from Xinjiang
JIAN Da-Wei1,2, ZHOU Yang2, LIU Hong-Wei2, YANG Li2, MAI Chun-Yan3, YU Li-Qiang4, HAN Xin-Nian5, ZHANG Hong-Jun2,*, and LI Hong-Jie2,*
1Institute of Agricultural Science, the Fourth Division of Xinjiang Production and Construction Corps, Yining 835000, Xinjiang, China;2Institute of Crop Sciences / National Key Facility for Crop Gene Resources and Genetic Improvement, Chinese Academy of Agricultural Sciences, Beijing 100081, China;3Xinxiang Innovation Center for Breeding Technology of Dwarf-male-sterile Wheat, Xinxiang 453731, Henan, China;4Zhaoxian Experiment Station, Shijiazhuang Academy of Agriculture and Forestry Sciences, Zhaoxian 515300, Hebei, China;5Key Laboratory of Xinjiang Production and Construction Corps for Cereal Quality Research and Genetic Improvement, Shihezi 832000, Xinjiang, China
Illuminating genetic variations in the improved cultivars and landraces of wheat from Xinjiang is important for further improvement of wheat in this region. In this study, 52 functional markers were used to examine 136 Xinjiang wheat accessions. Compared with the improved cultivars, the semi-dwarfing allelesand, facultative allele, T1BL·1RS translocation,allele for high lipoxynase activity,allele for lower polyphenol oxidase activity,allele for low yellow pigment content and() allele for high kernel weight were absent in the landraces. Furthermore, the frequencies of photoperiod insensitivity allele(77.6%), strong gluten subunits Dx5+Dy10 (35.4%),allele (25.0%) for hard grain texture, and(63.3%),() (33.8%),() (93.7%),() (77.9%),(78.5%),(50.0%), and(92.1%) alleles associated with high kernel weight were higher in the improved cultivars than in the landraces. The frequencies of most favorable alleles displayed a discontinuously increasing trend over breeding periods. As for the favorable alleles associated with adaptation and quality, the spring cultivars had higher frequencies than the winter cultivars. Analysis of the functional markers showed that the improved cultivars had higher genetic diversity than the landraces. Based on the principle component analysis, the wheat accessions were clustered into two groups: improved cultivars and landraces, and the improved cultivars were further divided into two groups of winter and spring cultivars. There were significant genetic differences between the improved cultivars and the landraces. The favorable alleles and allele combinations identified in this study provide important information for further improvement of wheat cultivars in Xinjiang.
; Xinjiang Uygur Autonomous Region; functional marker; agronomic traits; genetic variation
2017-08-03;
2018-01-08;
2018-01-26.
10.3724/SP.J.1006.2018.00657
本研究由国家自然科学基金项目(31771881, 31401468)和国家重点研发计划项目(2017YFD0101000, 2016YFD0101600, 2016YFD0101004, 2016YFD0100102)资助。
This study was supported by the National Natural Science Foundation of China (31771881, 31401468) and the National Key Research and Development Program of China (2017YFD0101000, 2016YFD0101600, 2016YFD0101004, 2016YFD0100102).
张宏军, E-mail: zhanghongjun01@caas.cn; 李洪杰, E-mail: lihongjie@caas.cn
E-mail: 81809902@qq.com
http://kns.cnki.net/kcms/detail/11.1809.S.20180126.0856.002.html
猜你喜欢
作物学报(2022年6期)2022-04-08
国际医学放射学杂志(2021年5期)2021-10-22
麦类作物学报(2020年7期)2020-04-01
西北农业学报(2019年10期)2019-11-20
小小说月刊(2019年18期)2019-09-24
小学生作文(中高年级适用)(2018年4期)2018-05-14
麦类作物学报(2018年4期)2018-05-11
第一财经(2017年36期)2017-09-25
农家科技中旬版(2016年7期)2016-10-20
中国火炬(2011年2期)2011-07-25
