
桃主要致敏蛋白Pru p 3 T细胞表位鉴定
2018-12-24 01:45王瑞琦
中华临床免疫和变态反应杂志 2018年5期
邓 珊,尹 佳,王瑞琦
作者单位: 100730 北京,中国医学科学院 北京协和医学院 北京协和医院变态(过敏)反应科,过敏性疾病精准诊疗研究北京市重点实验室
桃是我国成人食物过敏中较常见的致敏食物,且桃过敏反应常见于蒿花粉症患者中,是蒿花粉相关食物过敏最常见形式。桃过敏患者除了有局部接触部位荨麻疹、口腔黏膜变态反应综合征(oral allergy syndrome, OAS)外,还会发生严重变态反应。曾有研究发现,在907例严重变态反应患者中,由进食蔬菜/水果导致的严重变态反应占33%,其中桃是最常见的致敏食物。在桃诱发的严重变态反应中,71%患者并发蒿花粉症[1]。笔者前期研究已鉴定出蒿花粉相关桃过敏反应的主要致敏蛋白为Pru p 3,是相对分子质量为9 000的磷脂转移蛋白(lipid transfer protein,LTP),由91个氨基酸组成。约90%的蒿花粉相关桃过敏患者血清中存在Pru p 3 特异性IgE(specific IgE,sIgE)[2]。桃Pru p 3与蒿Art v 3(LTP)存在交叉反应,目前认为Art v 3是引起我国桃过敏反应的初始致敏蛋白[3-4]。
植物来源的LTP是相对分子质量约为9 000的蛋白质家族(90~95个氨基酸组成),广泛分布于花粉和植物类食物中,其主要功能是参与植物对外来病原微生物的免疫防御反应,当有病原体入侵时LTP表达量增加[5]。LTP具有耐加热和胃蛋白酶消化的特性,因此增加了其致敏性和引发严重变态反应的风险[6-8]。Pru p 3是最具有代表性的LTP分子。Pasquato等于2005年首次报道了Pru p 3 三维晶体结构,发现其呈高度可塑性的非对称性结构,中间包含一个可结合多个磷脂配体的空腔。这个结构基序由4个α螺旋构成,中间连接2个短环和4个二硫键,在分子的外部是3个高度保守的氨基酸序列。突变分析结果显示,随着氨基酸的插入,该区域的IgE结合能力会发生相应变化。当插入3个氨基酸后,与IgE结合的能力将明显下降。目前的研究发现Pru p 3是可被sIgE识别的区域[9],而且Pru p 3已被重组表达并用于临床诊断[10]。国外也有一些关于Pru p 3 T细胞表位的研究报道[11-12],但由于国外大多数桃过敏反应的初始致敏蛋白来源于桃,尤其地中海地区国家桃过敏患者常不并发花粉症[13],而在我国蒿花粉Art v 3被认为是引起桃过敏的交叉反应致敏蛋白[4]。同时由于种族和地域性差异,目前尚不清楚Pru p 3引起我国蒿花粉相关桃过敏反应的T细胞表位。Pru p 3 T细胞表位的确定为研究桃过敏反应的免疫学机制提供了重要信息,同时可以帮助发现与其有交叉反应的其他食物或花粉变应原的相关表位。主要T细胞表位的确定为将来利用T细胞表位合成的肽段对食物过敏患者进行特异性免疫治疗提供了理论基础。本研究以蒿花粉相关桃过敏患者和健康对照外周血单个核细胞(peripheral blood mononuclear cell,PBMC)作为效应细胞,观察其在Pru p 3 重叠肽段激活下的增生反应,对Pru p 3的主要T细胞表位进行初步研究,探讨可能参与蒿花粉和桃变应原交叉反应的T细胞表位。
1 材料与方法
1.1 研究对象
纳入蒿花粉相关桃过敏患者20例和健康对照者10例。桃过敏患者中男性6例,女性14例;健康对照者中男性5例,女性5例。蒿花粉相关桃过敏(以下简称桃过敏)患者来自北京协和医院变态反应科门诊就诊患者,需同时满足以下入组标准:(1)有夏秋季鼻炎和/或哮喘病史,同时蒿花粉提取物皮内试验结果≥“++”和蒿sIgE>3.5 kUA/L;(2)有明确进食桃2 h内出现全身皮肤风团样皮疹、口腔黏膜水肿、呼吸困难、恶心、腹痛,甚至变应性休克的病史;(3)血清sIgE检测结果显示Pru p 3 sIgE阳性、Pru p 1和Pru p 4 sIgE均阴性。
1.2 方法
1.2.1 血清特异性IgE检测:采用ImmunoCAP法测定受检者蒿花粉及桃血清sIgE水平,同时检测桃组分Pru p 1、Pru p 3和Pru p 4 sIgE水平(Thermo Fisher Scientific, Uppsala, Sweden)。血清sIgE>0.35 kUA/L判断为阳性。
1.2.2 Pru p 3多肽合成和纯化:利用多肽合成仪采用固相合成法合成8条覆盖Pru p 3.0101全部氨基酸序列的肽段,每条肽段之间重叠5个氨基酸残基(Invitrogen, Calif, USA)。8条肽段序列如下:P1(Pru p 31-16): ITCGQVSSALAPCIPY;P2(Pru p 312-27): PCIPYVRGGGAVPPAC;P3(Pru p 323-38): VPPACCNGIRNVNNLA;P4(Pru p 334-49): VNNLARTTPDRQAACN;P5(Pru p 345-60): QAACNCLKQLSASVPG;P6(Pru p 356-71): ASVPGVNPNNAAALPG;P7(Pru p 367-82): AALPGKCGVHIPYKIS;P8(Pru p 378-91): PYKISASTNCATVK。合成肽段存储于DMSO中(10 mg/ml),使用前用PBS稀释至1 mg/ml,加入细胞培养基。
1.2.3 PBMC增生试验:将新鲜分离的PBMC用含10%胎牛血清、2 mmol/L谷氨酰胺和10 mmol/L羟乙基哌嗪乙硫磺酸溶液(N-2-hydroxyethylpiperazine-N-ethane-sulphonicacid,HEPES)RPMI 1640培养基调整细胞数为1×106/ml。加入96孔培养板中,每孔200 μl,每组设3个复孔。分别加入8条肽段作为刺激物(5 μg/ml),同时设置PHA为阳性对照孔和不加任何刺激物的阴性对照孔。37 ℃、体积分数5% CO2条件下孵育5 d,然后向各孔中加入20 μl CCK-8溶液。37 ℃、5% CO2条件下孵育5 h,测定波长为450 nm处的吸光度(A)值。淋巴细胞增生水平以刺激指数(stimulation index,SI)表示,SI=变应原刺激后A450值/阴性对照A450值。SI≥2为结果阳性。
1.2.4 T细胞表位筛选:将20例桃过敏患者和10例健康对照的PBMC分别与8条Pru p 3来源的肽段共同孵育,比较桃过敏患者和正常对照PBMC增生后的SI,分析不同肽段的变应原性。
1.3 统计学方法
采用SPSS 17.0统计学软件(SPSS I,Chicago,IL,USA)进行统计分析,采用Mann-WhitneyU检验比较蒿花粉相关桃过敏患者和健康对照PBMC受Pru p 3来源各肽段刺激后的增殖水平。P<0.05为差异有统计学意义。
2 结果
2.1 患者基本情况
20例桃过敏患者年龄6~46岁,平均(25.7±11.6)岁,其中男性患者6例,占30%。患者中有季节性哮喘者9例,占45%;患有季节性鼻炎症状者16例,占80%;同时有鼻炎和哮喘症状者6例,占30%。患者桃过敏症状仅表现为OAS者4例,占20%;表现为全身反应者16例,占80%。20例蒿花粉相关桃过敏患者的基本情况、临床表现及血清sIgE水平见表1。
2.2 T细胞表位筛选结果
桃过敏患者PBMC对8条肽段表现出不同的增生能力:肽段Pru p 367-82对桃过敏患者PBMC的活化程度最高,20例桃过敏患者中对该肽段SI阳性者14例,占70%;Pru p 323-38对桃过敏患者PBMC的活化率为55%(11/20);其他各肽段的增生阳性率为10%~35%(图1,表2)。健康对照者PBMC在各肽段刺激下的增生率为0%~20%,与桃过敏患者比较差异有统计学意义(P<0.05)(图1)。

表1 20例蒿花粉相关桃过敏患者的临床资料Table 1 Clinical data of patients with mugwort pollen related peach allergy
2.3 参与蒿花粉相关桃过敏交叉反应的潜在T细胞表位
Pru p 367-82在桃Pru p 3 3D结构中的位置紧邻以往研究发现的IgE结合区域。比对Pru p 367-82和蒿花粉组分Art v 3氨基酸序列发现,Pru p 367-82与Art v 390-105氨基酸一致率为81%(图2)。

图 1 蒿花粉相关桃过敏患者和健康对照PBMC在8条来源Pru p 3肽段(P1-P8)作用下增生率Fig 1 Proliferative rate of positive responses of PBMC from peach-allergic patients and control subjects to 8 synthetic peptides (P1-P8) representing the aa sequence of Pru p 3
3 讨论
蒿花粉相关桃过敏反应是我国比较常见的一种食物过敏类型,Pru p 3是蒿花粉相关桃过敏反应的主要致敏蛋白。虽然以往有文献报道Pru p 3的T细胞表位,但由于蒿花粉Art v 3是引起我国桃过敏的初始致敏蛋白,与国外多数桃过敏反应发生机制有所不同,如地中海地区国家中桃过敏患者并不伴有花粉症。因此,导致我国蒿花粉相关桃过敏反应发生的Pru p 3 T细胞表位尚不清楚。明确Pru p 3特异性的T细胞表位有助于更深入了解该变应原的免疫学特性,为研究食物过敏发生的免疫学机制提供重要信息。此外,通过研究花粉-食物过敏综合征中食物变应原的T细胞表位有助于发现潜在的参与花粉-食物之间交叉反应的T细胞表位[14]。
变应原特异性免疫治疗是治疗变态反应性疾病唯一有效的方法[15]。目前应用低变应原性的免疫制剂降低了其与IgE的结合能力,有望成为特异性好、更安全、更有效的治疗方法[16-17]。CD4+T淋巴细胞不仅参与Ⅰ型超敏反应[18],还诱导免疫耐受,在特异性免疫治疗中发挥作用。研究发现,在疫苗接种过程中,典型的Th2型免疫反应逐渐向Th1型转化,以及生成调节性T细胞,最终抗原特异性的Th2型免疫反应受到抑制[19-20],而且T细胞表位肽段仅作用于T淋巴细胞,不会活化肥大细胞和嗜碱性粒细胞[21-22],避免治疗中发生严重变态反应的风险。综上所述,基于T细胞表位的肽段免疫治疗非常有前景。因此,明确桃主要致敏蛋白Pru p 3 的主要T细胞表位有望用于桃过敏的特异性免疫治疗。
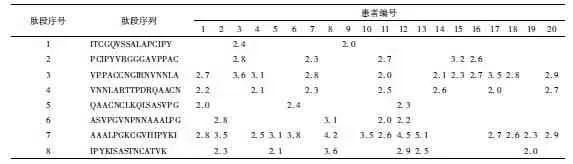
表2 蒿花粉相关桃过敏患者PBMC在P1-P8肽段刺激下细胞增生水平Table 2 PBMCs from 20 peach-allergic patients stimulated by 8 overlapping peptides representing the aa sequence of Pru p 3(SI)

图 2 蒿花粉组分Art v 3氨基酸序列以及潜在的参与蒿花粉-桃交叉反应的T细胞表位Fig 2 Amino acid sequence of mugwort Art v 3 and its potential epitope cross-reacting with Pru p 3
本研究分析Pru p 3单致敏的蒿花粉相关桃过敏患者PBMC增生情况,对Pru p 3 主要T细胞表位进行初步鉴定。通过比较8条覆盖Pru p 3分子全部氨基酸序列的合成肽段活化后的桃过敏患者PBMC增生水平,发现了两个活化率较高的T细胞表位:Pru p 367-82是主要T细胞表位,70%桃过敏患者T细胞能识别该表位,发生增生反应;而另一T细胞表位Pru p 323-38活化55%桃过敏患者外周血PBMC。值得注意的是,这两个T细胞表位均位于Pru p 3分子3D结构中IgE结合区域附近,即Pru p 367-82与Pacios等[23]报道的IgE构象表位存在序列重合,而Pru p 323-38包含了一个IgE表位区域(Pru p 323-36)[9]。本研究结果与以往文献报道的T细胞表位Pru p 365-80和Pru p 361-75有相同的氨基酸序列[24-25]。近来研究发现,Art v 3是引起我国桃过敏的交叉反应致敏蛋白。本研究比对了Pru p 367-82和Art v 390-105氨基酸序列,发现81%氨基酸残基一致,因此推测Pru p 367-82可能是参与蒿花粉-桃交叉反应的T细胞表位,当然这个结果还需要细胞学实验加以证实。
本研究还发现,健康对照者在Pru p 3各肽段的刺激下也有部分PBMC发生了细胞增生。以往文献报道,某些变应原,如Art v 1和Fel d 1 T细胞表位可以同时活化变态反应患者和健康人的T淋巴细胞,但检测其分泌的细胞因子发现,健康对照组细胞上清干扰素γ(interferon-γ,IFN-γ)/白细胞介素4(interleukin-4,IL-4)高于变态反应患者组[26-27]。这表明变态反应患者外周血T细胞在变应原刺激下发生活化和增生,分泌Th2型细胞因子,而无变态反应的正常T细胞在变应原作用下主要发生Th1型免疫反应。这就解释了为什么健康对照者PBMC在Pru p 3肽段的刺激下也发生增生反应的现象。
本研究是首个在中国蒿花粉相关桃过敏患者中针对桃主要致敏蛋白T细胞表位的研究,发现Pru p 323-38和Pru p 367-82是引起我国蒿花粉相关桃过敏反应的主要T细胞表位,但这只是一个初步研究,还需要进一步采用Pru p 3特异性的T细胞系和T细胞克隆进行T细胞表位的鉴定。明确Pru p 3致敏发生的关键T细胞表位将有助于更深入地研究蒿花粉相关桃过敏交叉反应发生的免疫学机制,同时基于T细胞表位的肽段免疫治疗是特异性免疫治疗的趋势,明确Pru p 3主要T细胞表位为将来肽段免疫治疗用于临床奠定了基础。
猜你喜欢
哈哈画报(2022年8期)2022-11-23
传染病信息(2022年4期)2022-09-17
中国高原医学与生物学杂志(2022年3期)2022-06-22
浙江医学(2022年7期)2022-05-16
中山大学学报(医学科学版)(2022年1期)2022-02-28
中国中西医结合耳鼻咽喉科杂志(2021年3期)2022-01-01
医学综述(2021年3期)2021-03-27
智慧健康(2021年2期)2021-03-15
温州医科大学学报(2019年4期)2019-04-28
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30
