
水稻粒形遗传调控研究进展
2019-03-18 12:56曾智锋杨维丰柯善文
安徽农业科学 2019年5期
尉 鑫,曾智锋,杨维丰,韩 婧,柯善文*
(1.华南农业大学农学院,广东广州 510642;2.亚热带农业生物资源保护与利用国家重点实验室,广东广州 510642)
水稻(OryzasativaL.)是重要的粮食作物,全球50 %以上的人口以稻米为主食[1]。随着人类活动的影响以及工业化生产的加剧,有效耕地面积持续下降,近年来世界水稻的产量一度停滞不前[2]。为了解决粮食供应不足的问题,提高水稻单产已经成为粮食总产量增加的重要途径。水稻产量主要由3个重要因素决定:千粒重、有效穗数和穗粒数。其中,有效穗数由植株的分蘖能力决定,穗粒数主要取决于枝梗数和结实率,而千粒重则受籽粒的形状、大小以及灌浆饱满度的影响[3]。
稻米的长度、宽度和长宽比影响水稻的粒形,而粒形是稻米品质鉴定的重要组成部分。中国优质稻谷标准规定:长宽比不低于2.8的列为优质籼稻品种。育种家在选育优质稻米品种过程中将谷粒形状作为重要指标之一[4]。多数QTL(quantitative trait loci)的分析结果表明,水稻粒形的遗传受多基因控制,且存在一因多效的现象,不同粒形性状间有不同程度的相关性[5-9]。笔者从水稻粒形相关基因的克隆及功能分析方面综述了前人的研究成果并展望研究前景,以期为水稻粒形基因克隆及高产育种提供理论指导。
1 水稻籽粒的结构
水稻加工成的消费产品主要来源于籽粒,籽粒中主要包含淀粉等营养成分,这就为人体的健康饮食提供了良好的材料。通常植物种子主要由3部分组成:胚、胚乳和种皮。与其他谷类作物一样,水稻种子在结构上明显不同于双子叶植物的种子,例如拟南芥(图1)。拟南芥种子中的胚占据了大部分的空间,其主要营养物质储存于子叶中,而水稻种子中的大部分空间被胚乳占据,并且胚乳中储存了大量的淀粉、少量的蛋白质和脂质以及其他微量物质(图1),因此水稻的胚乳部分是人们主要的采食部位。水稻的粒形可以影响胚乳部分的大小并决定其重量,而其重量与粒重直接相关,所以水稻粒形是决定水稻产量的重要因素。

图1 水稻和拟南芥成熟种子的纵切面[10]Fig.1 Longitudinal sections of mature seeds of Arabidopsis and rice [10]
2 水稻粒形相关基因的克隆
图位克隆技术是挖掘功能基因的重要技术,该方法首先利用表型上差异较明显的材料进行杂交处理来构建初级定位群体,例如F2群体、RIL群体等;然后采用分子标记筛选并结合表型分析的操作来发现效应值较大的QTLs,并且在此基础上继续发展新的作图群体,进一步缩短目标区间的遗传距离;最后对定位区间的候选基因进行分析与验证,以此来确定目的基因的正确位置。
到目前为止,研究学者们已经克隆了一些直接或间接与水稻粒形调控相关的基因(表1)。其中仅有少数几个基因对水稻的粒形起主要调控作用,其他的大部分基因均表现出一因多效的现象,它们是水稻粒形调控的微效基因,这些微效基因对水稻其他组织器官发育的影响更为显著。
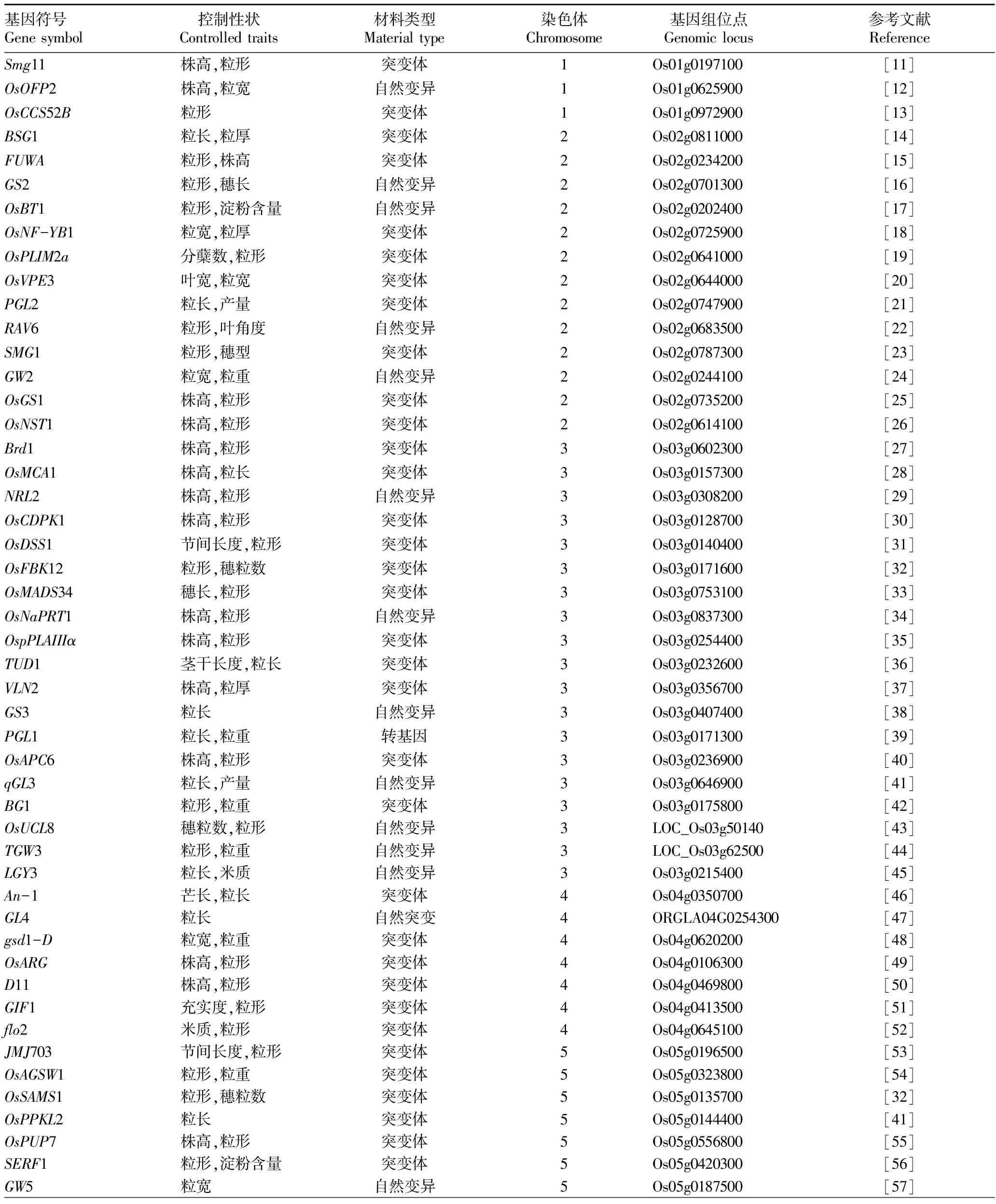
表1 已克隆的水稻粒形相关基因

接下表
注:数据主要来源于国家水稻数据中心,基因数据库,http:∥www.ricedata.cn/gene/
Note:Data originated from China Rice Data Center and gene database,http:∥www.ricedata.cn/gene/
总结发现,93个已被克隆的水稻粒形相关基因分布于12对染色体上的数目各不相同,其中第3号染色体上的最多,第1、9、12号染色体上的较少(图2)。经过多年来研究学者的努力,比较多的水稻粒形调控基因被成功克隆,但是与利用不同背景材料得到定位的粒形QTLs相比仍然很少,而且被克隆的粒形基因中大多数源自突变体材料,很难直接运用于生产实践,所以基于自然变异条件下的基因定位、克隆对作物品种的改良和应用具有更加重要的意义。

图2 已克隆水稻粒形基因在染色体上的分布Fig.2 Distribution of cloned grain shape genes on chromosomes in rice
3 水稻粒形主效基因的克隆
近年来,许多重要的水稻粒形基因都是利用栽培品种间表型差异较大的自然变异对目标QTL进行克隆而得到的。这类基因通常只对水稻粒形性状具有显著效应,并且不影响植物体的正常形态和发育,例如GW2、GS2、GS3、qGL3、TGW3、GL4、GW5、GS5、TGW6、GW7、GLW7和GW8等。
调控水稻粒长的主效基因主要包括GS3、qGL3、GL4和GLW7。Fan等[38]利用大粒的明恢63与小粒的川7构建NIL群体,利用5 740个BC3F2个体中的1 384个隐性个体和11个分子标记,将GS3定位在第3号染色体上着丝粒附近的7.9 kb区间内。Zhang等[41]选用小粒的N643和大粒的N411进行杂交,通过分析BC2F3代的结果将qGL3范围缩小到RM15548和RM3513标记之间,高精度连锁分析将qGL3进一步定位到XJ39和XJ26之间的46.6 kb区间内。Wu等[47]将渗入系中大粒的GIL25和小粒的IRGC102305进行杂交,对F2代的186个单株进行检测,将目标区域确定在第4号染色体上的RM3335和RM5608之间,最后利用后代纯合的分离株系将GL4定位在M3和M4之间的5.9 kb区域内。Si等[74]利用小粒性状和大粒性状的381个不同水稻品种进行全基因组关联分析(GWAS),成功克隆了一个位于第7号染色体上控制粒长的主效QTL-GLW7,其编码植物特殊的转录因子OsSPL13。
调控水稻粒宽的主效基因主要包括GW2、GW5、GS5和GW8。Song等[24]利用小粒的FAZ1与大粒的WY3构建BC2F2群体,利用加密标记不断地缩短定位区间,最后通过对BC3F4群体的检测,将GW2定位在第2号染色体的标记W024和W004之间的8.2 kb区间内。Shomura等[97]利用窄粒Kasalath和宽粒日本晴杂交自交产生的F2代分离群体进行QTL定位分析从而将qsw5精细定位于第5号染色体短臂端的2 263 bp区间内,但是Liu等[57]重新将GW5定位于第5号染色体短臂端,编码钙调素结合蛋白。Li等[59]利用珍汕97和H94构建DH群体,与此同时多次回交珍汕97发展作图群体,从而将GS5定位在第5号染色体上短臂端的11.6 kb区间内。Wang等[83]利用华粳籼74(受体亲本)和Basmati385、Basmati370(供体亲本)构建的单片段代换系,鉴定出位于第8号染色体上长臂端的主效QTL-GW8,继续进行精细定位以及高精度连锁分析,将GW8定位在第8号染色体上的7.5 kb区间内。
调控水稻粒形长宽比的主效基因主要包括GS2、TGW3和GW7。Hu等[16]利用小粒Zhonghua11和大粒Baodali构建分离群体,将GS2定位在第2号染色体上的7.4 kb区间内。Ying等[44]将籼稻小粒品种黄华占(HHZ)和粳稻大粒品种JZ1560进行杂交构建RIL群体,利用SLAF-seq测序技术构建遗传图谱并进行QTL定位分析,成功检测到一个位于第3号染色体上控制籽粒大小和千粒重的主效QTL-TGW3。Wang等[72]利用TaifengA(TFA)和HJX74构建作图群体,最终将GW7基因准确定位于第7号染色体的2.6 kb区间内,其表达量上调可导致细长谷粒的形成。
综上所述,水稻粒形主效基因的克隆主要涉及粒长、粒宽和长宽比3个方面,但是定位方法有所不同,大致分为3种:第一种,利用表型差异较大的双亲品种构建F2代分离群体、高回交世代群体、DH群体、NIL群体和RIL群体等作图群体进行基因克隆;第二种,利用渗入系、CSSL和SSSL排除遗传背景干扰,针对目标性状进行基因克隆;第三种,利用测序技术,例如GWAS,针对某一性状对大量不同品种材料进行测序分析,从而直接找到目标QTL,然后进行基因的克隆。就目前来看,利用测序分析技术进行基因克隆的方法与其他2种相比占有一定的优势,不但节省了科研人员的工作量而且大大缩短了试验时间。
4 水稻粒形基因间的互作
4.1GW2、GS3、GIF1和GW5/qSW5之间的互作Yan等[98]为了研究GS3、GW2、qSW5/GW5和GIF1这4个基因间的关系,对GS3-RNAi、GW2-RNAi系和qSW5的CSSL进行了基因表达分析。发现qSW5和GW2正向调控GS3的表达,GW2的表达能够下调qSW5的表达,qSW5正向调控GIF1的表达,而GW2和GS3负向调控GIF1的表达;此外,利用一个含有180个水稻品种的自然群体,详细分析了qSW5和GS3的等位基因作用,结果表明qSW5对粒长的作用受到GS3的影响,GS3对粒宽的作用受到qSW5的影响(图3)。该研究为更加深入理解水稻籽粒大小发育的分子机制及提高水稻产量提供了有用的信息。

图3 GS3、GW2、qSW5/GW5和GIF1之间的互作Fig.3 Interaction between GS3,GW2,qSW5/GW5 and GIF1
4.2qGL3/GL3.1、OsPPKL2和OsPPKL3之间的互作Zhang等[41]发现位于OsPPKL1中第2个Kelch功能域上保守的AVLDT区域的D364E的稀有等位变异qgl3导致长粒表型;OsPPKL1和OsPPKL3在水稻粒长调控中发挥负调节子的作用,而OsPPKL2是一个正调节子;Kelch功能域在OsPPKL1的负调节功能中是充分必要的(图4)。田间试验表明,优异等位基因qgl3的利用可以通过增加粒长、灌浆速率和粒重而显著提高常规稻和杂交稻的产量。

图4 OsPPKL1、OsPPKL2和OsPPKL3之间的互作Fig.4 Interaction between OsPPKL1,OsPPKL2 and OsPPKL3
4.3PGL1、PGL2和APG之间的互作Heang等[39]的研究表明一对拮抗作用的碱性螺旋-环-螺旋(bHLH)蛋白通过调控内外颖细胞长度,参与决定水稻籽粒长度。在内外颖过量表达PGL1,转基因水稻籽粒长度和重量增加,是一种结合DNA的典型bHLH的抑制子。在内外颖过量表达PGL2同样可以增加水稻粒长和粒重,并且这种增加与转基因表达水平相关。PGL1的互作因子APG与其产生拮抗作用,是一个负调控因子。PGL1和APG在体内互作,都定位在核内,沉默APG的表型与过量表达PGL1一致,2个已知的籽粒长度相关基因GS3和SRS3的表达在PGL1过量表达株系和APG沉默植株中基本不受影响。PGL2与典型bHLH蛋白APG在体外和体内互作,与PGL1功能冗余,通过与APG形成异源二聚体抑制APG的功能,从而正向调节水稻籽粒长度。PGL1-APG代表了一种新的籽粒长度和重量调控的途径(图5)。

图5 PGL1、APG、GS3和SRS3之间的互作Fig.5 Interaction between PGL1,APG,GS3 and SRS3
4.4其他粒形基因间的互作Wang等[72]研究发现,GW7编码一种募集基蛋白(TONNEAU1),在细胞分裂的过程中调控横向细胞分裂减少,纵向细胞分裂增加,过量表达可使籽粒变得更加细长。GW8(OsSPL16)是对粒宽具有调控作用的包含SBP结构域的转录因子,可以与GW7启动子结合从而抑制其表达,并且OsSPL16-GW7模块的相互作用可同时高稻米品质和稻米产量。与此同时,gs3和gw8的双突变材料(以HJX74为背景的NIL-gw8gs3)表明谷粒细长的程度与亲本相比呈现出累加作用,从而证明了GW8与GS3的调控途径是相互独立的。
Hu等[99]研究证明,TGW3编码一种蛋白激酶OsGSK5/OsSK41,此蛋白激酶属于GSK3/SHAGGY-Like家族。OsARF4是一种转录抑制因子,功能缺失可增加水稻粒长,在植物生长素途径中发挥作用。两者相互作用会使OsARF4磷酸化,而OsSK41和OsARF4的共同表达导致OsARF4的转录抑制功能增强。进一步的研究表明,GL3.3(TGW3)与GS3之间存在相互作用,其叠加效应可使水稻籽粒显著增大,已经应用于大粒粳稻的品种选育中[100]。
5 水稻粒形调控的网络途径
在水稻和拟南芥中,调控种子粒形的信号途径是相对保守的,均是通过影响胚乳和胚的生长发育来调控籽粒大小,并且是通过多种信号途径共同作用的结果。将这些信号途径总结归纳主要分为6种:IKU信号途径、泛素-蛋白酶体途径、G-蛋白信号途径、MAPK信号途径、植物激素途径和转录调控因子途径,其中植物激素途径中主要包括的激素是油菜素内酯和生长素[101]。
这些途径中所涉及到的已克隆基因可能通过大量不同的信号通路以及某些未知通路来参与调控细胞分裂和细胞增殖的过程。其中GW2和GW5/qSW5参与泛素-蛋白酶体途径的调控;GS3和DEP1(qPE9-1)参与G-蛋白信号途径的调控;OsMKK4(SMG1)和OsMAPK6参与MAPK信号途径的调控;qGL3、CycT1;3和GS5参与植物激素途径中的油菜素内酯调控途径;BG1和TGW6参与植物激素途径中的生长素调控途径;而调控水稻粒形的大部分基因在转录调控因子途径中发挥作用,例如GW6a(OsglHAT1)、GL7、GW8、GS2、SRS5等(图6)。

图6 水稻主要粒形基因的调控解析[101]Fig.6 The major signaling pathways of grain shape control in rice[101]
目前并没有发现已克隆水稻粒形相关基因参与IKU信号途径的调控,其主要存在于拟南芥中。IKU信号途径与细胞分裂素和脱落酸信号相调控共同参与拟南芥种子结构中胚乳的形成,起到了正向调控的作用[101]。对胚乳形成具有反向调控作用的TGW6是一个控制水稻粒长和籽粒充实度的基因,其调控决定了水稻籽粒最终形态的形成[68],与IKU信号途径一样直接作用于种子胚乳的形成过程。
TGW3、LGY3、GL4、GSN1、WTG1和GS9为最新发现的水稻粒形主效调控基因,其中LGY3作用于G-蛋白信号途径[45],GSN1作用于MAPK信号途径[60],WTG1作用于泛素-蛋白酶体途径[81],GS9作用于油菜素内酯调控途径[85],而TGW3和GL4的作用途径尚不明确[44,47]。但毋庸置疑的是水稻粒形基因调控网络已初步被阐明。
6 小结与展望
由于世界人口的不断增加,耕地面积的不断减少,作为主要粮食作物的水稻产量问题一直受到人们的高度关注,而水稻粒形又与水稻产量有着非常密切的联系,因此水稻粒形基因的克隆已经成为水稻产量和米质性状研究的热点之一。然而水稻自身遗传背景的复杂性为水稻粒形的研究增加了困难,选择粒形差异较大或者遗传远缘的品种材料会起到至关重要的作用。
经过多年来研究学者的不断努力,已经有很多水稻粒形相关基因被成功克隆。笔者归纳了93个已克隆的与水稻粒形相关的基因,并且对多基因间的相互作用进行了详细介绍,同时揭示了水稻粒形调控网络。通过总结发现,一些基因既能独立对水稻粒形性状产生调控作用,也会相互联系共同存在于同一个信号调控网络,以此来实现控制细胞分裂和细胞伸长的功能,其中许多基因在育种选择过程中显著提高了水稻产量。这些发现既为科研人员针对水稻粒形性状的分子调控机制的进一步研究提供了理论基础,也为作物遗传育种的基因改良工作创造了有利的条件。
如今关于水稻粒厚方面的研究报道比较少,而控制水稻粒形的基因是紧密联系、相互作用的,粒厚基因的遗传同样受主效基因的调控,其遗传研究与粒长、粒宽等相比较还不够成熟[102]。利用新开发的基因组编辑技术CRISPR/Cas9在相同的遗传背景下生成已知水稻粒形基因的突变体,将有助于建立控制水稻籽粒大小的遗传调控网络[101]。值得注意的是,相同的等位基因突变可能会导致不同遗传背景下水稻的不同表型,研究为何相同的突变在不同品种上有不同的效果,将有助于育种工作者合理利用等位基因突变来提高特定遗传背景下的水稻产量。
猜你喜欢
环球时报(2022-09-20)2022-09-20
现代畜牧科技(2021年4期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
今日农业(2020年24期)2020-12-15
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
河北农业科学(2018年2期)2018-07-26
商洛学院学报(2017年2期)2017-05-17
兽医导刊(2016年12期)2016-05-17
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
