
ERH 基因对人膀胱癌细胞周期的影响
2021-01-06 08:44周荣升韩从辉史振铎张治国3庞
中国医药导报 2020年33期
周荣升 韩从辉,3 陈 波 郝 林,3 史振铎 姜 波 张治国,3庞 昆,3▲
1.江苏省徐州市中心医院 南京中医药大学附属徐州中心医院 东南大学医学院附属徐州医院徐州医科大学徐州临床学院泌尿外科,江苏徐州 221009;2.江苏师范大学生命科学学院,江苏徐州 221000;3.苏州大学附属第三临床学院泌尿外科,江苏常州 213000
在世界范围内,膀胱尿路上皮癌(BUC)在恶性肿瘤中发病率排第五位[1]。在2018 年美国恶性肿瘤统计中,BUC 的发病率高居第四位,占恶性肿瘤总发病率的7%[2-3],在男性泌尿生殖系统中,仅次于前列腺癌,高居第二位,每年约有70 000 例新病例,并导致约15 000 人死亡[4]。在中国,BUC 发病率和相关死亡率逐年升高[5-6]。BUC 的主要治疗方式为手术治疗,但BUC 术后3 年内复发率仍高达70%,而且复发者中约1/3 病例向更高分级和分期进展[6-7],其中,浸润性BUC 的10 年生存率仅为50%[8]。化疗和放疗也被广泛用于晚期BUC的治疗,但是这些方法会引起严重的副作用[9]。为有效治疗BUC,减少不良反应,分子靶向治疗引起了临床泌尿学家的关注[10-11]。其具体致病机制尚不清楚,但与增殖基因的过度激活、凋亡因子的活性下降、与转移有关基因的表达增强和炎症因子的激活等[12-13]密切关系。故迫切需要找到BUC 发生发展、进展和转移的潜在机制,继而寻找一个有效的诊断、预后生物标志物和潜在的治疗靶标。
在之前的研究中[14],本课题组通过免疫组化技术和聚合酶链式反应(PCR)技术在蛋白和mRNA 水平方面发现未成熟同源蛋白增强子(ERH)基因在人BUC的T24 细胞系中表达较正常膀胱尿路上皮细胞明显上调,敲低ERH 基因表达后,会显著抑制BUC 的T24细胞和5637 细胞增殖,促进凋亡[15]。本课题组的后续实验表明[16],敲除ERH 基因后,人BUC 细胞在裸鼠的皮下成瘤能力受到显著抑制。
为了进一步研究ERH 基因调控BUC 增殖凋亡的分子生物学机制,本研究通过人转录组表达谱芯片技术,检测了敲除ERH 基因后,BUC 细胞基因的表达变化;通过对表达差异基因进行生物信息学分析,发现ERH 基因可能调控细胞周期相关蛋白;ERH 基因通过对细胞周期的调控,影响BUC T24 细胞的增殖和凋亡功能。现报道如下:
1 材料与方法
1.1 伦理、细胞培养及分组
所有实验均经江苏省徐州市中心医院伦理委员会批准并通过。膀胱癌细胞系T24 购自中国科学院上海生命科学研究院细胞资源中心。将细胞在含有10%FBS、链霉素和青霉素的RPMI 1640 培养基中于37℃、含有5%CO2的培养箱中培养。根据是否使用慢病毒敲除ERH 基因,将人膀胱癌T24 细胞分为ERH 敲除组和ERH 正常组,慢病毒敲除方法如前期文献所述[4,6]。
1.2 主要仪器和试剂
RNA 质检采用NanoDrop 2000 仪(美国Thremo 公司,型号:OD1000+)、Agilent RNA 6000 Nano Kit 试剂盒(货号:5067-1511)和Agilent 2100 Bioanalyzer(美国Agilent 公司)分析仪(部件号:G2939BA);体外反转录和aRNA 纯化采用GeneChip 3’ IVT Express Kit试剂盒(美国Affymetrix 公司,货号:902790);芯片杂交和洗染的试剂盒为GeneChip Hybridization Wash and Stain Kit 试剂盒(美国Affymetrix 公司,货号:900 720);芯片杂交采用GeneChip Hybirdization Oven 645仪、芯片洗染采用GeneChip Fluidics Station 450 仪,芯片扫描采用GeneChip Scanner 3000 仪 (三台仪器均购自美国Thremo 公司);流式细胞仪为Guava easyCyte HT(美国Millipore 公司);荧光显微镜为日本Olympus公司的IX71。
1.3 人转录组表达谱芯片
通过人转录组表达谱芯片技术对ERH 敲除组和ERH 正常组细胞进行分析比较,每组设3 个样本,共6 个样本,使用RNA marker 作为梯子(参考样本)。质检衡量标准为RIN(RNA Integrity Number,RNA 完整指数)值,这是检测RNA 质量的质控方法,RIN 数值大小即反应样品完整性情况,数值越接近10 表明样品完整性越高,反之RIN 值越小完整性越差。Agilent 2100 Bioanalyzer RNA 质检合格的标志是RIN(RNA完整指数)≥7.0 且28S/18S>0.7[17]。
总RNA 样本经Agilent 2000 分析后,制备成aRNA,然后经一链合成得到cDNA,然后二链合成双链DNA模板,体外反转获得带生物标记的aRNA,纯化后将其片段化,然后与芯片探针杂交,杂交完成后,对芯片进行洗染,扫描得到图片和原始数据。检测全转录组表达谱中受ERH 敲除影响的基因表达情况。
反应条件:一链cDNA 合成,42℃,2 h 后,4℃保持;双链cDNA 合成,16℃,1 h,65℃,10 min 后,4℃保持;反转录,40℃,16 h 后,4℃保持;片段化,94℃,35 min后,4℃保持;芯片杂交,98℃,10 min,45℃,3 min 后,4℃保持。
1.4 差异基因富集分析和筛选
使用Metascape 数据库(http://metascape.org/gp/index.html)对筛选出来的差异基因进行功能富集分析。Metascape 数据库可以根据差异基因列表,根据其功能进行分析、注释和富集。采用基因功能(gene oncology)和KEGG 通路(KEGG pathway)数据库进行分析。然后根据基因功能做图,并根据BioGrid6,In-Web_IM7,OmniPath8 数据库对相应的蛋白互作进行分析,绘制蛋白互作网络图。
1.5 细胞周期功能研究
使用流式细胞技术观察ERH 基因敲除对人膀胱癌T24 细胞周期的影响。使用6 孔板,细胞生长至覆盖率约80%的时候通过胰酶消化,重悬,细胞计数将数目控制在106个/孔,共设置3 个复孔,洗涤3 次,后乙醇固定,使用PI 细胞染色,然后上机检测,采用ModFit 软件进行分析数据。
1.6 统计学方法
采用SAS 8.02 统计学软件对所得数据进行分析,计量资料以均数±标准差()表示,采用student’s t 检验;计数资料采用例数或百分比表示。以P <0.05为差异有统计学意义。
2 结果
2.1 RNA 质检
ERH 正常组三个样本的RIN 值分别是8.9、9.5、9.5,28S/18S 值分别是1.9、1.9、1.8,质检合格;ERH 敲除组三个样本的RIN 值均为9.4,28S/18S 值分别是1.8、1.9、1.8,质检合格。见图1。

图1 Agilent 2100 Bioanalyzer RNA 质检图谱
2.2 ERH 敲除后的其余基因表达变化情况
人转录组表达谱芯片结果显示,ERH 基因在ERH 敲除组表达明显降低,FC 值为-4.50,P <0.0001。ERH 敲除组比ERH 正常组有344 个基因表达上调,254 个基因表达下调(截断值|FC|>2)。见图2。

图2 ERH 正常组和ERH 敲除组基因表达散点图
2.3 ERH 敲除后的差异基因及互作网络
通过Metascape 数据库对筛选出的598 个基因进行生物功能富集分析。结果显示,有显著差异的598 个基因,与肿瘤相关的通路有:细胞因子介导的信号通路、NOD 样受体信号通路、肿瘤坏死因子信号通路、嗜同性细胞黏附、P53 下调通路、负向调节蛋白质磷酸化、负向调节细胞增殖、细胞凋亡通路等。见表1、图3。图3 为log10(P)值由小到大的前20 位差异基因对应的生物学功能。
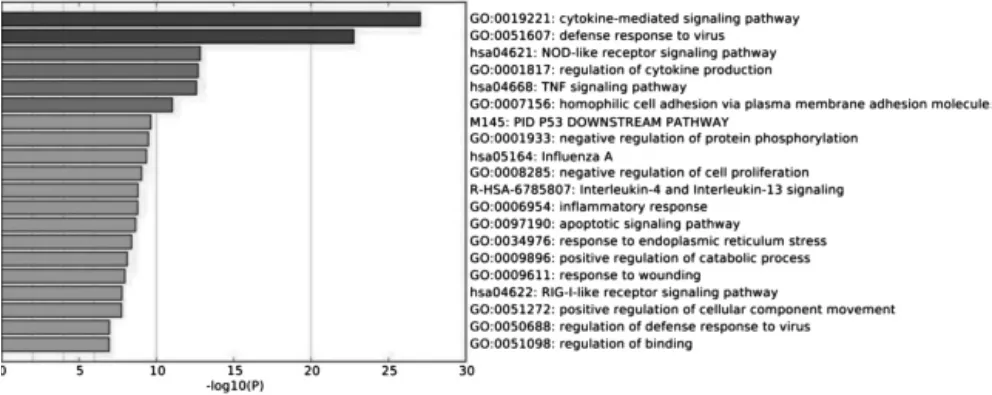
图3 差异基因生物功能富集热图
数据库进一步对筛选的598 个基因进行互作功能研究,使用Cytoscape5 可视化网络图,每个节点代表一个生物功能,类似的生物功能聚为一个集群,用相同颜色表示。见图4(封四)。

2.4 ERH 基因敲除后,差异基因对应蛋白的互作网络与功能分析
继续采用Metascape 数据库链接BioGrid6,In-Web_IM7,OmniPath8 数据库对相应的部分蛋白互作关系做出分析并绘制互作网络图(图5)。发现ERH敲除后的598 个基因中,有大量差异基因对应的蛋白与细胞周期密切相关。

表1 差异基因对应的肿瘤相关生物功能

图5 差异基因相关蛋白互作网络图
2.5 ERH 基因敲除对细胞周期的影响
ERH 敲除组G1期和G2/M 期的细胞数目少于ERH 正常组,ERH 敲除组S 期细胞数目多于ERH正常组,差异均有统计学意义(均P <0.05)。见表2、图6。
3 讨论
ERH 基因位于人类14 号染色体,优先表达于细胞核[18],除了核仁外,也表达于细胞质中,作为基因增强子和转录调控因子,具有多种结合蛋白,如RPS3或RPL19[19],功能涉及嘧啶生物合成、细胞周期、转录和细胞生长控制过程。有研究表明[20],ERH 蛋白通过与锌指蛋白Ciz1 结合,成为与p21cip1/Waf1相互作用的DNA 复制因子。ERH 蛋白和RNA 结合蛋白Mmi1 形成一个复合物,称为ERH-Mmi1 复合物(EMC)[21],与剪接体蛋白SNRPD3 相互作用,是有丝分裂运动蛋白CENP-E 的mRNA 剪接所必需的[22]。这些研究均提示ERH 基因可能会参与癌细胞的细胞周期[23-24]。在前期研究中,本课题组首次发现ERH 基因在BUC T24 细胞中呈高表达丰度,而且ERH 基因敲除对T24 细胞的增殖、克隆形成具有抑制作用,对凋亡具有促进作用[15,25]。本课题组进一步在动物体内发现ERH 基因敲除后,裸鼠皮下成瘤能力明显减弱[16]。
表2 ERH 基因敲除后细胞各周期变化(%,)

表2 ERH 基因敲除后细胞各周期变化(%,)
注:ERH:未成熟同源蛋白增强子

图6 ERH 基因敲除后细胞各周期变化
本研究将ERH 基因敲除后,使用流式细胞技术观察ERH 基因敲除对细胞周期的影响。结果可见,ERH 敲除组G1期和G2/M 期的细胞数目少于ERH 正常组,ERH 敲除组S 期细胞数目多于ERH 正常组,差异均有统计学意义(均P <0.05),说明ERH 基因主要影响细胞周期的S 期。本研究进一步通过人转录组表达谱芯片技术,在抑制ERH 基因的表达后,检测转录组基因表达情况。结果显示,有598 个基因表达受ERH 基因敲除的影响发生上调或下调。然后,通过生物信息分析技术,筛选与肿瘤通路有关的差异基因,发现ERH 基因敲除与细胞因子介导的信号通路、NOD 样受体信号通路、肿瘤坏死因子信号通路、嗜同性细胞黏附、P53 下调通路、负向调节蛋白质磷酸化、负向调节细胞增殖、细胞凋亡通路等有关。这些通路介导了BUC T24 细胞的增殖和凋亡过程,这与本课题组前期研究相一致[16]。然后,本研究继续通过生物信息分析技术来寻求蛋白的互作网络与功能分析,结果提示ERH 基因可能通过细胞周期蛋白影响BUC T24 细胞的增殖与凋亡,这也与本课题组前期研究结果一致[16]。
在BUC T24 细胞系中,本研究通过细胞周期实验发现ERH 基因主要影响细胞周期的S 期,通过人转录组表达谱芯片技术,找到了ERH 基因敲除相关的差异基因598 个。通过生物信息分析技术,发现了ERH 基因通过调控细胞周期影响BUC T24 细胞的增殖和凋亡。
猜你喜欢
心电与循环(2020年1期)2020-02-27
国际呼吸杂志(2019年4期)2019-03-12
江苏农业科学(2017年5期)2017-04-15
发明与创新(2016年23期)2016-10-13
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
湖北工业大学学报(2016年5期)2016-02-27
医学研究杂志(2015年5期)2015-06-10
湖北农业科学(2014年3期)2014-07-21
河南科技(2014年10期)2014-02-27
