
枯萎期芦苇收割时间对湿地脱氮效果及根系呼吸代谢的影响
2021-08-23 10:23王俊力付子轼乔红霞陈桂发刘福兴
环境科学研究 2021年8期
王俊力, 付子轼, 乔红霞, 陈桂发, 刘福兴*
1.上海市农业科学院, 上海 201403 2.上海低碳农业工程技术研究中心, 上海 201415
氮(N)是生态系统中构成生物体的重要元素,但也是引起水体富营养化的主要元素之一[1]. 自工业革命以来,全球范围内人为源氮排放从1860年的15×1012g/a增至20世纪90年代初的156×1012g/a,预计到2050年将达到267×1012g/a[2]. 我国农业生产使用的氮肥中有25%的氮随降雨径流和渗漏进入水环境,是导致水体氮浓度升高和污染加重的重要原因[3]. 湿地作为陆地与水体之间的过渡带,对拦截陆地方向的氮污染源、降低水体中的氮水平具有重要作用[4].
湿地通过复杂的物理、化学和生物等方面的作用实现氮去除,方式包括挥发、氨化、硝化/反硝化、植物吸收、基质吸附等[5],其中生物(植物和微生物)起到关键作用[6]. 研究[7-8]表明,超过50%的氮通过微生物硝化/反硝化作用去除,而植物会影响湿地微生物群落结构的时空变化. 芦苇(Phragmitesaustralis)是湿地生态系统中的主要植物之一,常见于河口、湖滨、海岸等地,在全球广泛分布[9]. 在我国的亚热带季风气候区,分布有大面积芦苇湿地,该地区积温高,无霜期长,芦苇通常在3月份出芽,到11月份进入枯萎期,冬季根系休眠时间较短[10]. 研究表明,生长期芦苇湿地的脱氮效率为50%~90%[11-12],冬季则降至20%左右[13]. 虽然冬季芦苇地上部分枯萎,但其根系仍存在活力[14],了解冬季芦苇根系活动,对如何采取有效措施促进冬季湿地脱氮效果具有重要意义.
收割是湿地植物最主要的管理措施之一. 芦苇一年收割一次能够去除地上部枯死量,有效恢复其活力[15]. 在冬季开始时,湿地植物地上部分的氮逐渐向地下转移,如果不收割,植物吸收的氮可能会随着植株的枯萎腐烂重新回到湿地系统中,出现水体氮浓度升高现象[16]. 大型芦苇湿地系统中,收割措施具有局限性[17],因为其需要更多的人力、物力和财力投入. 因此,选择最佳收割时间,是影响冬季芦苇湿地脱氮效果的重要因素.
呼吸代谢的持续进行是植物休眠过程的显著特点[18],在亚热带地区,冬季芦苇进入休眠,但其根系仍存在活力,冬季芦苇根系呼吸代谢的变化可能会导致湿地脱氮过程发生变化. 枯萎期不同时间收割是否会影响根系的呼吸代谢特征,从而影响湿地脱氮效果,有关这方面的研究还鲜见报道. 该研究系统探讨了枯萎期不同收割时间情况下,芦苇根系活力和呼吸代谢途径关键酶活性,及其与湿地脱氮效果的关系,以期探讨枯萎期收割影响植物根系呼吸的作用机制,为人工湿地的管理和功能发挥、富营养化水体的治理提供理论依据.
1 材料与方法
1.1 研究区概况
试验研究位于上海市农业科学院庄行综合试验站(30°53′N、121°23′E). 该区属亚热带季风气候,多年平均降水量 1 191.5 mm,蒸发量 1 236.8 mm,年均温度16.1 ℃,全年日照时数 1 900.2 h,无霜期224.4 d.
1.2 试验设计
试验采取模拟芦苇湿地的方式进行,构建12个相同结构的传统水平潜流湿地系统(见图1),长×宽×深为150 cm×50 cm×80 cm,进水端及出水端各设置长度为20 cm的布水区和集水区,并设有出水管,主体填料区域长度为110 cm. 进出水均采用穿孔板,出水孔与出水管底标高一致. 为了便于测定湿地内部的水质参数,在系统中间沿程竖向设置内径5 cm穿孔管. 内部填充粒径约1~2 cm的砾石基质,基质层深70 cm,运行时水在介质表面以下流动,水深为60 cm.
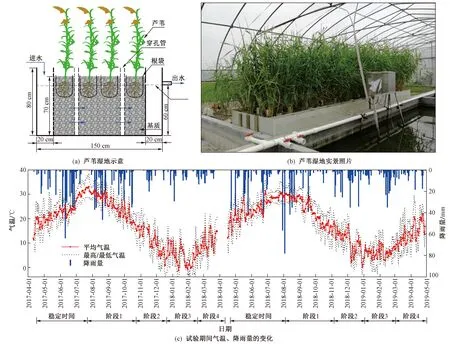
注: 试验第1年(2017年7月—2018年3月)芦苇于2017-04-10移栽,湿地稳定至2017-07-14正式开始试验,阶段1为正式试验至2017-10-13枯萎前收割,阶段2为枯萎前收割后至2017-12-12枯萎中期收割,阶段3为枯萎中期收割后至2018-02-08枯萎末期收割,阶段4为枯萎末期收割后至2018-03-27结束试验;试验第2年(2018年8月—2019年4月)芦苇于2018-04-16移栽,湿地稳定至2018-08-01正式开始试验,阶段1为正式试验至2018-10-31枯萎前收割,阶段2为枯萎前收割后至2018-12-28枯萎中期收割,阶段3为枯萎中期收割后至2019-02-28枯萎末期收割,阶段4为枯萎末期收割后至2019-04-28结束试验. 下同.图1 芦苇湿地示意、实景照片以及试验期间气温、降雨量的变化Fig.1 Schematic diagram of a single wetland system, picture of the experimental site, and fluctuation of air temperature and daily precipitation during the experiment
植物采用当地常见芦苇(Phragmitesaustralis),选取长势相近的芦苇幼苗移栽至根袋(300目尼龙网袋,20 cm×30 cm)中,袋内填充基质与袋外相同[19],系统内的初始种植密度为2株/袋(约29株/m2),芦苇在系统中培养约2个月,待长势平稳后开始进水,2年试验时间轴如图1所示(第1年为2017年7月—2018年3月,第2年为2018年8月—2019年4月). 进水利用河道水配置,用尿素调节进水ρ(TN),利用恒流泵进行连续进水,水力负荷(HLR)为100 mm/d,水力停留时间(HRT)为2.4 d. 系统运行约1个月趋于稳定后,正式开始试验.
试验设置4个处理,即W1(对照,芦苇不收割)、W2〔枯萎前(10月)收割〕、W3〔枯萎中期(12月)收割〕、W4〔枯萎末期(翌年2月)收割〕,每个处理3次重复. 收割方式为去除湿地系统表面10 cm以上的植物部分,并定期对凋落物进行清理.
试验在自然环境中进行,第1年试验结束后,重新布置第2年试验,2年的进水ρ(TN)不同,第1年较高〔相当于小麦季地表径流中较高ρ(TN)〕,第2年较低〔相当于小麦季地表径流中较低ρ(TN)〕[20],进水参数如表1所示. 以收割为节点将每年的试验分为4个阶段:阶段1为试验开始至W2收割前; 阶段2为W2收割后至W3收割前; 阶段3为W3收割后至W4收割前; 阶段4为W4收割后至结束试验.

表1 试验期间进水水质情况
1.3 取样与测定
水体取样进出水同时进行,进水在布水区采集(3个重复),出水在每个湿地系统的出水口采集,每月一次(阶段1),当收割开始后,每月2次(阶段2、3、4),并现场测定系统中水体的pH、ORP、EC、ρ(DO)、温度(T)等指标.ρ(TN)采用GB 11894—1989《水质 总氮的测定 碱性过硫酸钾消解紫外分光光度法》测定,ρ(NH4+-N)、ρ(NO3--N)和ρ(NO2--N)使用流动分析仪(AA3,Seal,德国)测定.
根系取样在每次收割时和试验结束时进行,方法是小心将根袋从系统中拿出,为了弥补系统取样后的植物损失,补充另外培养的对应处理芦苇维持系统平衡. 根系活力采用氯化三苯基四氮唑(TTC)法测定. 己糖激酶(HK)、丙酮酸激酶(PK)、磷酸果糖激酶(PFK)、异柠檬酸脱氢酶(ICDH)、细胞色素氧化酶(COX)等根系呼吸代谢关键酶活性采用紫外可见分光光度计(DR 5000,Hach,美国)进行测定,且以μmol/min(以每mg蛋白质计)表示.
1.4 数据处理
利用IBM SPSS 22.0软件对数据进行统计分析;采用单因素方差分析(one way ANOVA)检验不同收割处理对各参数的影响,显著性检验采用Duncan在0.05显著性水平下进行;用SigmaPlot 12.5软件作图,图表中数据为平均值±标准差;利用Canoco 5.0软件进行冗余分析.
2 结果与分析
2.1 枯萎期芦苇收割对湿地脱氮效果的影响
从芦苇湿地氮去除效果(见图2)可以看出,2年间TN和NH4+-N去除率的变化趋势相似,与第1年相比,第2年TN和NH4+-N去除率更高,其中第1年W1处理的TN和NH4+-N平均去除率分别为51.2%±24.5%和59.0%±21.9%,第2年分别为64.5%±25.8%和71.8%±30.5%.
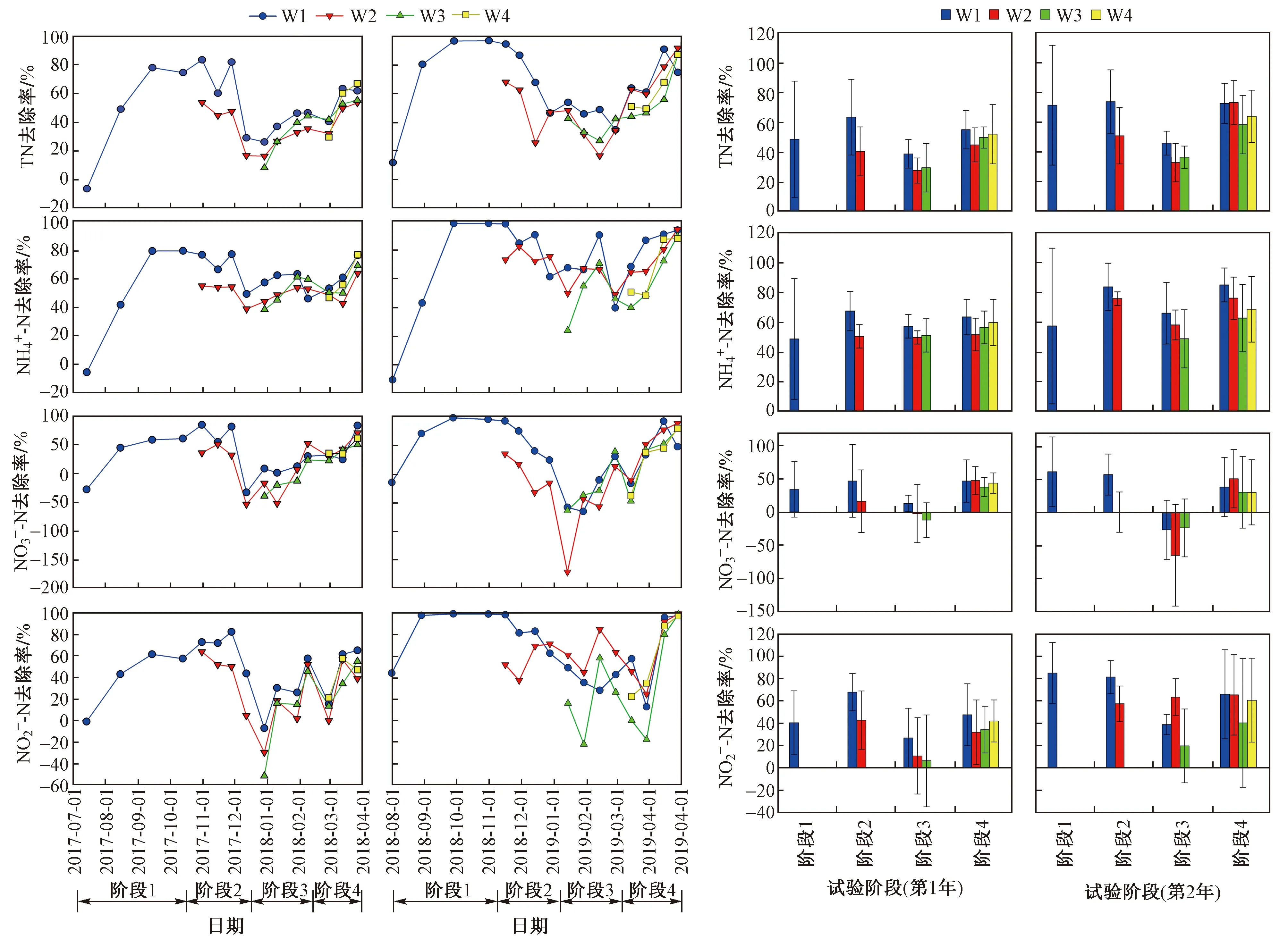
图2 不同收割时间下芦苇湿地TN、NH4+-N、NO3--N和NO2--N去除率的变化Fig.2 Removal of TN, NH4+-N, NO3--N and NO2--N from the reed wetlands under different harvesting time
从不同处理上看,与收割处理相比,不收割处理的TN和NH4+-N去除率较高. 每次收割后,TN和NH4+-N去除率均为下降趋势,W2处理使阶段2的TN和NH4+-N去除率在第1年分别下降了23.0%和17.0%,第2年分别下降了22.5%和7.5%,随着时间的延长,去除率降低趋势逐渐减小,但始终低于W1处理. W3处理TN和NH4+-N去除率的变化趋势与W2处理相似,阶段3的TN和NH4+-N去除率在第1年分别下降了24.0%和10.8%,第2年分别下降了18.2%和25.2%. 与W2和W3处理相比,W4处理TN和NH4+-N去除率的下降趋势较小,阶段4的TN和NH4+-N去除率在第1年分别下降了3.0%和3.7%,第2年分别下降了8.0%和16.0%.
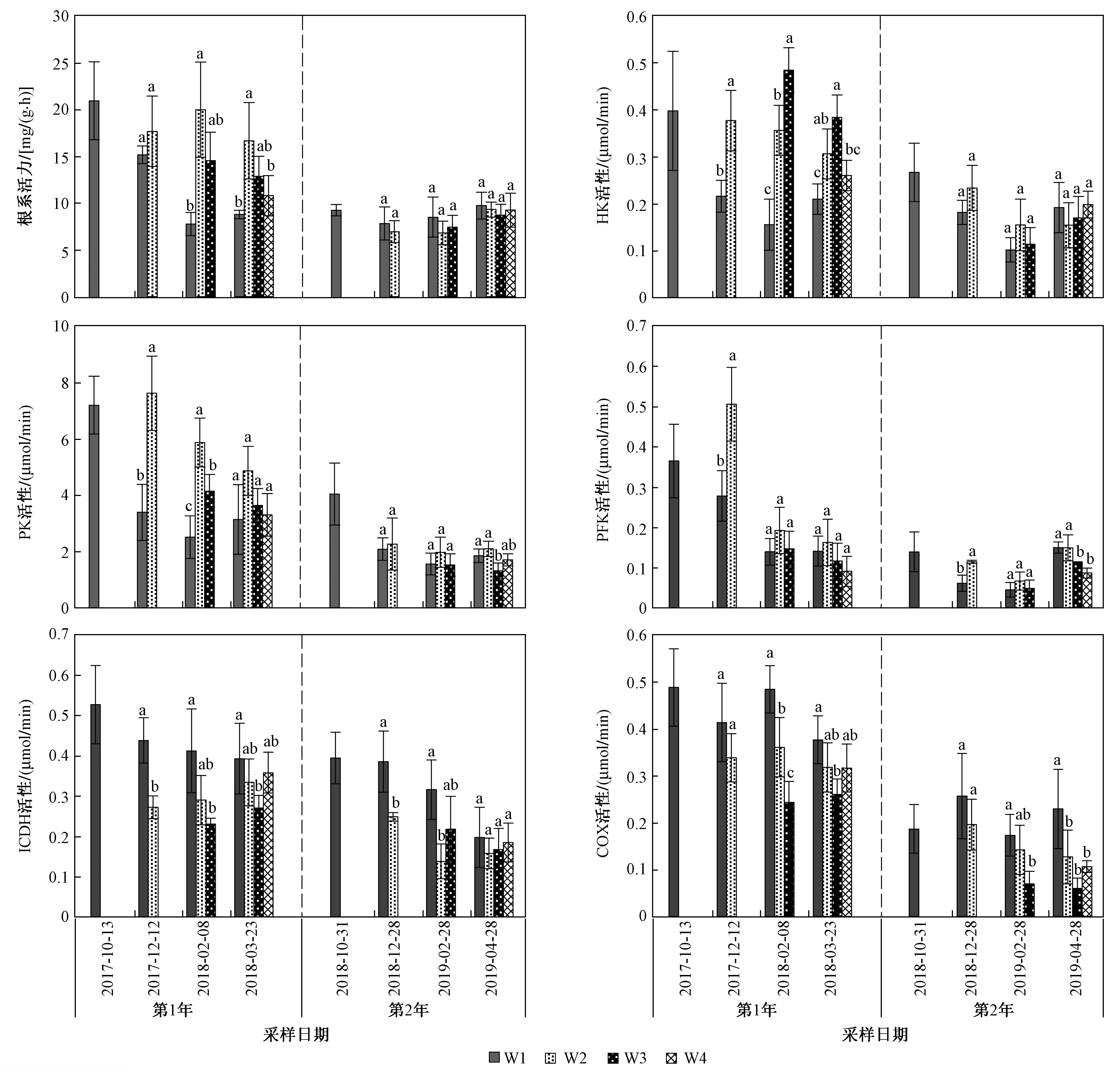
注: 不同小写字母表示相同采样日期不同处理下各参数差异显著(P<0.05).图3 不同采样日期不同处理芦苇根系活力和呼吸代谢关键酶活性Fig.3 Reed root activity and key enzyme activity of respiratory metabolism under different treatment on different sampling dates
2年试验中阶段3的氮(TN、NH4+-N、NO3--N、NO2--N)去除率均最低,且NO3--N和NO2--N去除率波动均较大,甚至有负值出现. 与TN和NH4+-N去除率趋势一致,每次收割后,NO3--N和NO2--N去除率基本为下降趋势,且W4处理的去除率下降趋势最小.
2.2 枯萎期芦苇收割对根系呼吸代谢的影响
年份和处理对根系活力和根系呼吸代谢关键酶HK、PK、PFK、ICDH、COX活性均有显著影响(P<0.05);年份与处理的交互作用对根系活力以及HK、PK活性有显著影响(P<0.05),但对PFK、ICDH、COX活性无显著影响(P>0.05,见图3).
试验期间2年同期相比,第1年的芦苇根系活力和呼吸代谢关键酶活性均明显高于第2年(P<0.05). 第1年,与不收割相比,收割处理均增加了芦苇根系活力以及HK、PK活性,且W2和W3处理增加的更为明显(P<0.05);第2年,处理间根系活力以及HK、PK活性基本无显著差异. 2年试验中,W2处理均显著增加了刚收割后的根系PFK活性(P<0.05),而随着试验时间的延长,差异逐渐不明显,试验第2年W4处理显著降低了PFK活性(P<0.05). 与W1相比,收割处理均降低了根系ICDH和COX活性,且W3处理对第1年的ICDH活性和2年的根系COX活性降低的更为明显(P<0.05);第2年,W2处理对根系ICDH活性降低的更显著(P<0.05).
2.3 湿地脱氮效果与芦苇根系呼吸代谢的关系
用RDA分析湿地脱氮效果与芦苇根系呼吸代谢的相互关系,以根系活力和呼吸代谢关键酶活性为环境变量,解释湿地氮去除率的变化. 结果(见表2)显示,试验第1年,轴1和轴2的特征值分别为0.847和0.069,去除率与呼吸代谢关系累计解释量为96.8%;试验第2年,轴1和轴2的特征值分别为0.696和0.061,去除率与呼吸代谢关系累计解释量为93.4%,说明轴1和轴2能较好地反应二者之间的关系,且主要由轴1决定.
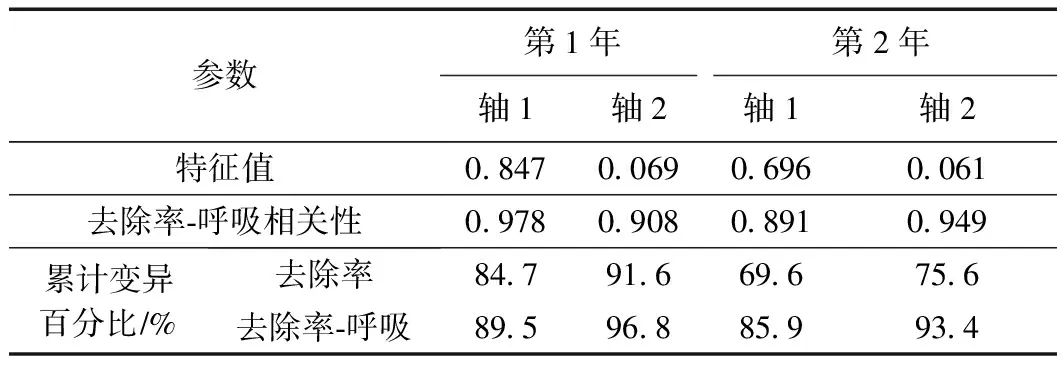
表2 湿地氮去除率与芦苇根系呼吸代谢关系的RDA结果
由图4可见,试验第1年,湿地氮去除率的变化整体上由ICDH和HK活性主导,其与ICDH活性呈正相关,与HK活性呈负相关. 试验第2年,湿地氮去除率的变化由根系呼吸代谢指标(HK、PK、PFK、ICDH、RA、COX活性)主导,均呈正相关,且HK活性对湿地氮去除率的影响最大.
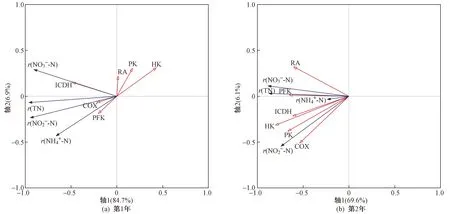
注: r(TN)表示TN去除率; r(NH4+-N)表示NH4+-N去除率; r(NO3--N)表示NO3--N去除率; r(NO2--N)表示NO2--N去除率; RA表示根系活力; HK表示己糖激酶活性; PK表示丙酮酸激酶活性; PFK表示磷酸果糖激酶活性; ICDH表示异柠檬酸脱氢酶活性; COX表示细胞色素氧化酶活性.图4 湿地氮去除率与芦苇根系呼吸代谢关系的RDA分析Fig.4 RDA analysis of wetland nitrogen removal and reed root respiratory metabolism
3 讨论
3.1 不同收割时间对湿地脱氮效果的影响
温度影响湿地的脱氮效率[21],该研究中,阶段3的湿地脱氮效率最低,可能与低温条件抑制生物过程有关[22],这与Hu等[23]的研究结果一致,其证明了冬季(12月—翌年2月)湿地出水ρ(NO3--N)增加,反硝化作用受到抑制. 该研究中,试验第1年的进水ρ(TN)高于第2年,而第2年的TN和NH4+-N去除率高于第1年,这与Liu等[24]的研究结果不同,其表明进水氮浓度增加,TN和NH4+-N去除率显著提高. 湿地的脱氮效率不是绝对的,受到各种因素的影响,如环境因子(pH、温度、ORP等)、系统配置、水力状况、进水模式、植物种类及植物收割方式等[25-27].
收割是湿地植物的主要管理方式[14],应优化收割时间,以提高湿地在冬季的脱氮效率. 该研究中,2个试验年之间表现出相同的脱氮趋势,均为不收割处理的TN和NH4+-N去除率最高,这与Shuai等[28]的研究结果不同,其表明与不收割处理相比,收割处理提高了湿地脱氮效率. 该试验过程中定期对凋落物进行清理,避免芦苇中的氮通过腐烂返回湿地中,更有利于解释收割时间对湿地脱氮效果的影响. 研究[29]表明,植物地上部收割会影响反硝化微生物生长的根际微环境,从而影响湿地脱氮过程. 该研究结果表明,收割时间越晚,对湿地脱氮效果的影响越小,与不收割处理相比,枯萎末期收割的TN和NH4+-N去除率下降趋势较小,可能是因为不收割芦苇可以为湿地保温,促进了冬季的反硝化作用,而枯萎末期收割后,气候逐渐变暖,为植物的生长提供了有利条件[30],从而有利于湿地系统恢复脱氮效果. 另外,植物根系的泌氧作用会影响微生物的硝化-反硝化作用,从而间接影响湿地的脱氮效率[31]. 枯萎末期收割保持了芦苇在冬季的完整性,这可能会保护植物根系,从而持续为根际环境供给氧气[32],减少了对湿地脱氮效率的影响.
3.2 不同收割时间对根系呼吸代谢的影响
氮是植物生长发育必需营养元素,环境中的氮含量与植物的生理生化过程有密切关系,对维持植物呼吸的作用较为显著[33]. 根系呼吸作用是植株地下部代谢的重要过程,它对养分的吸收、根系更新以及植株生长发育具有重要意义,极易受到外界条件变化的影响[34]. 收割作为湿地人工干预最常见的方式,可能会引起收割后植物根系呼吸代谢的变化,这也是植物适应干扰的方式. 根系活力的大小反映根系代谢能力强弱,研究[35]表明,根系对外界产生应激反应的表现是根系活力先快速提高,通过加强代谢来缓解干扰伤害,促进养分和水分的吸收,随着胁迫时间延长,植物本身储存物质消耗加剧,根系活力又降至对照水平. 该研究发现,试验第1年,收割处理提高了芦苇根系活力,且越早收割越能增强芦苇冬季的根系活力,枯萎末期收割则对根系活力影响较小;这可能与根系的应激反应有关,早期的收割处理对芦苇的干扰可能较大,植物为获得足够的氮而做出补偿性反应[36],使得根系活力增加;晚期收割时,由于植物本身处于休眠期结束开始生长阶段,收割处理对其干扰较小,根系活力与不收割相比差异不显著. 试验第2年,收割处理对根系活力无显著影响,可能是在低进水ρ(TN)条件下,植物根系活力显著低于高进水ρ(TN)条件(见图3),收割干扰引起的应激反应与不收割相比差异不显著,说明根系活力不仅受收割时间的影响,还与环境中氮浓度有关.
植物呼吸代谢中的糖酵解(EMP)途径是有氧呼吸和无氧呼吸的共同途径,HK、PK、PKF是调控EMP途径的关键酶. 一般情况下,植物呼吸代谢以EMP途径进入三羧酸(TCA)循环为主,TCA循环是植物维持生命活性所需的主要能量来源,ICDH是调控TCA循环的关键酶. 细胞色素(CP)途径是末端氧化电子传递链的主要途径,COX是链上的终端酶[34]. 该研究中,收割处理使根系呼吸代谢关键酶活性变化趋势在2年试验之间有显著差异,说明根系呼吸代谢的变化与进水ρ(TN)高低有关,环境中的氮对根系呼吸的调控作用甚为重要,根系呼吸在进水ρ(TN)高的情况下较高,这与陈贵等[37]研究结果一致. 收割处理下HK、PK和PFK活性的变化规律基本一致,试验第1年,枯萎前期收割均显著增加了HK、PK和PFK活性,可能是收割时根系活力较强,干扰后植物为获得足够的氮而做出补偿性反应[36],EMP途径加快,而枯萎末期根系活力较弱,收割处理与不收割处理相比差异不显著. 但是冬季植物呼吸代谢并非越高越好,有研究[38]表明,冬季植物加强维持呼吸,会导致植物碳储量耗尽,减少翌年春季新根生长能力和植物的生产力,抵消春季温暖给植物带来的积极影响. 试验第2年,枯萎中期和末期收割均会降低PK、PFK活性,EMP途径也有一定程度的受阻. 该研究中,2年试验的收割处理均降低了ICDH和COX活性,且与枯萎末期收割相比,早期收割降低的更为明显,说明收割会影响根系呼吸中的TCA循环和CP途径,且末期收割的影响程度相对较小.
3.3 湿地脱氮效果与根系呼吸代谢的关系
湿地污染物净化效果与植物的种类及生理特性关系密切[39]. 研究表明,根系呼吸最小值出现在12月—翌年3月[40],而该研究结果显示该时间段湿地脱氮效率同样最低. 通过RDA分析表明,根系活力和呼吸代谢关键酶活性与湿地氮去除率之间密切相关(第1年解释量为96.8%,第2年解释量为93.4%),说明冬季芦苇根系呼吸代谢机制是其适应收割干扰的重要策略,与湿地脱氮效果存在密切关系. 试验第1年,湿地氮去除率的变化整体上由ICDH和HK主导,其与ICDH活性呈正相关,与HK活性呈负相关. ICDH是连接C-N代谢酶的关键,是TCA循环中最重要和最具有调控意义的限速酶[41]. ICDH活性增加会提高根系呼吸和根系活力,从而增加根系泌氧促进微生物的硝化-反硝化过程[31],提高湿地的脱氮效率. HK催化EMP途径的第一步反应,将葡萄糖磷酸化,这有利于它进一步参与合成和分解代谢[34]. HK活性对EMP途径的影响最大,HK活性降低会使EMP途径贡献率降低,而湿地氮去除率增加,这与第2年试验结果(HK活性与湿地氮去除率呈正相关)正好相反,可能是根系呼吸代谢途径对环境氮浓度高低的响应不同,从而影响湿地脱氮效果. 该研究中采取的试验方式是将湿地中发生的生物和非生物反应整合起来评估湿地脱氮效果,以探索最佳收割时间,根系相关的或其诱导机制还需要更加深入的研究,以了解各反应机制在冬季湿地脱氮过程中的贡献.
4 结论
a) 枯萎期不同时间收割影响芦苇湿地的脱氮效果和根系的呼吸代谢特征. 与不收割处理相比,收割处理会降低湿地的脱氮效果,但枯萎末期收割对湿地脱氮效果的影响较小.
b) 收割时间对芦苇根系活力和呼吸代谢关键酶的影响与进水ρ(TN)高低有关,在进水ρ(TN)较高的年份,枯萎前收割显著增强了芦苇的根系活力以及HK、PK、PFK活性;在进水ρ(TN)较低的年份,收割处理对根系活力以及HK、PK、PFK活性的影响不大,而收割处理均降低了2个年份的根系ICDH和COX活性,但枯萎末期收割对其影响程度相对较小.
c) 收割时间对冬季芦苇湿地脱氮效果的影响与根系呼吸代谢特征存在一定的相关性,进水ρ(TN)较高的年份,湿地氮去除率的变化整体上由ICDH和HK活性主导;进水ρ(TN)较低的年份,湿地氮去除率的变化由根系呼吸代谢指标(HK、PK、PFK、ICDH、RA、COX活性)主导,均呈正相关,且HK活性对湿地氮去除率的影响最大.
猜你喜欢
现代园艺(2022年7期)2022-11-19
中国资源综合利用(2022年9期)2022-10-13
林业科技(2022年5期)2022-10-08
海外文摘·文学版(2022年4期)2022-04-14
现代矿业(2022年3期)2022-04-09
黄河之声(2021年19期)2021-02-24
魅力中国(2020年23期)2020-12-08
湖北农业科学(2019年22期)2019-12-23
岁月(2018年2期)2018-02-28
中国诗歌(2017年12期)2017-11-15
