
有机固废堆肥中产臭及除臭技术的微生物作用机制研究进展
2021-10-27 13:56吴伟霞席北斗黄彩红李艳红马彩云唐朱睿
环境科学研究 2021年10期
吴伟霞, 席北斗, 黄彩红, 李艳红, 李 伟, 马彩云,4, 郭 威, 唐朱睿
1.桂林理工大学环境科学与工程学院, 广西 桂林 541006
2.中国环境科学研究院, 环境基准与风险评估国家重点实验室, 北京 100012
3.中国环境科学研究院, 国家环境保护化学品生态效应与风险评估重点实验室, 北京 100012
4.兰州交通大学环境与市政工程学院, 甘肃 兰州 730070
有机固废主要包括农林废弃物、畜禽粪便、餐厨垃圾和污水剩余污泥等,年产量大,若得不到有效处理,其产生的渗沥液、恶臭气体和温室气体等会导致许多环境和公共卫生问题. 全球的城市每年产生约1.3×109t固体废物,预计到2025年将增至每年2.6×109t,而在全球产生的总固体废物中,有机固废占比(46%)最大[1-2]. 2017年我国主要城市固体废物产生量约3.5×108t,垃圾存放引起的“邻避效应”已经成为城市固体废物处理管理者面临的难题之一[3-4]. 据2019年度全国“12369”环保举报情况统计,各类举报中大气污染举报占比(50.8%)居于首位,而大气污染举报中恶臭异味举报占比(41.0%)最高[5]. 采用填埋、焚烧等传统方式对有机固废进行处理,由于环境危害较大,使得其不具有环境可持续性. 有机固废可持续管理的最佳选择是采用资源回收、减量化、再利用和再循环的概念,如堆肥、厌氧消化等生物处理方法[6]. 堆肥是目前最有前途的生物处理方法之一,成本低廉,能处理不同成分的废弃物,并生产出更为稳定的有机肥料,近年来在有机固废的回收利用方面受到了广泛关注[7].
可持续和循环经济协同发展是全世界共同努力的目标,我国也制定了“有机肥代替化肥”等行动计划,倡导可持续发展、循环经济与绿色农业并行,这促使了堆肥技术在我国的推广应用. 堆肥是在微生物作用下有机物经历复杂的代谢反应产生腐殖质的过程,产品可用于土壤改良、增加农作物产量等[8-11]. 尽管堆肥有助于废弃物的管理和消纳,但不可避免地会释放恶臭气体,其中以NH3、H2S等氮硫类气体为主. 堆肥释放的恶臭气体中NH3对气味排放的贡献最大[12]. 虽然H2S等挥发性硫化物(VSCs)的排放量低于NH3,但由于其嗅阈值低,即使低浓度的VSCs也会产生强烈的气味[13-14].
堆肥中物质的转化涉及多种微生物的共同驱动作用. 堆体中有机物被微生物降解的同时也会产生一些带异味的气态代谢产物或中间产物[15]. 微生物的多样性变化和群落演替是堆肥过程的关键特征,其代谢功能取决于微生物群落的组成,而微生物群落的组成主要受温度、pH、通风量等代谢环境的影响,所以任何改变微环境的行为都可以改变微生物作用的方向[16-17]. 微生物在减少恶臭气体排放方面也起着重要作用,许多正在开发或已经运用的生物除臭技术大都依赖于微生物的特性,但堆肥所产生的恶臭气体组分不一且复杂,目前关于恶臭气体生成转化过程中微生物作用机制的研究仍不够透彻,使得微生物治理恶臭气体难以达到同步削减或消除. 因此,对堆肥中产生的恶臭气体进行有效控制仍然十分困难. 该文梳理了堆肥中恶臭气体的主要组成、生物除臭技术,重点阐述了恶臭气体的生物转化机制,包括相关核心微生物、关键功能基因及微环境主控因子等,并提出了基于微生物正向调控实现过程控制的展望,以期为高效精准控制堆肥恶臭气体提供理论参考和技术支撑.
1 臭气的主要组成
有机固废在堆肥过程中会释放组分多样的恶臭气体. 如在牛粪、厨余垃圾及市政污泥中的主要致臭物质分别为NH3、H2S和甲硫醚,H2S、甲硫醇、1,3-二甲基苯、邻二甲苯、二甲基二硫醚和对二甲苯,NH3、甲硫醚和二甲基二硫醚[18-20]. 尽管不同物料堆肥释放的臭气组分不一,但主要可分为含氮和含硫两大类恶臭气体.
含氮类恶臭气体主要包括NH3和含氮有机物. NH3具有刺激性气味,嗅阈值(0.027 mg/m3)相对较高,是有机固废堆肥过程中的主要致臭物质之一. NH3是大气气溶胶的前体,释放到大气中会导致土壤和水体的酸化[21];长期暴露于过量的NH3环境中会对人类健康构成威胁,如中枢神经麻痹和心肌损伤等[22]. NH3挥发是氮素损失的主要途径,占总氮损失的47%~77%[23]. 因此,减少NH3排放是控制堆肥中氮素损失的关键. 堆肥产生的含氮有机物中以胺类、吲哚类和酰胺类为主,当曝气过量时会显著增加含氮有机物的释放[24]. 胺类为腐鱼臭味,与NH3相比胺类对人体危害更大,其毒性随着取代氢原子个数的增加而增加,暴露于较高浓度的三甲胺环境中,可能会引发结膜炎[25-26]. 吲哚类为粪臭味,具有中等毒性,直接接触会危害人体的呼吸系统和皮肤,且对眼睛有严重伤害[27]. 粪臭素为强烈粪臭味,能引起反刍动物的急性肺水肿[28].
挥发性硫化物(VSCs)等含硫类恶臭气体可分为H2S、二硫化碳(CS2)等挥发性无机硫化物和甲硫醇(MT)、甲硫醚(DMS)、二甲基二硫醚(DMDS)等挥发性有机硫化物(VOSCs),具有检测阈值低、气味活性强等特点,已被确定为有机废物堆肥过程中释放的主要气味[29]. 堆肥中会释放出大量的VSCs,据报道,初始总硫中约有55%以VSCs的形式流失[29]. H2S为腐蛋臭味,低浓度的H2S使人有疼痛感,一旦浓度超过2.5×10-4则会导致昏迷甚至死亡,是堆肥过程中产生量最高的VSCs气体,占VSCs总释放量的39%~43%[30-31]. VOSCs在低浓度时是重要的风味成分(如啤酒和奶酪),但在较高浓度下毒性很大,会对人体神经造成损害,且大部分VOSCs都具有腐烛性[32]. 其中,MT嗅阈值(0.7×10-3mg/m3)较低,可作为评估挥发性有机物排放情况的指示气体[33]. 在厨余垃圾堆肥产生的挥发性有机化合物中,MT、DMDS和DMS等的占比为35%~38%[34].
2 臭气生成转化的途径及核心微生物
2.1 臭气生成转化的途径
堆肥中有机底物丰富,无论是好氧还是厌氧环境,伴随着微生物的代谢会经历复杂的物质代谢,均会不同程度地释放恶臭气体. 堆体中氮硫类化合物转化所发生的氨化作用、硝化作用、反硝化作用以及硫酸盐还原作用等是恶臭气体生成转化的主要途径. 堆肥中恶臭气体生成转化的主要途径如图1所示.

图1 堆肥中恶臭气体生成转化的主要途径
NH4+-N是NH3的前体物,也是硝化和同化作用的底物,在堆肥的氮转化过程中起到关键作用[35]. 堆肥初期含水量高,氨基酸态氮等有机氮经氨化作用分解生成的氨主要以NH4+-N存在于堆体中[36]. 高温期由于具有高温及高pH的特性,NH4+-N会转化为NH3释放出堆体外. NH3的产生与NH4+-N密切相关,NH4+-N产量越多,NH3的释放量也越多. Jarvis等[37]研究表明,堆体中NH4+-N含量对NH3的释放有显著影响,通过提高氨氧化速率可以降低NH4+-N含量,进而减少NH3释放. 高温期的温度不适宜硝化细菌生长,所以在整个高温期N2O的产生量较低. 冷却期和腐熟期NH3的挥发量显著减少,该阶段部分NH4+-N的不完全硝化作用或局部厌氧环境中NO2--N和NO3--N的反硝化作用,均会产生以N2O、NO2等氮氧化物主的副产物[35]. N2O的释放量与NOx--N含量相关,Osada等[38]研究发现,当堆体中NOx--N的含量增加时,N2O的产生量也随之增加. NH4+-N还可经同化作用固定为生物有机氮,生物固氮是提高堆肥中氮含量的有效途径. Zhong等[39]研究发现,牛粪堆肥生物固氮主要发生在高温和冷却阶段,在堆肥过程中平均总氮含量增加了1.80%. 胺类、吲哚类和粪臭素类等挥发性含氮有机物主要通过氨基酸脱羧作用、L-色氨酸降解代谢等过程产生[28].
挥发性硫化物(VSCs)本质上是由含硫氨基酸产生的. 蛋白质在蛋白酶、肽酶、裂解酶等酶的作用下水解为氨基酸,进一步地,含硫类氨基酸降解为VSCs[40]. 堆肥中半胱氨酸和蛋氨酸等含硫氨基酸的生物降解可形成H2S和甲硫醇(MT),而H2S和MT的甲基化可分别形成MT和甲硫醚(DMS),其中MT可氧化形成二甲基二硫醚(DMDS)[41]. 此外,DMDS也可以通过DMS的甲基化或MT的脱水作用而形成[42]. DMS和DMDS是不易被氧化的惰性还原性物质,释放量较大,会逐渐累积在周围的空气中[43]. 其中,硫醇类由于具有较高的反应活性,是许多其他VSCs的前体物[44]. H2S是半胱氨酸的主要降解产物. 当堆体中氧化还原电位较低时,硫酸盐在硫酸盐还原菌(SRB)的作用下也会产生H2S[45]. 此外,硫化物甲基化反应产生的MT和DMS经产甲烷菌产甲烷过程中也会生成H2S[46]. 在堆肥过程中,升温期和高温期氧气供应不足,导致有机物降解不完全,是VSCs的主要释放时期. Zhang等[47]研究发现,相比于两个较高的通气量(0.2和0.3 L/min),在低通气量(0.1 L/min)下,VSCs的产生量更加显著. 氧气不足普遍被认为是VSCs产生的主要原因,但有报道称,即使在好氧条件下,VSCs也会大量释放[34].
实际上,堆肥中臭味物质的排放会受到堆肥原料或改良剂等生化成分的影响,不同的生化成分(如淀粉、蛋白质、脂质等)参与不同的微生物代谢途径,形成各种中间产物,从而影响恶臭气体的种类[48]. 研究[49]表明,堆肥过程中以蛋白类物质为原料产生的主要臭味物质为NH3和VSCs,而以纤维类化合物、碳水化合物为原料时则分别为醛类(醛类恶臭气体)、萜烯(萜烯类恶臭气体)和硫化合物,其中产生臭味潜力最高的是蛋白类物质,碳水化合物次之,纤维类化合物最低. 不同的改良剂也会影响微生物的代谢途径,Chen等[50-51]研究发现,微生物会通过控制有机氮的转化来影响NH3的产生,当堆肥原料为蔬菜废弃物时,氨基酸氮和氨基糖氮是NH4+-N的主要来源,而当分别使用麦秸、玉米秸秆作为改良剂时,氨氮、氨基酸氮最容易转化为NH4+-N.
2.2 臭气生成转化的核心微生物
微生物是物质转化的关键驱动力. 细菌由于其代谢的多功能性,相较于真菌及放线菌等在堆肥中更具优势[16]. 微生物分泌出具有特异性功能基因的酶以实现对物质的降解转化,在该过程中温度、pH、含水率等多种因素均会影响微生物的代谢功能和群落变化,进而影响恶臭气体的产生. 因而,了解堆肥中的微生物是减少恶臭气体产生的关键,也是建立有效控制恶臭气体技术的重点.
2.2.1核心微生物
微生物是堆肥化进程的主体. 臭气生成转化所发生的氨化作用、硝化作用、反硝化作用及硫酸盐还原作用分别是由氨化细菌、硝化细菌、反硝化细菌及硫酸盐还原菌驱动完成. 这些微生物是物质代谢的关键微生物,也是恶臭气体生成转化的关键微生物. 不同时期发挥作用的核心微生物有所不同,但在堆肥进程中都是不可或缺的[52].
氨化细菌是氨化作用的重要驱动力,主要存在于堆肥前期,适宜在中性或碱性条件下生长繁殖,是产生NH4+-N的主要菌种. NH4+-N的变化与氨化细菌的数量和活性相关,但NH4+-N的变化滞后于氨化细菌的变化,往往表现为氨化细菌数量的减少,导致NH4+-N含量下降,从而减少NH3的释放量[53]. Li等[54]研究表明,膜覆盖堆肥系统中以假单胞菌属(Pseudomonas)和芽孢杆菌属(Bacillus)为主的氨化细菌的平均相对丰度和共现率均低于非膜覆盖堆肥系统,可有效保留堆体中的氮,减少了58.64%的NH3排放量. Asano等[55]等对牛粪酸化堆肥处理系统中的微生物进行研究,发现Bacillus和乳酸杆菌属(Lactobacillus)是堆体氨化过程的关键菌种,酸化处理条件下NH3排放量低于可检测水平.
硝化细菌多出现在堆肥早期及后期阶段,对温度较为敏感,为嗜温菌,当堆体温度过高时硝化细菌数量有所降低. 这主要是因为,堆肥初期有机质丰富,可为硝化细菌提供良好的生长环境,而进入高温期,耐受高温的硝化细菌得以保留,不耐高温的硝化细菌死亡,到后期温度下降,硝化细菌表现出新的活性[56]. 堆肥体系中研究较多的硝化细菌为氨氧化细菌(AOB)和氨氧化古菌(AOA). 硝化过程中氨氧化是第一个步骤,也是主导NH3和N2O排放的关键阶段,主要由AOB驱动完成. Zhong等[39]对牛粪堆肥中的氮循环进行研究,发现从进入腐熟阶段开始,AOB的数量急剧增加,其中亚硝化单胞菌属(Nitrosomonas)是最丰富的AOB,占比超过氨氧化细菌总数的98%. Meng等[57]研究表明,Nitrosomonas是奶牛粪堆肥中贡献最大的AOB,在堆肥后期最为活跃,是腐熟时期的优势菌属.
反硝化细菌主要存在于堆肥后期,多为厌氧型微生物,在高温、有氧环境中其生长繁殖会受到抑制. 最常见的反硝化细菌大多为变形菌门(Proteobacteria). 在鸡粪、猪粪和牛粪等废弃物堆肥体系中均发现了Proteobacteria是最主要的反硝化微生物,其中在鸡粪堆肥中以Proteobacteria为主的反硝化细菌,占总样本的61.64%~96.68%[58-60]. 在堆肥过程中已经分离出了多种反硝化微生物,如芽孢杆菌属(Bacillus)、假单胞菌属(Pseudomonas)、鞘氨醇单胞菌属(Sphingomonas)及黄杆菌属(Flavobacterium)等,不同时期的反硝化优势菌有所不同. 许本姝等[61]研究发现,反硝化细菌的多样性和丰富度在堆肥中均呈先降低后升高的趋势,升温期、高温期及腐熟期的优势菌属分别为Pseudomonas和陶厄氏菌属(Thauera)、Pseudomonas和无色杆菌属(Achromobacte)及鲍特氏菌属(Bordetella). Zhong等[39]研究表明,牛粪堆肥中高温期嗜盐单胞菌属(Halomonas)是最丰富的反硝化细菌,而腐熟期Labrenziaalba、Pseudomonasflexibili和Steroidobacterdenitrificans是反硝化优势菌.
硫酸盐还原菌(SRB)是一种厌氧菌,对有氧环境极为敏感,常见的主要有脱硫弧菌属(Desulfovibrio)、脱硫杆菌属(Desulfobacter)、脱硫球菌属(Desulfococcus)、脱硫线菌属(Desulfonema)、脱硫肠状菌属(Desulfotomaoculum)以及脱硫单胞菌属(Desulfomomas)等. 甲硫氨酸可被恶臭假单胞菌(Pseudomonasputida)和亚麻短杆菌(Brevibacteriumlinens)等微生物降解转化为MT、DMS、DMDS等VSCs[62]. 硫酸盐还原菌与堆体中H2S的产生密切相关. 例如,在厨余垃圾堆肥中添加园林废物可有效改善堆肥原料的理化特性,降低Desulfotomaculum的相对丰度,从而减少H2S的排放[63];而在沼气残渣膜覆盖堆肥系统中,相较于非膜覆盖堆肥系统,减少了38.13%的H2S释放量,硫酸盐还原优势菌为Desulfovibrio[54]. 此外,局部厌氧条件下产甲烷菌的丰度也与堆肥中H2S的释放有关[64].
2.2.2功能基因
微生物作用下恶臭气体的生成转化由一个或多个功能基因参与完成. 常见的功能基因,如氨化作用中的ureC,编码脲酶,将有机氮矿化为NH4+-N;硝化作用中的AOA-amoA、AOB-amoA和nxrA,分别编码古菌氨单加氧酶、细菌氨单加氧酶和亚硝酸盐氧化还原酶,将NH4+-N氧化为NO2--N、NO3--N;反硝化作用中的napA/narG、nirS/nirK、cnorB/qnorB和nosZ,分别编码硝酸还原酶、亚硝酸盐还原酶、一氧化氮还原酶和一氧化二氮还原酶,促进NO3--N、NO2--N、NO和N2O转化为N2;硫酸盐还原过程中的aprA、dsrA/dsrB,分别编码腺苷5’-磷酸硫酸酐还原酶、异化型亚硫酸盐还原酶,将SO42-、SO32-等还原为H2S等硫化物.
不同形态有机氮的转化需要不同的酶来驱动氨化作用,其中脲酶被广泛应用于农业氨化过程的研究,而堆肥原料中有机肥的前体含有大量尿素,因而脲酶在堆肥过程中对氨化作用尤为重要[65-66].ureC是脲酶的标记基因,可催化尿素氮分解为NH4+-N. Yu等[67]研究了有机-无机共堆肥过程中氮素生物转化相关功能基因的变化,发现ureC的绝对丰度与NH4+-N浓度呈中等程度显著相关.
amoA和nxrA是NH4+-N转化的主要功能基因.amoA通常被认为是NH4+-N氧化为NO2--N的标记基因,能直接影响NH4+-N和NO3--N的含量;nxrA常被用作NO2--N氧化为NO3--N的标记基因.amoA可以对NO3--N产生正效应,且NH4+-N与NO3--N呈负相关,可指示NH4+-N与NO3--N之间发生的转化,amoA是引起NO3--N积累的主要功能基因之一[58,68]. 因此,通过一定的手段来调控amoA和nxrA,促进NH4+-N氧化,可以减少堆体中NH3的释放. Guo等[59]在猪粪堆肥中添加具有硝化作用的巨大芽孢杆菌(Bacillusmegaterium),可以提高高温阶段amoA基因的绝对丰度,增强amoA将NH4+-N氧化为NO2--N的能力,减少NH3的排放. 此外,反硝化过程中NO3--N可作为N2O的反应底物,所以可通过amoA和nxrA实现其对NO2--N和NO3--N的调控,减少NO3--N积累,这对降低堆肥过程中N2O的释放具有积极作用.
narG和napA参与了反硝化的第一步,将NO3--N还原为NO2--N;nirK和nirS参与了反硝化过程的第二步,将NO2--N还原为NO;cnorB和qnorB参与了反硝化过程的第三步,将NO还原为N2O;nosZ则参与了反硝化过程的最后一步,将N2O还原为N2. 其中,nirK、nirS和nosZ常用作反硝化过程的标记基因,Maeda等[69]研究得出,在N2O活跃释放的阶段,检测到许多假单胞杆菌属(Pseudomonas)的nirK和nosZ片段. 降低堆肥中napA/narG、nirS/nirK和cnorB/qnorB的丰度(或削弱其表达),或增加nosZ的丰度(或增强其表达),是减少N2O排放的有效方法. 在堆肥中加入生物炭可降低narG、norB和nirK的绝对丰度,增加nosZ的绝对丰度,从而减少N2O排放[70];而通入交变磁场可削弱narG和nirS的表达(最大削弱率为83.7%和95.5%),并使nosZ的表达增加36.5倍,N2O的释放量减少39.8%[71].
aprA介导SO42-向SO32-转化,在硫酸盐还原过程中起主导作用,但SO42-非常稳定,不易被还原,不能直接作为电子受体,故需要在硫酸腺苷转移酶(Sat)的作用下先生成氧化性较强的腺嘌呤磷酸硫酸盐(APS),再进行下一步还原转化[72].dsrA和dsrB广泛用于研究硫酸盐还原菌(SRB)的群落多样性和数量. 其中,dsrA是SRB的高度保守功能基因,在硫酸盐还原过程中起着关键的酶促作用,存在于所有的SRB中[73].
2.2.3影响核心微生物的微环境因素
温度是堆肥微生物活性的重要影响因素,能够影响微生物的生长速率、酶活性,从而影响微生物的数量和多样性[74]. 微生物的功能会受到温度的影响. 氨化细菌、硝化细菌和反硝化细菌的适宜生长温度多为20~35 ℃[75]. 在40 ℃下的酸性环境中,氨氧化菌和反硝化菌的生长会受到明显的抑制作用[76]. 氨氧化细菌在堆肥的中温期和高温早期变化较大,而氨氧化古菌在高温期晚期、冷却期和腐熟期变化最为明显[77]. 硫酸盐还原菌在堆体温度为35~37 ℃时生长代谢最好,硫酸盐还原为硫化物的速率最大.
含水率对堆肥物料的物理和生物特性具有重要影响,包括营养物质的溶解和传输、微生物的代谢和转移等[78]. 堆肥中最适宜的含水率为50%~70%[79]. 陈雅娟等[80]对鸡粪堆肥过程中硝化细菌的变化进行研究,发现含水率是导致硝化细菌群落结构发生改变的关键因素. Li等[78]研究发现,在含水率为53%的条件下,亚硝酸盐细菌、硝化细菌和固氮细菌表现出较好的内源氮转化和外源固氮作用,堆体中NO3--N和TN的含量较高,在一定程度上减少了含氮类恶臭气体的产生.
微生物的正常生长繁殖需要一定量的碳、氮元素,更需要适宜的C/N. 微生物分解有机物的速率会随C/N的变化而发生变化. C/N在35∶1~25∶1之间有利于微生物的正常生命活动. 在适宜的C/N范围内,适当提高C/N可增加氮转化细菌的相对丰度和促进细胞内的碳代谢,产生更多的ATP和α-酮戊二酸,有利于氨的同化,减少堆肥中含氮类恶臭气体的产生[81-82]. 如在污泥堆肥过程中添加蔗糖(增加碳含量),可同时提高氨同化细菌的数量和相关酶的活性,减少NH3的释放[83].
堆肥中pH为6~9,是影响微生物代谢和繁殖的重要环境因素之一. 堆肥中微生物间的相互作用使得pH处于不断变化的状态. 同时,pH也会对微生物的酶活、酶促反应及代谢产生影响[84]. 硝化细菌适于生长在pH为7.5~8.0的偏碱性环境. 硫酸盐还原菌在pH为 6.5~7.5时可进行良好的生长繁殖,但当堆体pH>8时不利于其生长. 王红等[85]研究发现,当pH<4.5时,氨化细菌的生长活性完全被抑制,当pH>4.5时,氨化细菌的生长活性随时间的延长表现为持续增强,在pH为6.5~8.0时可维持较高的活性,并在pH=7 时其活性达到最高.
通风具有供氧、散热、脱水三大作用,也是影响堆肥微生物的关键因素之一. 当堆体内O2含量为5%~10%时,可使堆肥正常进行[86]. 通过调节通风量,可以调节堆体的反应温度,为微生物的生长提供合适的环境. 胖是等[87]研究发现,通风能显著影响堆肥中的氨化细菌和反硝化菌,并建议在升温期通风量采用0.15 L/min、高温期采用0.05 L/min,可使堆肥产品在达到无害化的基础上减少NH3的排放. 硫酸盐还原菌在溶解氧浓度为4.5 mg/L时能够存活,但在接近饱和溶解氧浓度(9.0 mg/L)时无法存活,故可适量向堆体中通气以抑制其生长繁殖,减少含硫类恶臭气体的释放.
3 生物除臭技术
为了对堆肥产生的恶臭气体进行有效控制,国内外学者研发了多种处理技术,包括物理、化学和生物法等. 相比于物理、化学法除臭,生物法除臭通常具有效率高、无二次污染、操作简便、成本低廉等优点,是去除臭气的有效方法[88]. 但生物法除臭过于依赖于微生物,微生物的活性将直接影响除臭效果,其正常工作的条件是使反应器内的温度、湿度、营养液浓度及pH等参数满足微生物正常生长的需要,在实际应用方面也存在一定的挑战性[89]. 生物法除臭中微生物可利用恶臭组分进行自身代谢并将其转化为无臭组分,如含氮恶臭组分被氧化分解成NH4+、NO2-、NO3-,含硫恶臭组分被氧化分解成S、SO32-、SO42-,是减少恶臭气体最为直接有效的方法[90-91]. 常用的生物法除臭包括原位添加微生物菌剂和异位生物反应器处理(如生物滤池、生物滴滤器和生物洗涤器). 生物除臭技术的优缺点对比如表1[92-94]所示.
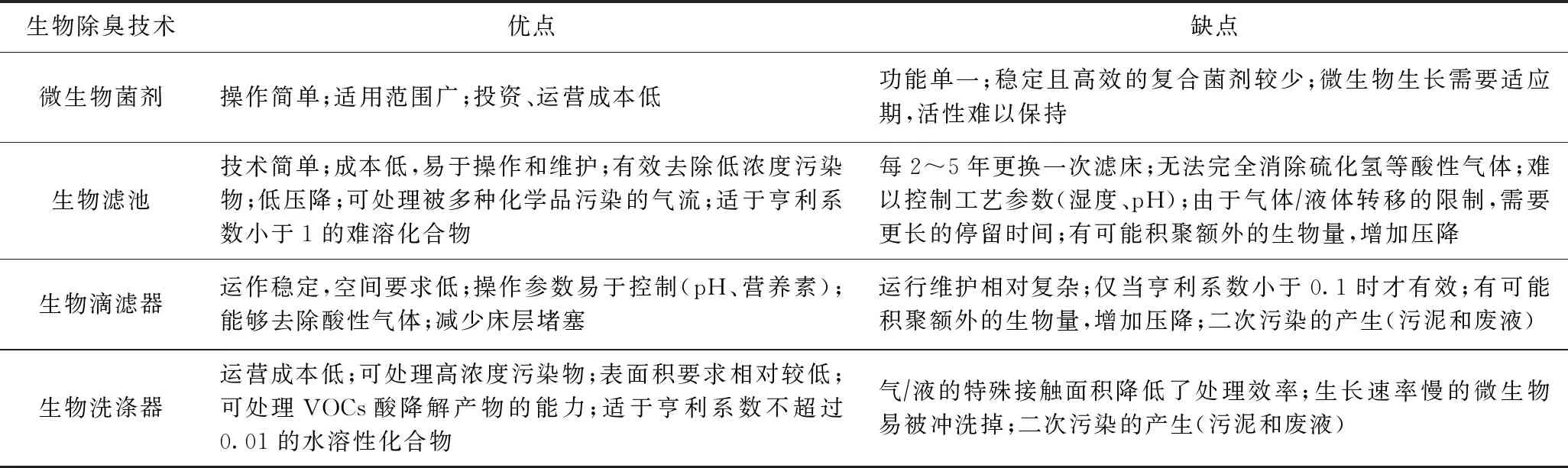
表1 生物除臭技术的优缺点比较[92-94]
3.1 接种外源微生物
在堆肥过程中接种外源微生物,可对臭气起到直接降解作用,在源头上直接减少了臭气的产生,是目前重要的除臭手段. Zhu等[95]研究发现,接种微生物菌剂可以减少NH4+-N产生,降低堆肥过程NH3挥发量. 可见,接种外源微生物同时起到了保氮除臭的作用,是控制含氮类恶臭气体产生的理想方法. 然而,由于臭气组分的复杂性,各组分难以同步达到理想的去除效果,因此有必要研发除臭性能高的复合微生物菌剂. 张生伟等[96]将从堆肥中筛选的数株高效除臭菌和纤维素分解菌群优化组合后制备成复合微生物除臭剂,发现该菌剂在堆肥前20天可使NH3和H2S的去除率分别在70%和60%以上.
3.2 生物反应器
生物滤池可以同时处理多种污染物,特别是对于处理大量低浓度的恶臭气体是一种较为经济的选择. 细菌和真菌是生物滤池中的两个主要微生物菌群,但有报道称,随着细菌种群的增长,也会出现酵母菌、藻类以及高等原生生物(动物、轮虫、线虫等)[97]. 于承泽等[98]研究发现,生物滤池对NH3、H2S、甲硫醇的平均去除率在95%以上,起主要除臭功能的菌属为丝状菌属(Kineothrix)、芽孢杆菌属(Bacillus)、魏斯氏属(Weissella). 生物滴滤器(BTF)是一种处理臭气的有效方法. BTF不仅能减少污染物的排放,而且能够输送营养物质,并且还可通过循环水流控制pH及副产物的积累[99]. BTF中的生物膜上附着有大量细菌和少量真菌,主要发挥作用的为细菌. BTF中最为常见的细菌菌属为假单胞杆菌属(Pseudomonas)、Bacillus、葡萄球菌属(Staphylococcus)、红球菌属(Rhodococcus)等[100]. 虽然BTF中污染物的降解通常是由细菌引起的,但有时真菌也能起到重要作用. 与细菌相比,在低湿度和高酸度的环境中真菌可以更好地生存,因此以真菌为主要降解微生物的BTF具有很大的潜力[101]. 生物洗涤器易于操作,添加的生物填料不易引起堵塞,适用于处理亨利系数不超过0.01的SO2、乙酸、胺、H2S和硫醇类等水溶性气体,也可处理浓度低于 5 000 mg/m2的化合物[102-103]. 而对于处理过程产生的剩余污泥和废液必须进行进一步的后端处理,否则会影响其处理性能.
4 结论与展望
a) 堆肥过程中含氮、硫类恶臭气体种类多、赋存形态与污染途径不一,如何阐明微生物在其中的作用机制仍然是一个复杂的问题. 今后需深入研究堆肥微生物代谢过程中的物质转化途径及相关机理,解析物质全过程转化规律与代谢归趋,并阐明恶臭气体的生物成因与关键节点,构建恶臭气体生成转化的微生物代谢网络,为有机固废的可持续安全利用与恶臭气体的风险控制奠定理论基础.
b) 在现有相关研究中,基于环境因子、微生物及恶臭气体的统计学分析结果表明,堆肥微环境对微生物群落特征影响显著,进而会影响臭气物质的种类、浓度与转化途径. 故可在阐明核心微生物代谢网络的基础上,筛选出影响微生物的微环境主控因子,也将为堆肥臭气过程控制的参数设置提供科学依据.
c) 就末端处理技术而言,生物法处理恶臭气体效益高且环境友好,依然优势显著,应注重与物理、化学方法相结合,侧重制备研发高效除臭复合菌剂、新型生物活性表面剂、高浓度恶臭气体生物反应器等,为分散式有机固废资源化臭气污染控制保驾护航.
猜你喜欢
中国乳业(2022年5期)2022-06-05
中国农业大学学报(2022年3期)2022-05-19
哈尔滨工业大学学报(2022年5期)2022-04-19
建材发展导向(2021年15期)2021-11-05
绿色科技(2021年16期)2021-09-09
食品与发酵工业(2020年21期)2020-11-20
安徽农业科学(2018年30期)2018-11-02
农家顾问(2016年11期)2017-01-06
科学种养(2016年8期)2016-08-11
农村农业农民·B版(2015年9期)2015-10-16
