
重金属胁迫对植物生理生化的影响及其抗性机理研究
2021-12-03 05:39王培军邵海林郭家雁
山西林业科技 2021年3期
葛 坤,王培军,邵海林,郭家雁,杜 宾
(太原学院,山西 太原 030024)
根据公开文献资料显示,我国城市空气存在重金属污染,对环境质量、人身健康、社会发展造成严重威胁。绿化植物因其较广的应用范围,较大的覆盖面积,较长的生命周期,对空气中重金属有吸附、富集、转化、降解作用,可有效提高空气质量[1]。
1 大气中重金属来源
大气中重金属来源一方面为工业排放及化石燃料燃烧,另一方面为土壤中的沙尘。重金属来源的分析方法中,富集因子法(EF)最为常用,不仅可以判断大气中重金属富集的程度,还可以判断其性质[2]。甘小凤等[3]用富集因子法分析得出,西安市PM2.5中Cl,Br,S,Mo,Zn,As,Cd,Pb主要来源于人为活动,而Fe,Cr,Mn,Ti,Ca,K主要来源于岩石风化。杨卫芬等[4]采用富集因子法对南京市雾霾天颗粒物PM2.5中重金属进行分析,得出Cd,Zn,As,Se,Ag,Cu,Pb等来自于人为影响。其中,机动车尾气排放、炼铁、化工厂是主要影响因素。
2 重金属对植物生理生化的影响
2.1 重金属对植物生长发育的影响
Cd,As,Pb等重金属是植物生长的非必需元素,毒性大,可破坏植物生化活性。重金属进入植物体内,会与植物细胞核发生协同作用,改变叶绿体内部结构,减少叶绿素生成,抑制光合作用相关酶的活性,进而抑制植物的光合作用,使植株表现为矮小、叶枯黄[5]。一定浓度的重金属元素,会影响植物对必需元素的吸收并产生拮抗作用,浓度高时可直接导致植物死亡。
2.2 重金属对植物细胞膜透性的影响
植物细胞膜是防止细胞外物质自由进入细胞的天然屏障,同时也为植物生理生化反应提供了稳定的环境。植物细胞膜最重要的特性即选择透过性,有选择地调节、控制细胞内外物质的运输和交换。当植物受到重金属胁迫时,细胞膜的选择性机能受损,透性增加,影响植物叶片细胞质膜的组成。植物失去了保护屏障,离子和有机物渗出细胞膜外,有害物质进入细胞内,从而影响了细胞内酶及代谢反应[6]。通常通过测定植物细胞膜外渗透液电导度和钾离子含量来研究重金属对细胞膜的伤害程度。试验表明,Cd浓度不断增大,玉米幼苗叶片电导度随之增高[7]。水生植物细胞膜外渗透液电导度以及钾离子浓度与 Cd 浓度有显著正相关。在Cd,Pb复合胁迫下,辣椒、小麦等植物细胞膜外渗液电导性与Cd,Pb浓度呈显著正相关,且Cd,Pb存在协同效应[8]。青菜叶片细胞膜透性会随着Cd,Pb,Cr处理浓度增加、处理时间的延长而增大[9]。
2.3 重金属对植物游离脯氨酸的影响
游离脯氨酸是植物体内水溶性最大的氨基酸,其疏水端与蛋白质结合,亲水端与水分子结合。蛋白质利用其较强的水合能力束缚水分子,防止植物体在渗透胁迫条件下脱水变性。当植物细胞含水量很低时,游离脯氨酸能提供足够水分,保证植物细胞正常的生理活动。植物处于逆境胁迫时,游离脯氨酸含量显著增加。因此,游离脯氨酸含量对植物所处环境具有一定指示作用[10]。对不同污染环境下白皮松针叶游离脯氨酸含量的测定发现,城区>郊区>山区,即空气污染严重地区游离脯氨酸含量较高[11]。Cu,Zn,Cd对小麦幼苗叶片游离脯氨酸的产生有诱导作用。重金属胁迫致使植物体水分缺失,通过产生一定量的脯氨酸来调节植物细胞水分平衡,降低植物细胞受损程度[12]。
2.4 重金属对植物保护酶系统的影响
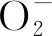
3 植物对重金属的抗性应答
3.1 植物不同部位对重金属的富集
叶片是植物的重要器官之一,可利用气孔及自身分泌的粘液、油脂等吸附重金属,从而起到对大气重金属污染的净化作用。赵曦等[15]通过测定 3 个污染严重地区的荔枝叶片,发现荔枝叶片对Cu,Hg,Cd有很强的富集能力。树皮不仅有减少植物水分蒸发、防止病虫害等功能,还具有富集重金属的功能。El-Hasan T 等[16]发现较光滑树皮而言,粗糙树皮对重金属富集能力更强,老树皮比枝条、树干的富集能力强,主要与其表面较深且不规则的凹槽有关。植物根系从土壤中吸收水分及营养元素的同时吸收多种重金属,Cu,Zn,Mn是植物生长发育所必需的元素,Ni,Pb,Cd,Cr则为非必需元素。因此,Cu,Zn,Mn含量会高于其它重金属元素。总之,植物的叶片、树皮、树干、根系均对重金属具有吸附净化能力。Unterbrunner R 等[17]研究表明,杨树各器官吸附能力为树叶>树皮>树干。车继鲁等[18]测定了香樟各器官对不同浓度重金属的吸附能力,结果表现为为树叶>树干 > 树枝。
3.2 植物细胞对重金属的结合钝化
3.2.1 植物细胞壁对重金属的结合钝化
细胞壁是植物细胞的最外层屏障,是使细胞免受重金属毒害的重要组织,其成分为果胶、半纤维素和纤维素。果胶占植物初生壁的 30%,是细胞壁阳离子结合的主要位点,可通过过氧原子结合金属阳离子[19]。Krzeslowska 等研究表明,在重金属Pb,Cd,Cu,Zn胁迫下,果胶甲酯酶活性增强,使更多低钾酯化果胶生成并重新排布,从而增加了细胞壁对重金属的吸收和积累。纤维素是细胞壁的重要组成成分,其中的葡萄糖单体形成高度对称的双螺旋结构,通过氧原子共用电子对吸附重金属离子。细胞壁中磷酸根与细胞壁蛋白质可将重金属离子钝化,形成重金属-磷酸盐、重金属-碳酸盐结合态,铅离子在细胞壁上主要以此形式存在[20]。
3.2.2 植物细胞膜对重金属的结合钝化
植物通过细胞膜控制离子进入细胞,一方面细胞膜对重金属离子有结合钝化作用,主要表现为重金属与细胞膜上的氨基酸、蛋白质、糖类、脂类等官能团结合形成鳌合物;另一方面通过转运蛋白将重金属离子转运流出[21]。细胞膜表面重金属转运蛋白有9种(表1),分为吸收蛋白和排出蛋白。吸收蛋白存在于细胞质膜中,主要功能是将重金属转运至细胞质。排出蛋白属于解毒蛋白,可对重金属胁迫起到重要防御作用,主要功能是将重金属从细胞质排出[22]。

表1 重金属转运蛋白的细胞定位及作用方式
3.3 植物代谢对重金属的解毒
植物代谢对重金属的解毒,主要是利用金属硫蛋白(MT)、植物络合素(PC)、热激蛋白(HSP)、有机酸、氨基酸、多胺(Pas)、谷胱甘肽(GSH)等物质将重金属络合,或形成复合物,从而降低重金属对植物的危害[23]。植物体中含有约50种金属硫蛋白(MT),也称金属硫组氨酸三甲基内盐,其分子量很低,富含易与重金属离子结合的疏基。在重金属胁迫环境中,MT可消除OH,O2-等自由基,起到保护植物细胞免受离子辐射和抗氧化物伤害的作用[24]。植物络合素(PC)由不同长度的肽链构成,肽链越长,与重金属结合能力越强。在重金属胁迫环境中,PC合成酶被金属离子激活,以谷胱甘肽(GSH)为底物催化合成PC.PC在植物中广泛存在,其类型与植物和重金属种类有关[25]。有研究表明,PC可与植物细胞吸收的Cd复合形成分子质量不同的复合物,进而起到对Cd的解毒作用。热激蛋白(HSP)可参与细胞增殖、调控免疫系统等生理活动,协助蛋白质折叠、装配和转运。在重金属胁迫下,起到对蛋白质的保护和修复作用,将重金属诱导变性的蛋白质重新折叠,维持蛋白质的稳定性[26]。有机酸、氨基酸(脯氨酸等)是植物重金属络合的主要配位体,含有羟基、羧基、氢基等官能团,可与重金属结合形成稳定化合物,并参与重金属离子的吸收、运输、积累过程,起到对植物重金属钝化解毒的作用[27]。多胺(Pas)可为植物提供能量,抑制还原型辅酶2(NADPH)氧化,消灭活性氧,维持线粒体氧化磷酸化和NADPH稳定,减轻重金属对植物细胞膜和蛋白质的伤害。谷胱甘肽(GSH)、抗坏血酸等非酶类抗氧化剂,对植物重金属有解毒作用。谷胱甘肽(GSH)是含有-SH信号的物质,普遍存在于植物体内。GSH与重金属离子发生螯合反应,从而防止自由基和脂类过氧化物对细胞膜的伤害。为防止GSH上的疏基被氧化,形成GSSG,GSH在转移酶的作用下进行GSH-二硫化物交换反应,以保证足够的GSH与重金属等有害物质结合[28]。通常情况,GSH作为PC与重金属离子螯合反应的供体,重金属离子在与PC螯合前先与GSH螯合,从而达到对重金属的解毒作用[29]。
4 展望
重金属对植物的危害及植物对重金属的解毒机制是当前研究的热点。重金属通过干扰细胞正常代谢而影响植物生长发育,甚至造成植物体损伤。因植物自身代谢及重金属元素毒性的差异,植物对重金属的吸附、适应、解毒机制极其复杂,仍需进一步研究。
目前,国家对环境保护极为关注和重视,从源头上对重金属污染排放较为严重的化工厂、冶金矿山等进行了严格治理,加大对繁殖速率快、地上部分密集的重金属超富集植物的驯化和推广。同时引入转基因技术,有针对性地将某重金属元素的富集基因转移到植物体内,达到对多种重金属元素的超富集是今后研究的重要方向之一。
猜你喜欢
中国真菌学杂志(2022年1期)2023-01-06
中老年保健(2022年3期)2022-11-21
安徽化工(2021年3期)2021-05-29
心肺血管病杂志(2020年5期)2021-01-14
三农资讯半月报(2020年18期)2020-10-14
棉花学报(2020年3期)2020-08-08
读与写(2019年31期)2019-11-26
科学与财富(2019年15期)2019-10-21
数码世界(2018年1期)2018-12-23
饮食科学(2016年9期)2016-11-18
