
miR-34a 对人牙周膜干细胞成骨分化的促进作用及其机制
2021-12-07 09:00王训霞
吉林大学学报(医学版) 2021年6期
董 霞, 王训霞, 杨 芳
(1.山东省青岛市海慈医疗集团口腔科,山东 青岛 266000;2.山东省青岛市立医院口腔医学中心,山东 青岛 266000)
牙周膜干细胞(periodontal ligament stem cells,PDLSCs)是一种具有与其他干细胞相似的具有自我更新和克隆形成特性的成体干细胞,在一定条件下具有成骨、神经和脂肪等多方向分化的能力[1]。研究[2]显示: PDLSCs不仅具有再生和修复牙槽骨组织的能力,而且还具有调节牙周组织分化和免疫调节的能力,是牙周组织再生和骨质缺损修复的重要种子细胞。然而PDLSCs 的成骨分化不仅受多种炎性因子的影响,还受内部错综复杂信号通路的调控。微小RNA-34a(microRNA-34a,miR-34a)作为一种小分子RNA,参与转录后mRNA水平表达调控。研究[3-4]显示: 抑制miR-34a 可增强人类脂肪来源的间充质干细胞增殖,促进成脂和成骨分化潜能。miR-34a 可能通过MAPK 途径调节PDLSCs 的成骨分化过程。上述研究提示:miR-34a 参与调控PDLSCs 的成骨分化,但其具体调控机制尚未证实。解整合素金属蛋白酶10(a disintegrin and metalloprotease 10,ADAM10)是一种Notch 信号通路最关键的限速酶,能够介导膜外段Notch 水解促进Notch 信号通路的活化,进而调控干细胞的分化潜能[5-6]。研究[7]显示:Notch信号通路在PDLSCs 的成骨过程中起关键作用,但miR-34a 是否通过Notch 信号通路调控PDLSCs 成骨分化尚不清楚。本研究探讨miR-34a 是否通过靶向调控ADAM10 表达进而介导Notch 信号通路影响PDLSCs 成骨分化。
1 材料与方法
1.1 质粒、主要试剂和仪器miR-34a 过表达质粒(pMSCV-miR-34a)及其阴性对照空载质粒(pMSCV-vector)的构建及其慢病毒的包装、浓缩、纯化以及慢病毒滴度(2×108TU·mL-1)测定均由吉满生物科技(上海)有限公司完成;miRNA-34a 模拟物(miR-34a mimics)和阴性对照miRNA(miRNA NC)由上海吉玛公司合成。α -MEM 培养基和胎牛血清(fetal bovine serum,FBS)购自美国Gibco 公 司,荧光抗体FITCCD90、APC-CD105、APC-CD34 和PE-CD45 购自美国BD Biosciences 公司,逆转录试剂盒和绿色荧光定量试剂盒均购自日本TaKaRa 公司,碱性磷酸酶(alkaline phosphatase,ALP)检测试剂盒(PNP 比色法)购自北京雷根生物技术有限公司,Notch1 抗体、Notch 胞内结构域(Notch intracellular domain,NICD)、Hes 家族bHLH 转录因子1(Hes family bHLH transcription factor 1,Hes1)抗体、ADAM10 抗体和GAPDH 抗体购自英国Abcam 公司,辣根过氧化物酶标记的山羊抗兔或抗小鼠IgG二抗购自武汉博士德生物工程有限公司,茜素红染液购自北京索莱宝科技有限公司。Dual-Luciferase®Reporter System购自美国Promega 公司,流式细胞仪购自美国BD 公司,荧光定量PCR(Real-time fluorescence quantitative PCR,RT-qPCR)仪购自美国ABI 公司,全自动凝胶成像分析仪购自上海天能公司,酶标仪购自美国BioTek 公司。
1.2 PDLSCs 的分离和鉴定收集山东省青岛市海慈医疗集团口腔科18~24岁患者因正畸而拔除且健康完整的前磨牙或第三磨牙,无菌条件下分离牙周膜组织并采用含2% 双抗(链霉素/青霉素)的PBS缓冲液清洗干净,剪碎,加入0.5% Ⅰ型胶原酶消化45 min,然后1 500 r·min-1离心5 min,弃上清,加入含10% FBS 的α-MEM 培养液重悬,接种于6 孔细胞培养板并置于37℃、5% CO2培养箱中培养,每3d换液1次,待细胞从组织块周围爬出并长至80% 融合率时,采用有限稀释法挑选单克隆并扩大培养。取第3代分离得到的细胞,采用流式细胞术检测间充质干细胞表面分子标志物CD90、CD105 和造血干细胞表面标志物CD34 及CD45 的表达来鉴定PDLSCs。本研究经山东省青岛市海慈医疗集团伦理委员会批准,所有患者均签署知情同意书。
1.3 细胞感染取第3代PDLSCs,以每孔1×105个细胞的密度接种于6 孔细胞培养板中,待细胞生长至铺板量达到约70% 时,将细胞分为空白对照组(不经任何处理)、空载组(感染空载慢病毒)和miR-34a 过表达组(感染miR-34a 过表达慢病毒)。按照病毒感染复数(multiplicity of infection,MOI)为50∶1分别加入miR-34a 过表达慢病毒及其阴性对照空载慢病毒,在37℃、5% CO2培养箱中培养12h后,更换新鲜培养液进行培养。感染48h后采用嘌呤霉素筛选稳定表达株,并采用RT-qPCR法检测各组PDLSCs中miR-34a表达水平。
1.4 成骨诱导分化取第3代PDLSCs 或经慢病毒感染的PDLSCs,以5×104mL-1的密度接种到6 孔细胞培养板,采用α-MEM 培养液将细胞常规培养至70% 融合率时,弃去原培养基,加入成骨诱导培养基(含10%FBS的α-MEM培养液、10 mmol·L-1β-甘油磷酸钠、50 mg·L-1维生素C和10 nmol·L-1地塞米松)进行培养,每3d换液1次,按实验需要进行后续实验。
1.5 RT-qPCR 法检测各组细胞中miR-34a、ALP、Runx2 和OCN mRNA 表达水平取成骨诱导分化的或经慢病毒感染后的PDLSCs,根据RNA 提取试剂盒说明书,提取总RNA,将RNA 逆转录合成cDNA。以cDNA 为模板链,采用RT-qPCR 试剂盒进行扩增。miR-34a 引物序列:Forward 5′-AGGGTGGCAGTGTCTTAGC-3′,Reverse 5′-GAGCAGGGTCCGAGGT-3′;U6 引物序列: Forward 5′-CTCGCTTCGGCAGCACA-3′,Reverse 5′-AACGCTTCACGAATTTGCGT-3′;ALP 引 物 序列:Forward 5′-ACTGGTACTCAGACAACGAGAT-3′,Reverse 5′-ACGTCAATGTCCCTGATGTTATG-3′;Runx2 引物序列: Forward 5′-TGGTTACTGTCATGGCGGGTA-3′,Reverse 5′-TCTCAGATCGTTGAACCTTGCTA-3′;OCN 引 物序 列: Forward 5′-AGCCCATTAGTGCTTGTAAAGG-3′,Reverse 5′-CCCTCCTGCTTGGACACAAAG-3′;GAPDH 引物序列: Forward 5′-CAGGGCTGCTTTTAACTCTGG-3′,Reverse 5′-TGGGTGGAATCATATTGGAACA-3′。反 应程序:95℃预变性10 min,95℃变性15 s,60℃退火30 s,72℃延伸30 s,共40个循环。采用2-ΔΔCt法计算各组PDLSCs 中miR-34a、ALP、Runx2 和OCN mRNA表达水平。
1.6 Western blotting法检测各组PDLSCs 中Notch1、ADAM10、Hes1 和NICD蛋白表达水平取成骨诱导分化的或慢病毒感染后的PDLSCs,加入含有蛋白酶抑制剂的裂解缓冲液进行裂解30 min,12 000 r·min-1离心25 min,取上清液,BCA 法测定蛋白浓度。取20 μg 总蛋白煮沸变性后,采用10% SDS-聚丙烯酰胺凝胶电泳分离蛋白并将蛋白转移到聚偏氟乙烯(polyvinylidene fluoride,PVDF)膜上,5%脱脂牛奶室温封闭45 min 后,TBST 洗涤3次,分别加 入 一 抗Notch1( 1 ∶1 000 )、ADAM10( 1 ∶500)、Hes1(1∶500)、NICD(1∶2 000)和GAPDH(1∶1 000),置于4℃中孵育过夜。TBST 洗涤3次,加入辣根过氧化物酶标记的山羊抗兔或抗小鼠IgG 二抗体,室温孵育1 h,采用超敏ECL 化学发光试剂盒进行显影曝光,采用Image J 软件检测各蛋白条带的灰度值,以目的蛋白条带灰度值与GAPDH蛋白条带灰度值比值表示目的蛋白表达水平。
1.7 茜素红染色检测各组PDLSCs 矿化结节情况各组慢病毒感染的PDLSCs 进行成骨诱导21 d后,采用PBS 缓冲液洗涤3次,再采用4% 多聚甲醛将细胞固定30 min,PBS 缓冲液洗涤3次,滴加适量茜素红染液对细胞进行染色5 min,PBS 缓冲液清洗掉染液后,将细胞置于倒置显微镜上观察矿化结节情况,采用Image-Pro Plus 6.0 软件进行矿化结节定量分析。矿化结节形成量=矿化结节形成面积/细胞面积×100%。
1.8 各组PDLSCs 中ALP 活性检测各组慢病毒感染的PDLSCs 进行成骨诱导14 d 后,弃去培养基,收集细胞 沉 淀并裂解细胞,1 500 r·min-1离心15 min,取上清。根据ALP 检测试剂盒说明书操作,加入显色底物并充分混匀,37℃孵育10 min,终止反应,采用酶标仪在405 nm 波长处读取吸光度(A)值,以A 值表示各组PDLSCs 中ALP 活性。
1.9 双荧光素酶报告系统检测miR-34a 与ADAM10 基因的靶向关系采用靶基因预测软件TargetScan 7.1 对miR-34a 与ADAM10 基因结合位点进行预测,构建野生型(WT)和突变型(Mut)ADAM10 3′-UTR荧光素酶报告质粒,并与miR-34a mimics 和miR-34a NC 共转染至293T 细 胞中。共培养48h后,采用双荧光素酶报告系统试剂盒分别检测萤火虫荧光素酶活性和海肾荧光素酶活性,以萤火虫荧光素酶活性/海肾荧光素酶活性表示相对荧光素酶活性(R/F)。
1.10 统计学分析采用SPSS 21.0 统计软件对数据进行统计学分析。细胞表面标志物CD90、CD105、CD34 和CD45 阳性表达率,各组PDLSCs中miR-34a、ALP、Runx2 和OCN mRNA 表达水平,各组PDLSCs 中Notch1、ADAM10、Hes1 和NICD 蛋白表达水平,相对荧光素酶活性,细胞矿化结节形成量以及细胞中ALP 活性等均呈正态分布,以±s 表示,多组间样本均数比较采用单因素方差分析,组间两两比较采用LSD-t检验。以P<0.05 为差异有统计学意义。
2 结 果
2.1 PDLSCs 的鉴定流式细胞术检测结果显示:分离所得细胞间充质干细胞表面标志物CD90 和CD105 呈阳性表达,其阳性表达率分别为(94.24±1.11)%和(92.09±2.17)%,而造血干细胞表面标志物CD34 和CD45 呈阴性表达,其阳性表达率分别为(0.51±0.07)% 和(1.22±0.19)%。见图1。
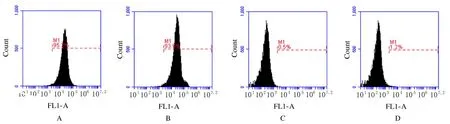
图1 PDLSCs 的鉴定Fig.1 Identification of PDLSCs
2.2 各组PDLSCs 中miR-34a、ALP、Runx2 和OCN mRNA 及ADAM10 蛋白表达水平成骨诱导分化第0、7 和14天检测结果显示:与第0天比较,PDLSCs 中miR-34a 以及成骨标志物ALP、Runx2和OCN mRNA 表达水平逐渐升高(P<0.05)。见图2。Western blotting 法检测结果显示:与成骨诱导第0天比较,成骨诱导分化第7 和14天后,PDLSCs 中ADAM10 蛋白表达水平降低(P<0.05)。与成骨诱导第7天比较,成骨诱导第14天PDLSCs 中ADAM10 蛋白表达水平降低(P<0.05)。见图3。

图2 各组PDLSCs 中miR-34a、ALP、Runx2 和OCN mRNA 表达水平Fig.2 Expression levels of miR-34a, ALP, Runx2,and OCN mRNA in PDLSCs in various groups

图3 各组PDLSCs 中ADAM10 蛋白表达电泳图(A)和直条图(B)Fig.3 Electrophoregram(A) and histogram(B) of expression levels of ADAM10 protein in PDLSCs in various groups
2.3 miR-34a 与ADAM10 基因的靶向调控关系
RT-qPCR 法检测结果显示:与空白对照组和空载组比较,miR-34a过表达组PDLSCs中miR-34a表达水平明显升高(P<0.05)。见图4。靶基因预测软件TargetScan分析结果显示: miR-34a 与ADAM10 基因存在靶向互补序列。见图5A。双荧光素酶报告系统检测结果显示:与miR-34 NC组比较,miR-34a mimics组野生型(ADAM10-Wt)ADAM10 3′-UTR 报告基因的相对荧光素酶活性明显降低(P<0.05),但miR-34a mimics组突变型(ADAM10-Mut)ADAM10 3′-UTR 报告基因的相对荧光素酶活性无明显变化(P>0.05),见图5B。Western blotting 法检测结果显示:与空白对照组和空载组比较,miR-34a 过表达组PDLSCs 中ADAM10 蛋白表达水平明显降低(P<0.05)。见 图5C 和5D。

图4 各组PDLSCs 中miR-34a mRNA 表达水平Fig.4 Expression level of miR-34a in PDLSCs in vari⁃ous groups

图5 miR-34a 与ADAM10 基因表达的关系Fig.5 Relationship between miR-34a and ADAM10 gene expression
2.4 各组PDLSCs中ALP活性和ALP、Runx2 及OCN mRNA 表达水平茜素红染色结果显示:与空白对照组和空载组比较,miR-34a 过表达组细胞中矿化结节形成量明显增加(P<0.05)。见图6 和7。与空白对照组和空载组比较,miR-34a 过表达组细胞中ALP 活性和ALP、Runx2 及OCN mRNA表达水平明显升高(P<0.05)。见图8 和9。

图6 各组PDLSCs 中矿化结节形成情况(茜素红,×100)Fig.6 Formation of mineralized nodules in PDLSCs in various groups(Alizarin red, ×100)

图8 各组PDLSCs 中ALP 活性Fig.8 ALP activities in PDLSCs in various groups
2.5 各组PDLSCs 中Notch1、NICD 和Hes1蛋白表达水平Western blotting 法检测结果显示:成骨诱导14 d 后,与空白对照组和空载组比较,miR-34a过表达组PDLSCs 中Notch1、NICD 和Hes1 蛋白表达水平明显降低(P<0.05)。见图10。

图10 各组PDLSCs 中Notch1、NICD 和Hes1 蛋白表达电泳图(A)和直条图(B)Fig.10 Electrophoregram(A) and histogram(B) of expressions of Notch1, NICD, and Hes1 proteins in PDLSCs in various groups
3 讨 论
牙周膜是一种薄的非矿化结缔组织,具有成骨细胞样特征。PDLSCs 存在于牙周韧带中,参与牙周组织的再生[8]。自从2004年首次报道PDLSCs以来,近年来学者对其进行了广泛的研究[9]。到目前为止,PDLSCs 的性质、微环境对PDLSCs 的影响以及调节其功能的机制尚不清楚。干细胞在再生治疗中的应用迅速发展,促进了对成体干细胞的研究。由于PDLSCs 具有先天性的成骨分化能力,因此研究PDLSCs 成骨分化过程中的特殊调控因素,将有助于PDLSCs 在临床治疗和生物学研究中的应用。

图7 各组PDLSCs 中矿化结节形成量Fig.7 Formation amounts of calcified nodules in PDLSCs in various groups

图9 各组PDLSCs 中ALP、Runx2 和OCN mRNA 表达水平Fig.9 Expression levels of ALP, Runx2, and OCN mRNA in PDLSCs in various groups
miRNAs 是调节转录后基因表达的小非编码RNA。特异性miRNAs 在发育过程中影响细胞周期、细胞分化和细胞凋亡等[10]。研究[11-12]显示:miR-34a 的缺失扩大了小鼠多功能干细胞的发育潜能,说明miR-34a 参与调控小鼠胚胎干细胞和诱导多功能干细胞的分化能力。青蒿琥酯诱导骨髓间充质干细胞中miR-34a 高表达,通过miR-34a/DKK1/Wnt 途径促进成骨细胞分化。另外,miR-34a 还与lncRNA 形成竞争性内源性关系介导肿瘤相关间充质干细胞的上皮间质转化[13],说明miR-34a 通过不同的形式影响各类干细胞的分化和细胞行为。本研究结果显示:在PDLSCs 成骨诱导过程中,随着诱导时间的延长,细胞中miR-34a 表达水平逐渐上调,且呈时间-剂量趋势,说明miR-34a 参与了PDLSCs的成骨分化。本研究结果显示:随着成骨诱导时间的增加,PDLSCs 中ADAM10 的表达水平逐渐下调,这与研究[14]报道的ADAM10 参与肌肉干细胞分化的结论一致。双荧光素酶报告系统结果显示:miR-34a 能靶向调控ADAM10 的表达,并且miR-34a 过表达可降低PDLSCs 中ADAM10 蛋白表达水平。同时也有研究[15]显示:ADAM10 是miR-34a的靶基因,能被miR-34a靶向负调控,且ADAM10 过表达能逆转miR-34a 对结直肠癌细胞增殖、迁移和侵袭能力的抑制作用。虽然miR-34a和ADAM10 均参与PDLSCs 的成骨分化,且miR-34a 能靶向抑制ADAM10 表达,但其具体调控通路尚不清楚。
本研究结果显示: ALP 活性和茜素红染色以及成骨标志物的表达水平一致,表明miR-34a 过表达能明显促进PDLSCs 成骨分化程度。结合miR-34a 与ADAM10 的靶向关系,可知miR-34a 通过靶向下调ADAM10 促进PDLSCs 成骨分化。这与miR-34a 过表达可抑制牙周膜细胞(periodontal ligament cell,PDLCs)体外成骨分化能力[16]不一致。可能原因是PDLSCs 与PDLCs 在生物学特性和功能方面存在较大差异[17],且作为牙周组织再生的种子细胞,PDLSCs 成骨能力远强于PDLCs。因此miR-34a 对两者的成骨分化作用不同。Notch 信号通路能调节多种细胞的分化和发育,由Notch 受体、配体、DNA 结合转录因子CSL 以及其他效应物组成。Notch1 是哺乳动物中表达的4种Notch 受体之一,在Notch 信号的激活过程中,Notch1 受体与配体相互作用后,NICD 与DNA 结合转录因子CSL 异源二聚体形成核转录复合物,并诱导下游靶基因如HES 和HEY 等的转录表达。研究[18]显示: 抑制Notch信号通路可促进PDLSCs体外成骨分化。但也有研究[19-20]显示:激活Notch 信号通路能促进干细胞的成骨分化。两者存在差异的原因可能是由于在不同个体或同一个体不同细胞中,Notch 受体表达存在差异,其表达也会因个体和种属差异而有所不同。ADAM10 是Notch 信号通路的关键限速酶,在多种细胞Notch信号通路可以影响细胞行为。因此,本文作者推测ADAM10 极有可能通过影响Notch 信号通路来干预PDLSCs 的成骨分化。因此本研究对Notch 信号通路相关蛋白Notch1、NICD 和Hes1 等蛋白表达水平进行了检测,结果显示miR-34a 过表达可明显抑制Notch1、NICD 和Hes1 等蛋白表达,初步说明miR-34a 可能通过靶向下调ADAM10 抑制Notch 信号通路来促进PDLSCs 的成骨分化能力。
综上所述,miR-34a 过表达靶向下调ADAM10,通过抑制Notch 信号通路促进PDLSCs成骨分化,本研究为PDLSCs 在临床上的骨再生治疗提供了依据。
猜你喜欢
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
口腔医学(2021年10期)2021-12-02
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
肝博士(2020年5期)2021-01-18
山东医药(2020年9期)2020-05-20
中华老年口腔医学杂志(2016年2期)2017-01-15
中国病理生理杂志(2015年8期)2015-12-21
天津护理(2015年4期)2015-11-10
