
伯氏疟原虫红细胞膜相关蛋白1 缺失突变体的构建和鉴定
2022-03-24 08:21魏超史小雨王倩
天津医科大学学报 2022年2期
魏超,史小雨,王倩
(天津医科大学基础医学院免疫学系,天津 300070)
疟疾是一种由疟原虫(Plasmodium)感染引起的以按蚊为媒介传播的感染性疾病,主要流行于热带和亚热带地区[1]。虽然我国在2019 年获得消除疟疾认证,但根据世界卫生组织不完全统计,2019 年全球仍有大约2.29 亿人感染疟疾,其中40.9 万人死于该疾病[2]。鉴于目前仍无有效的疟疾疫苗,因此对疟原虫输出到宿主细胞表面蛋白的研究就显得尤为重要。
疟原虫入侵的成熟红细胞是终末分化细胞,缺乏细胞器和主要的生物合成过程。当疟原虫侵染红细胞后,会分泌输出大量蛋白质进入宿主红细胞胞浆和胞膜,导致红细胞重塑,变形能力降低,促进感染红细胞与血管内皮细胞的黏附,从而使疟原虫感染红细胞逃避脾脏的免疫清除[3-5]。因此,研究疟原虫输出蛋白可以更好地探究宿主红细胞的免疫原性及免疫逃逸机制,为研制疫苗提供潜在的抗原靶点。红细胞膜相关蛋白1(erythrocyte membrane associated protein -1,EMAP1)蛋白是目前在伯氏疟原虫(Plasmodium berghei,Pb)中发现的被运输到红细胞膜的蛋白质[6]。本课题组前期研究发现,敲除内质网塑形蛋白PbYop1(P.bergheiYop1),红内期伯氏疟原虫生长缓慢,且显著抑制脑型疟疾的发生[7]。对感染红细胞膜表面蛋白进行质谱分析发现,PbYop1 缺失疟原虫较野生型疟原虫感染红细胞膜表面的PbEMAP1 表达量降低,推测PbEMAP1 可能与PbYop1 缺失所致的疟原虫致病力减低相关。为研究PbEMAP1对疟原虫致病性的影响及其潜在的分子机制,本研究利用疟原虫基因敲除常用载体pL0035,构建pL0035-ΔPbEMAP1 重组质粒,基于双臂同源重组的原理,在伯氏疟原虫中敲除EMAP1,获得PbEMAP1 缺失的单克隆虫株,并获得了特异性识别PbEMAP1的抗体,为后续研究疟原虫输出蛋白提供必要的分子工具。
1 材料和方法
1.1 材料 伯氏疟原虫ANKA 株(clone 15Cy1)由罗格斯大学新泽西医学院Purnima Bhanot 副教授提供;大肠杆菌质粒载体pL0035 于本实验室保存(见图1);限制性内切酶SacⅡ、KpnⅠ、EcoRⅠ、Apa1 和NEBuilder®RHiFi DNA Assembly 试剂盒购于NEB 公司;细胞核转染试剂盒购于Lonza 公司;感受态细胞Mach1-T1 购自北京博迈德基因技术有限公司;DIG High Prime DNA 标记及杂交检测试剂盒Ⅱ购于Roche 公司;质粒小量抽提试剂盒、PCR 产物纯化试剂盒和DNA 凝胶回收试剂盒购于全式金生物有限公司,基因组DNA 提取试剂盒购于QIAGEN 公司。

图1 pL0035 质粒图谱Fig 1 Plasmid map of pL0035
1.2 方法
1.2.1 重组质粒pL0035-ΔPbEMAP1 的构建 以野生型PbANKA 基因组DNA 为模板,PCR 合成EMAP1基因编码区上、下游同源臂Up HR arm(719 bp)和Down HR arm(763 bp)并引入相应酶切位点(表1)。PCR 扩增条件为:98℃预变性5 min,以98℃30 s,55℃20 s,68℃23 s 为一个循环,循坏35 次,最后68℃延伸5 min,4℃终止反应。PCR 产物经琼脂糖凝胶电泳鉴定后纯化。将经KpnⅠ和EcoRⅠ酶切的pL0035 质粒以及Down HR arm 在T4 连接酶的催化下进行连接,得到质粒pL0035-Down HR arm。将连接产物转入Mach1-T1 感受态细胞,涂布到含氨苄的固体培基37℃培养16 h。次日挑取单个阳性菌落接种到含氨苄的液体培养基,37℃,220 r/min,12 h。从菌液中提取质粒DNA,酶切鉴定后测序。测序正确的质粒Sac2Ⅱ酶切,将Up HR arm 和pL0035-Down HR arm 通过DNA Assembly 方式连接,50℃,30 min。连接产物转入感受态细胞后进行氨苄筛选培养,挑取阳性单个菌落扩增并提取DNA 酶切和测序鉴定。鉴定正确的重组质粒pL0035-ΔPbEMAP1 大量制备后经ApaⅠ和EcoRⅠ酶切线性化备用。
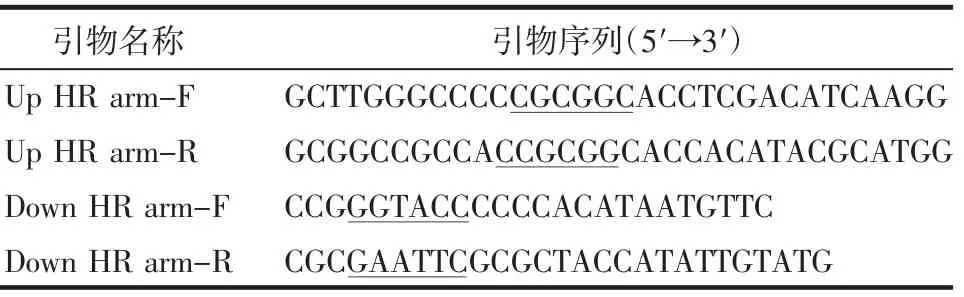
表1 PCR 扩增目的片段引物序列Tab 1 Primer sequences of PCR amplification target fragments
1.2.2 疟原虫转染 经尾静脉注射冻存的野生型Pb ANKA 株疟原虫感染Wistar 大鼠。每日取尾尖血制备薄血膜涂片,吉姆萨染色后油镜下观察并计数虫血率。当虫血率为3%~5%时,心脏取血进行体外同步化培养16~23 h。次日,制备薄血膜涂片观察其同步化情况,当80%以上的疟原虫发育为成熟裂殖体时,用碘海醇密度梯度分离液分离裂殖体感染红细胞备用。取30 μL 裂殖体感染红细胞、10 μg 线性化的质粒和100 μL 的电转液混合后电击,之后迅速加入100 μL 完全培养基,轻柔混匀后经尾静脉注射感染BALB/c 小鼠。
1.2.3 重组疟原虫筛选及单克隆筛选 感染小鼠次日尾尖取血涂片镜检,当观察到有疟原虫时,对小鼠给予乙胺嘧啶饮水,进行药物筛选,发生基因重组的疟原虫获得耐药性,可在乙胺嘧啶作用下继续生长,从而获得重组疟原虫。当虫血率增至5%左右时,取血冻存并利用PCR 对疟原虫基因型进行初步鉴定。利用有限稀释法,对鉴定为杂合的疟原虫进行单克隆筛选。
1.2.4 Southern 印迹鉴定重组疟原虫 PCR 扩增PbEMAP1 Down HR arm(757 bp)DNA 片段作为检测探针的模板(PCR 扩增探针引物:F:CCCCACATAATGTTCTTTATCAC,R:GCGCTACCATATTGTATGT-CATC),按照Roche DIG High Prime DNA 标记及杂交检测试剂盒Ⅱ说明书的方法制备地高辛标记的探针;WT 和ΔPbEMAP1 疟原虫基因组DNA 经限制性内切酶EcoRⅤ和HindⅢ消化过夜后,在0.8%的琼脂糖凝胶中电泳分离;通过毛细管虹吸印迹法将凝胶中的DNA 片段转移至尼龙膜上;转膜完成后将尼龙膜上的DNA 变性、中和;通过紫外交联法将DNA 固定于膜上,预杂交4~6 h;将标记好的探针经高温变性为单链后加入杂交瓶中杂交过夜;次日,洗膜后于室温封闭1 h;加入地高辛抗体(1:10 000)室温孵育45 min,洗膜后加检测液孵育5min,均匀滴加CSPD 显色液后曝光。
1.2.5 多克隆抗体制备 对PbEMAP1 蛋白的序列和结构进行预测分析[8],避开有二级结构的区域,挑选3 个肽段(图2),混合后免疫家兔;然后分别用3个肽段对免疫血清进行亲和纯化,获得抗PbEMAP1 蛋白的多克隆抗体。

图2 预测的PbEMAP1 蛋白三维结构Fig 2 The predicted three-dimensional structure of the PbEMAP1 protein
1.2.6 蛋白提取和免疫印迹实验 取20 μL 感染红细胞,加入200 μL 蛋白裂解液,充分裂解后离心,4℃,10 000 r/min,10 min。取上清至新的EP 管中即疟原虫感染红细胞蛋白样品。用BCA 法对蛋白样品进行定量。调节蛋白上样量后与上样缓冲液混合,65℃,15 min;进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳,通过湿转的方法将蛋白转印至PVDF 膜,5%脱脂牛奶室温封闭1 h;孵育抗PbEMAP1 抗体(1:500 稀释),4℃过夜;次日PBST 洗膜3 次,10 min/次;室温孵第二抗体(1:3 000),1 h;PBST 洗膜3 次,10 min/次;加化学发光显色液后曝光。
2 结果
2.1 构建PbEMAP1 缺失疟原虫的策略 首先在pL0035 质粒耐药标签hDHFR-yFCU 两端分别插入PbEMAP1 基因编码区上、下游Up HR arm 和Down HR arm 非编码区片段,得到重组质粒pL0035-ΔPbEMAP1。基于同源重组的原理,疟原虫基因组中EMAP1 被质粒上的耐药标签替换,即可通过药物筛选获得PbEMAP1 敲除的疟原虫(图3)。
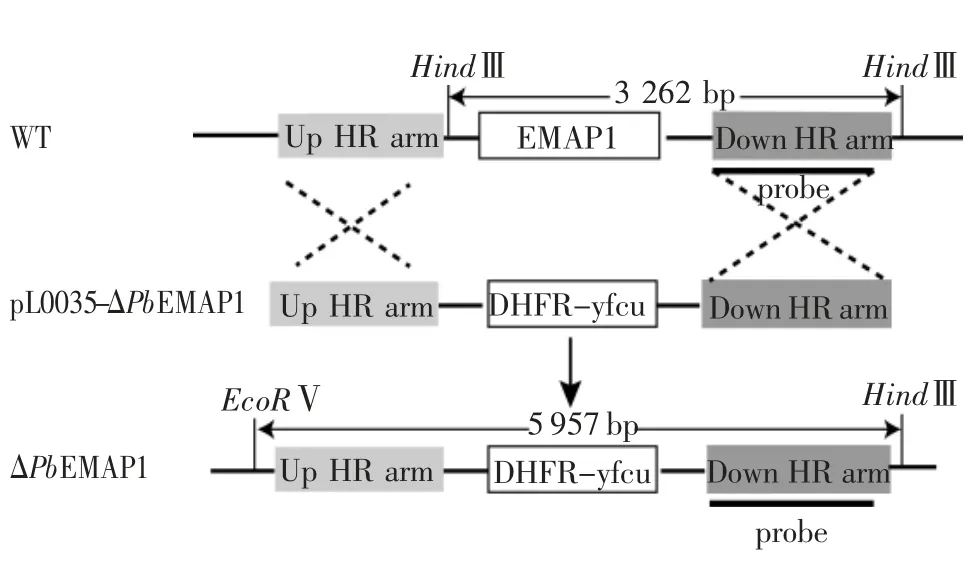
图3 伯氏疟原虫PbEMAP1 敲除策略Fig 3 Strategy of PbEMAP1 knockout in Plasmodium berghei
2.2 重组质粒pL0035-ΔPbEMAP1 的构建和鉴定 重组质粒pL0035-ΔPbEMAP1 的构建分为两步(图4A)。第一步,以WT 疟原虫基因组DNA 为模板,PCR 扩增Down HR arm,同时引入KpnⅠ和EcoRⅠ两个酶切位点,PCR 产物胶回收(图4B 左),经双酶切后纯化,与KpnⅠ和EcoRⅠ酶切处理的pL0035质粒在T4 连接酶的作用下连接、转化,从而将Down HR arm 插入pL0035,得到质粒pL0035-Down HR arm,酶切鉴定显示克隆②和⑥成功插入目的片段(图4B 右),测序正确。第二步,以WT 疟原虫基因组DNA 为模板扩增Up HR arm,并按照无缝克隆的要求在两段引入overlap 序列(图4C 左);将经SacⅡ酶切的pL0035-Down HR arm 通过HiFi DNA Assembly 试剂盒插入Up HR arm 片段,转化后得到重组质粒pL0035-ΔPbEMAP1,经SacⅡ酶切鉴定插入片段大小正确(图4C 右),测序无误。

图4 重组质粒pL0035-ΔPbEMAP1 的构建和鉴定Fig 4 Construction and identification of recombinant plasmid pL0035-ΔPbEMAP1
2.3 伯氏疟原虫转染子基因型鉴定 对转染后的疟原虫进行乙胺嘧啶筛选,获得重组疟原虫杂合子,进一步经单克隆筛选,最终获得突变虫株C4。提取C4 疟原虫基因组DNA 进行PCR 初步鉴定(表2),结果显示C4 仅有重组子条带而无野生型条带(图5A)。Southern 印迹对C4 疟原虫的基因型进行确证,WT 和ΔPbEMAP1 疟原虫基因组DNA 经EcoRⅤ和HindⅢ酶切后,探针所检测片段大小分别为3 262 bp和5 975 bp(图3)。结果显示C4 为PbEMAP1 缺失的单克隆虫株,仅检测到6 kb 的条带,而无3 kb 条带(图5B)。

图5 单克隆疟原虫ΔPbEMAP1 基因型鉴定Fig 5 Genotype identification of single clone of ΔPbEMAP1 Plasmodium

表2 鉴定重组疟原虫基因型引物序列Tab 2 Primers of genotype identification of recombinant Plasmodium
2.4 免疫印迹法验证PbEMAP1 的敲除 提取WT和ΔPbEMAP1 疟原虫蛋白,利用制备的抗PbEMAP1 抗体检测疟原虫EMAP1 蛋白表达水平。结果显示野生型蛋白样品可检测到38 kD 目的条带,与EMAP1 蛋白预期分子量相符;ΔPbEMAP1 蛋白样品在38 kD 处未检测到信号(图6),提示制备的抗PbEMAP1 抗体可用于Western 印迹检测,PbEMAP1缺失疟原虫构建成功。

图6 免疫印迹法鉴定ΔPbEMAP1 疟原虫Fig 6 Identification of ΔPbEMAP1 Plasmodium by Western blotting
3 讨论
在感染人的恶性疟原虫中,红细胞膜蛋白-1(erythrocyte membrane protein 1,EMP1),以依赖疟原虫输出基团(Plasmodium export element,PEXEL)的方式运输到红细胞膜,促进感染红细胞与内皮细胞表面各种受体结合,如CD36、细胞间黏附分子1(intercellular adhesion molecule-1,ICAM-1)、硫酸软骨素(chondroitin sulfate A,CSA)等,从而避免脾脏的清除,导致多器官功能障碍[9-12],是疟原虫致病的主要因素。
伯氏疟原虫和恶性疟原虫(Plasmodiumfalciparum,Pf)类似,采用高度相似的转运机制将自身蛋白输送到宿主红细胞[13]。因此,Pb 可以作为研究疟原虫蛋白质输出机制以及相关表型的模型。然而,在Pb 中尚未发现PfEMP1 的同源蛋白或具有相同结构域的蛋白[14]。PfEMP1 如何运输到红细胞膜上的具体机制尚未完全清楚。PbEMAP1 和PfEMP1 一样,被运输到红细胞膜上[6]。由于Pb 和Pf 具有相似的蛋白运输机制,因此研究PbEMAP1 蛋白运输机制可能为研究PfEMP1 如何运输到红细胞膜上提供新的思路;PbEMAP1 作为分泌到红细胞膜上的主要蛋白,推测其可能同PfEMP1 一样,与疟原虫致病性相关;另外还可以将PfEMP1 和内皮细胞结合的肽段嵌合到PbEMAP1基因上,即可在体内研究PfEMP1 的具体功能,从而为研发针对PfEMP1 靶点的药物提供帮助[15]。
此外,目前构建的PbEMAP1 缺失疟原虫含有耐药标签DHFR-yFCU,该耐药标签两端分别具有一段相同的序列3′pbdhfr/ts,对该疟原虫给予5-氟胞嘧啶可进行第二次药物筛选,即发生第二次同源重组。再次重组的PbEMAP1 缺失疟原虫可因删除耐药标签而存活。在此基础上可在疟原虫常用基因组改造位点230p(PbANKA_0306000)插入PbEMAP1,用于后续的回补实验。
综上所述,本研究基于同源重组的原理获得PbEMAP1 缺失的伯氏疟原虫,从DNA 和蛋白水平对筛选的单克隆重组疟原虫进行验证,并获得抗PbEMAP1 多克隆抗体,为今后研究PbEMAP1 对疟原虫生长发育及致病力的影响、挖掘潜在的抗疟药物靶点提供了必要的分子工具。
猜你喜欢
动物医学进展(2022年5期)2022-11-25
首都食品与医药(2022年19期)2022-11-19
中国农业科学(2022年16期)2022-09-19
中国科技教育(2021年4期)2021-10-18
科学导报(2020年69期)2020-11-09
中国实用医药(2019年36期)2019-01-30
分析化学(2018年11期)2018-01-16
课程教育研究·新教师教学(2016年14期)2017-04-12
海峡科技与产业(2016年11期)2016-12-26
女性天地(2016年6期)2016-09-09
