
百脉根根瘤内根瘤菌与假单胞菌的多样性分析
2022-04-13 07:47沈红玲梁红丽达晓玉王楠罗堂亮万传星
塔里木大学学报 2022年1期
沈红玲,梁红丽,达晓玉,王楠,罗堂亮,万传星
(塔里木大学生命科学与技术学院/塔里木盆地生物资源保护利用兵团重点实验室,新疆 阿拉尔 843300)
百脉根(Lotus japonicus),隶属蔷薇目豆科(Leguminosae)百脉根属(Lotus corniculatus),多年生草本植物,别名五叶草、牛角花、都草、鸟距草。我国百脉根属共有9个种,主要产自于西北地区。百脉根喜温暖湿润气候,根系萌蘖能力强、主根深入地下、对土壤环境要求较低,能够适应多种生境条件,在固持土壤、防风固沙、植被恢复和水土保持中有重要的作用[1]。百脉根花期长,花量大,除供观赏外,因种植简便、营养丰富、适口性好,还可作为饲料资源。百脉根根系发达,并与土壤中的固氮菌结合以改善土壤肥力从而提高共生固氮效能,能够作为荒漠生态恢复的先锋物种[2]。
根瘤菌是一种能与豆科植物建立共生关系的革兰氏阴性杆状细菌[3],包括固氮根瘤菌属(Azorhizobium)、茎瘤菌属(Allorhizobium)、慢生根瘤菌属(Bradyrhizobium)、中慢生根瘤菌属(Mesorhizobium)、根瘤菌属(Rhizobium)和中华根瘤菌属(Sinorhizobium)等。在缺氮条件下,根瘤菌定殖到豆科植物的根毛细胞,感染根瘤菌诱发宿主植物细胞分化为共生器官的原基,诱导刺激植物组织分化出根瘤,开始进行共生固氮过程,主要包括信号分子的感知、信号转导、侵染线的形成和皮层细胞的分裂等,由双方相关基因共同参与、相互识别、调节的过程,在这个过程中,豆科植物为根瘤菌提供碳源,根瘤菌向植物提供铵盐和氨基酸。
根瘤是一个复杂的微环境,内生菌具有丰富的生物多样性,除根瘤菌外,发现根瘤内还存在多种内生菌,例如,内杆菌属(Endobacter)[4]、类芽孢杆菌属(Paenibacillus)[5]、伯克霍尔德菌属(Burkholderia)[6]、草螺菌属(Herbaspirillum)[7]、假单胞菌属(Pseudomonas)[8]、克雷白氏杆菌属(Klebsiella)[9]等,因培养环境与微生物所处的自然环境之间存在很大差异,大量微生物通过可培养法不能获得且无法分离得到不可培养的内生细菌,所以不能够全面反映根瘤内生菌的群落结构。免培养法为内生菌多样性分析提供了有效的途径,从而能全面客观深入地揭示目标环境中微生物群落结构,其结果不仅可以丰富对根瘤内生菌多样性的认识,还将有助于进一步加深对豆科植物根系相关微生物的认识。张爱梅等[10]对中国沙棘(Hippophae rhamnoidoes)根瘤样品运用两种方法分析了中国沙棘根瘤内共生细菌多样性;何建清等[11]对西藏砂生槐根瘤内生细菌种群组成及其促生潜力进行了研究,发现根瘤内生细菌多样性丰富,且具有良好促生能力。假单胞菌属(Pseudomonas)是一类严格好氧的革兰氏阴性杆菌,是植物根际最为丰富细菌类群之一[12],具有较强的生态适应性和根际定殖能力,能够直接促进植物生长[13]和抑制植物病原菌,主要用于生物防治与生物化肥。
目前芽孢杆菌与根瘤菌和假单胞菌互作都有相关研究,百脉根在遗传多样性与系统进化分析[14]、抗逆境胁迫[15]、根瘤固氮[16]、缩合单宁[17]、作为生物反应器[18]以及药理学应用等方向已经有很深入的研究,而在根瘤内生菌多样性和根瘤菌与假单胞菌的互作关系方面研究较少。本研究以百脉根根瘤为研究材料,将免培养法和可培养法联合使用,全面揭示百脉根根瘤内生细菌的多样性,明确根瘤内共生细菌群落结构特点及组成,并获得具有潜在应用价值的内共生细菌资源;采用平板对峙法对根瘤菌与假单胞菌的关系进行探究,以期能够发现促进植物生长的促生菌,为使用合成群落促进生物固氮提供资源。
1 材料与方法
1.1 植物根瘤样品
百脉根材料采自塔里木大学校园内(81°17′E,40°32′N),由塔里木大学生命科学与技术学院刘艳萍教授鉴定为细叶百脉根。采集根瘤样品后先用流水清洗表面,再用75%乙醇和2.5%次氯酸钠溶液分别进行表面消毒,最后用无菌水反复冲洗,收集最后一次冲洗的无菌水进行无菌检测。
1.2 主要试剂与仪器
PCR仪(Bio-Rad公司)、凝胶成像仪(Bio-Rad公司)、电泳仪(Bio-Rad公司)、微量高速离心机(Bio-Rad公司);10×Easy Taq buffer、dNTPs、Taq(全式金生物公司);蛋白酶K和溶菌酶(天根生化科技北京有限公司);细菌引物27 F和引物1492 R(生物工程上海股份有限公司)。
1.3 免培养法分析百脉根根瘤内生菌多样性
1.3.1 内生菌总DNA提取与16S rRNA V5-V7区PCR扩增
根瘤内共生细菌总DNA用试剂盒(OMEGA Soil DNA Kit)提取,同时采用Nanodrop对DNA进行定量,并通过1.2%琼脂糖凝胶电泳检测DNA提取质量。根瘤内生菌16S rRNA的V5-V7可变区的PCR扩增采用带特异性引物799 F(5′-AACMGGATTAGATACCCKG-3′)和 1193 R(5′-ACGTCATC CCCACC TTCC-3′),PCR 扩增采用25 μL扩增体系,扩增反应条件为98℃预变性2 min;98℃变性15 s,55℃退火30 s,72℃延伸30 s,25个循环;最后72℃延伸5 min。将PCR扩增回收产物进行荧光定量,在荧光定量系统(Promega QuantiFluor)上对文库进行定量,文库合格后在Novaseq测序平台上测序,测序由上海派森诺生物科技有限公司(http://www.personalbio.cn)完成。
1.3.2 免培养法数据分析
样品经过质检,文库构建,运用Illumina Navaseq测序平台进行分析,根据Barcode序列拆分样本,使用DADA2进行去引物,过滤,去噪,拼接和去嵌合体获得优质序列,然后使用特征分类器根据sklearn naïve Bayes算法将优质序列归属到对应的ASVs(amplicon sequence variants)并与 Silva数据库(Release132 http://www.arb-silva.de)[19]进行比对获得物种分类信息。
1.4 可培养法分析根瘤内生细菌多样性
1.4.1 内生细菌的分离
选取粒大、饱满的根瘤,在无菌条件下,用镊子将根瘤压碎,挤出汁液,用灭菌的生理盐水稀释至10-4、10-5、10-6梯度,每个梯度10 个重复,涂布在含刚果红(终浓度25 ppm)的YMA,NA,R2A,1/2 LA固体培养基上28℃培养,长出菌落后采用划线分离法多次转至YMA培养基中获得单菌落,观察菌的形态、大小、透明度、粘稠度、颜色等,直到纯化为止。
1.4.2 内生细菌的鉴定
采用碱解法[20]提取内共生细菌DNA。16S RNA基因扩增通用引物27 F(5′-ACACTTTCAT CCTCCC TCAC-3′)和 1492 R(5′-CCCTACCTTCTTA CCACTT-3′)。PCR反应总体系为25 μL:引物各0.5 μL,10×Easy Taq buffer 0.5 μL,模板3 μL,dNTPs 1 μL,Taq 0.2 μL,加ddH2O至25 μL。PCR扩增反应条件为95℃预变性5 min;95℃变性1 min,56℃退火30 s,72℃延伸2 min,30个循环;最后72℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测后,送派森诺生物科技有限公司进行测序,将菌株的测序结果上传至EzBioCloud数据库进行相似性比对。
1.5 可培养根瘤菌系统发育树的构建
基于EzBioCloud和NCBI数据库比对结果,下载数据库中与分离菌株最相似的标准菌16S rRNA序列,分离获得的11种根瘤菌如下:新泻氨基杆菌(Aminobacter niigataensis)TRM95017、纤维素根瘤菌(Rhizobium cellulosilyticum)TRM95031、甘薯根瘤菌(Rhizobium ipomoeae)TRM95046、罗氏根瘤菌(Rhizobium rosettiformans)TRM95060、葡萄汁新根瘤菌(Neorhizobium vignae)TRM95085、附著剑菌(Ensifer adhaerens)TRM95093、鸡眼草申氏杆菌(Shinella kummerowiae)TRM95114、谷粒申氏杆菌(Shinella granuli)TRM95115、动胶菌样申氏杆菌(Shinella zoogloeoides)TRM95116、中慢生根瘤菌(Mesorhizobium sanjuanii)TRM95624、戈壁中慢生根瘤菌(Me-sorhizobium gobiense)TRM9524。将所有分离菌株及其最相近标准菌的16S rRNA序列整合到一个fasta文件中用于后续的进化分析。使用邻接法(Neighbour-Joining)对菌株进行同源性比较,构建菌株的系统发育进化树。
1.6 结瘤根瘤菌的筛选
选颗粒饱满、大小均匀的百脉根种子,先用75%乙醇处理30 s,再用2.5%次氯酸钠溶液消毒3min,最后用无菌水洗涤8~10次,均匀铺于0.8%水琼脂平板上,28℃正置催芽2~3 d。催芽后的种子播种于灭菌无氮营养培养基的组培瓶中,每瓶1颗,等到植物长出第一片子叶后,根系处接种200µL根瘤菌菌液,光照室温度30℃,光照16 h,黑暗8 h,光照室培养25~30 d,不定期观察幼苗的长势及结瘤情况。
1.7 平板对峙法分析根瘤菌与假单胞菌关系
采用平板对峙法将11种根瘤菌和15种假单胞菌在YMA固体培养基上以不同距离(0.3 cm或0.6 cm)共培养4 d,不定期观察根瘤菌和假单胞菌菌落生长情况及两者之间关系,详细的操作参照HAN Q等[21]方法。
2 结果与分析
2.1 免培养法分析百脉根根瘤内生菌结构组成
采用Novaseq测序平台进行测序,得到107 322条原始数据,拼接和质控后进行嵌合体过滤,得到可用于后续分析的有效序列74 076条。在门和属分类单元进行物种分类统计,各分类单元相对丰度前10的物种(图1A)。在门分类单元水平,百脉根根瘤内生菌主要属于变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、蓝细菌门(Cyanobacteria)、厚髌骨细菌门(Patescibacteria)、疣微菌门(Verrucomicrobia)、厚壁菌门(Firmicutes)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)和芽单胞菌门(Gemmatimonadetes),其中变形菌门、拟杆菌门和放线菌门为优势门。在纲水平上(图1B),α-变形菌纲(Alphaproteobacteria)是丰度最高的纲,占总菌的80.22%,而芽孢杆菌纲(Bacilli)、γ-变形菌纲(Gammaproteobacteria)和放线菌(ctinobacteria)分别占总菌的9.97%、7.63%和0.93%。
在属分类单元水平(图1C),百脉根根瘤内共生细菌主要包括中慢生根瘤菌属(Mesorhizobium)、金黄杆菌属(Chryseobacterium)、假单胞菌属(Pseudomonas)、氨基杆菌属(Aminobacter)、茎根瘤菌属-新根瘤菌属-副根瘤菌属-根瘤菌属(Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium)、剑菌属(Ensifer)、鞘氨醇单胞菌属(Sphingomonas)、申氏杆属(Shinella)、新鞘脂菌属(Novosphingobium),中慢生根瘤菌属、金黄杆菌属和假单胞菌属为百脉根根瘤内的优势类群。
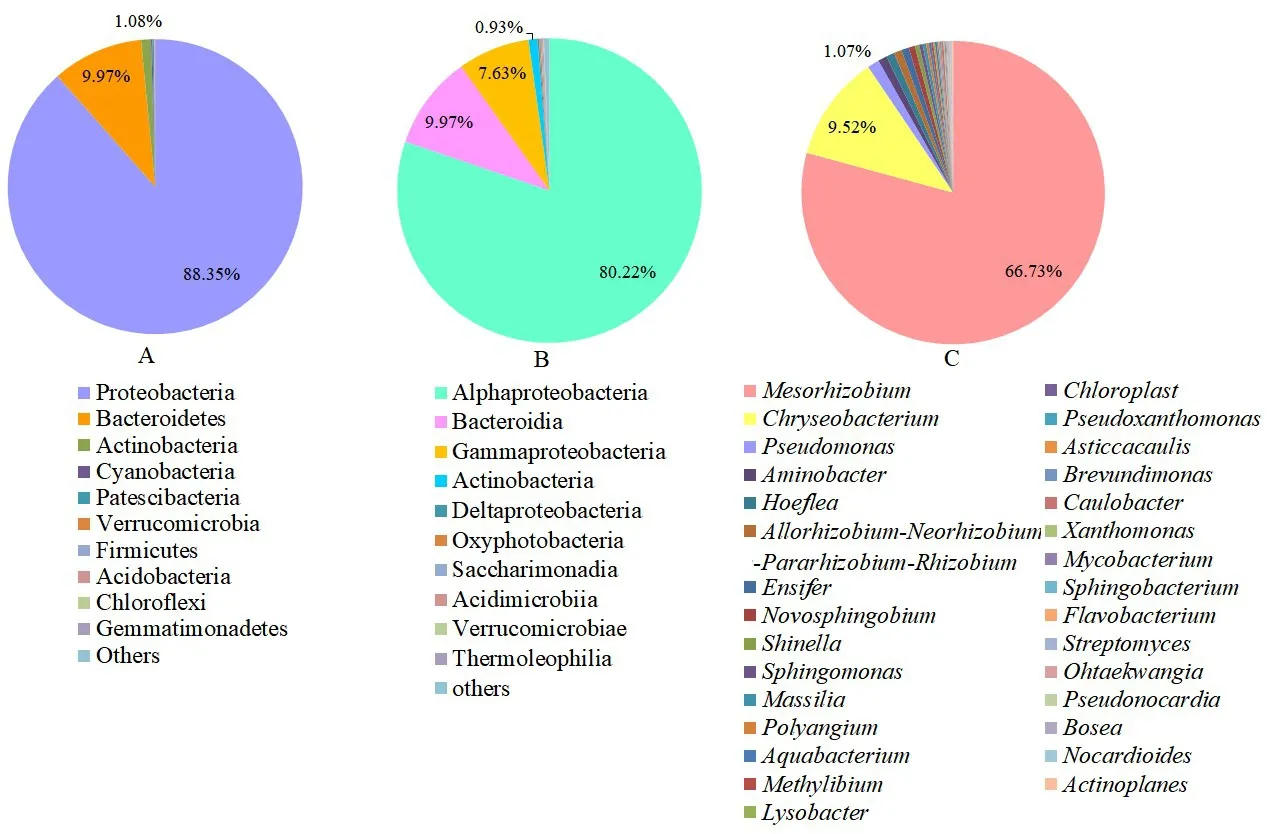
图1 百脉根根瘤免培养法细菌群落组成
2.2 可培养法分析百脉根根瘤内生细菌结构组成
本研究利用不同培养基从百脉根根瘤中分离获得278株内生细菌,经16S rRNA基因测序鉴定,这278株内生细菌分属于3门4纲12目17科28属58种。门水平上(图2D),可培养法从百脉根根瘤内分离的内生菌分属于变形菌门(Proteobacteria)为主导类群(168株,61.51%),其次是厚壁菌门(Firmicutes)(90株,27.34%)和放线菌门(Actinobacteria)(29株,9.71%)。纲水平上(图2E),γ-变形菌纲(Gammaproteobacteria)是丰度最高的纲,占总菌的32.73%,而芽孢杆菌纲(Bacilli)、α-变形菌纲(Alphaproteobacteria)和放线菌纲,分别占总菌的28.77%、28.77%和9.71%。同时,α-变形菌纲共包含11个属,γ-变形菌纲包含8个属,放线菌纲包含6个属,芽孢杆菌纲包含3个属。
属水平上(图2F),278株菌株分属于28个属,主要有:芽孢杆菌属(Bacillus)54株、假单胞菌(Pseudomonas)42株、根瘤菌属(Rhizobium)29株、申氏杆属(Shinella)20株、纤维菌属寡养单胞菌属(Stenotrophomonas)20株、纤维菌属(Cellulosimicrobium)12株、氨基杆菌属(Aminobacter)14株、微杆菌属(Microbacterium)10株、中慢生根瘤菌属(Mesorhizobium)5株,根瘤菌、芽孢杆菌和假单胞菌为优势种群,其中根瘤菌属有新泻氨基杆菌(Aminobacter niigataensis)、纤维素根瘤菌(Rhizobium cellulosilyticum)、甘薯根瘤菌(Rhizobium ipomoeae)、罗氏根瘤菌(Rhizobium rosettiformans)、葡萄汁新根瘤菌(Neorhizobium vignae)、附著剑菌(Ensifer adhaerens)、鸡眼草申氏杆菌(Shinella kummerowiae)、谷粒申氏杆菌(Shinella granuli)、动胶菌样申氏杆菌(Shinella zoogloeoides)、桑娟中慢生根瘤菌(Mesorhizobium sanjuanii)、戈壁中慢生根瘤菌(Mesorhizobium gobiense);假单胞菌有鱼假单胞菌(Pseudomonas piscium)、亚硝酸假单胞菌(Pseudomonas nitrititolerans)、猴假单胞菌(Pseudomonas simiae)、阿塔卡姆假单胞菌(Pseudomonas atacamensis)、氧化砷假单胞菌(Pseudomonas arsenicoxydans)、山绿假单胞菌(Pseudomonas yamanorum),青苔假单胞菌(Pseudomonas mosselii)、湖南假单胞菌(Pseudomonas hunanensis)、门多萨假单胞菌(Pseudomonas mendocina)、西里西假单胞菌(Pseudomonas silesiensis)、恶臭假单胞菌(Pseudomonas alloputida)、温哥华假单胞菌(Pseudomonas vancouverensis)、普农假单胞菌(Pseudomonas punonensis)、米氏假单胞菌(Pseudomonas migulae)、恶性假单胞菌(Pseudomonas viciae),根瘤菌、假单胞菌和芽孢杆菌为优势菌群,这与钟宇舟等[22]关于根瘤内生细菌的研究结果一致。免培养方法分析的前10个优势属在本次分离过程中大部分都有分离到,但不是优势属,表明免培养方法分析在一定程度上可以指导可培养分析。
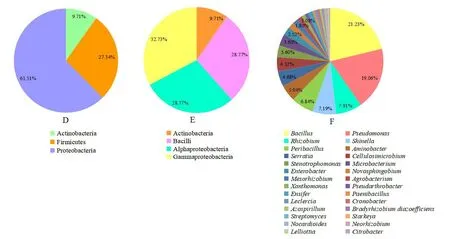
图2 百脉根根瘤可培养法细菌群落组成
2.3 可培养根瘤菌基于16S rRNA基因序列构建的系统发育树
对77株根瘤菌菌株的16S rRNA基因进行序列测定,选取11种代表性菌株利用NCBI中的Blast对测序结果进行同源性比对搜索,运用MEGA 7.0构建系统发育树。从系统发育树(图3)可知,百脉根根瘤内根瘤菌属于6个不同的属,分别为根瘤菌属、中慢生根瘤菌属、新根瘤菌属、氨基杆菌属、剑菌属和申氏杆属。

图3 可培养根瘤菌基于16S rRNA基因序列构建的Neighbour-Joining系统发育树
2.4 百脉根根瘤内生菌免培养法和可培养法多样性差异分析
在门、纲、目、科及属分类单元,免培养法共从百脉根根瘤菌内检测到内生菌分属于17门31纲69目103科140属,香农指数为5.81,辛普森指数为0.85,而可培养法只分离获得了3门4纲12目17科28属内生菌,香农指数为3.57,辛普森指数为0.84,结果表明:免培养法灵敏度较高、检测到的微生物物种数多、多样性要优于可培养法,可培养法因培养条件的限制使分离获得物种丰富度低于免培养法。
在属分类单元,运用两种方法比较了根瘤内生细菌的相对丰度,发现可培养法分离获得的内生细菌的相对丰度普遍高于其在免培养法中的丰度(图4)。可培养法分离获得的芽孢杆菌属的相对丰度为19.42%,但在免培养方法中没有检测到;相反,免培养法检测到的优势属中慢生根瘤菌属占66.73%,而可培养法只分离到1.80%,原因可能是中慢生根瘤菌生长速度较慢,培养初期生长速度较快的菌种“淹没”了中慢生根瘤菌的生长,或是中慢生根瘤菌未找到适合的培养基及培养条件。
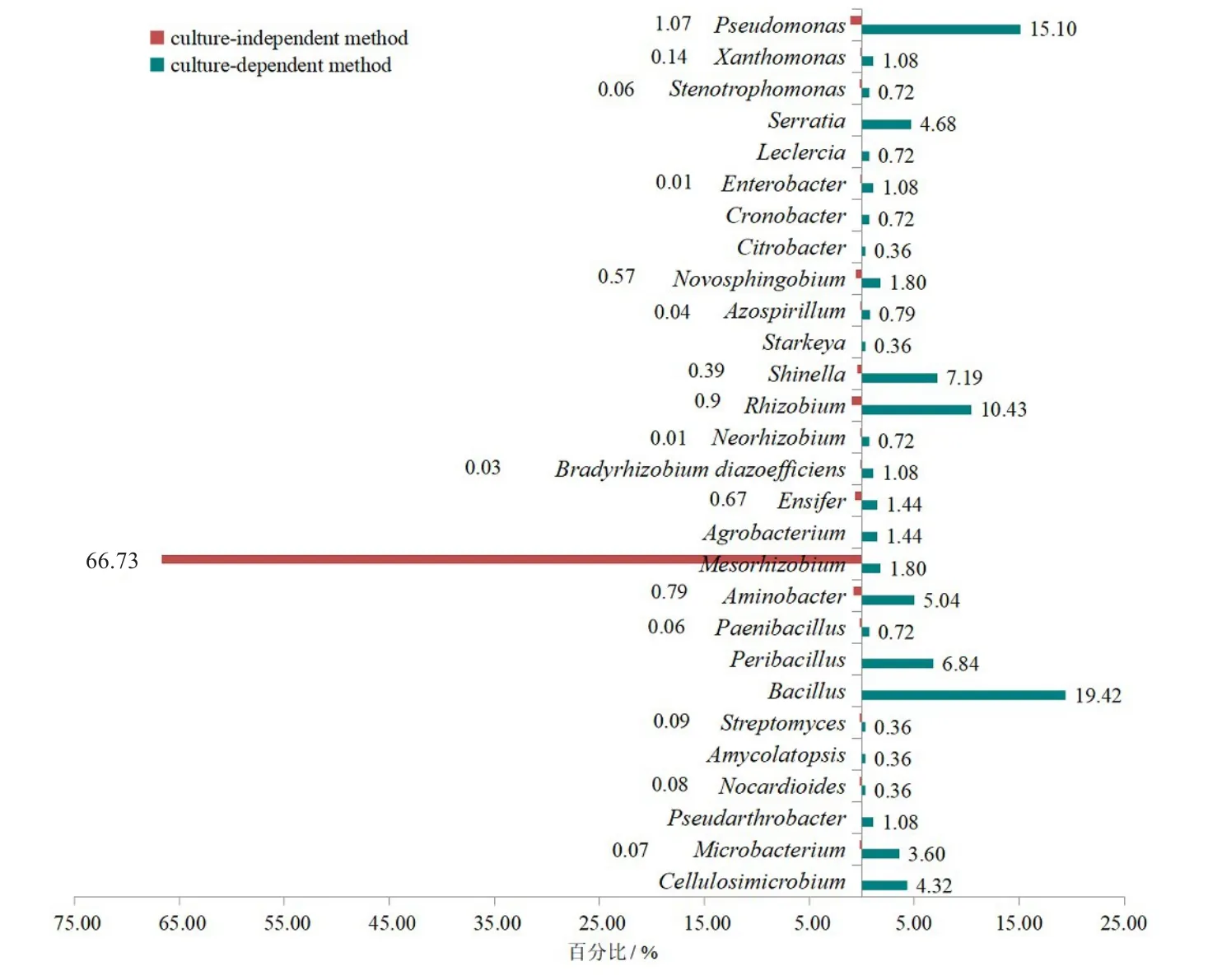
图4 免培养法与可培养法差异性分析
2.5 结瘤根瘤菌
将11种根瘤菌回接到百脉根植物根际,实验观察到,接种根瘤菌的植株长势相对较好,未接种根瘤菌的植株叶片黄,接种根瘤菌菌株TRM95017(Aminobacter niigataensis)后百脉根出现结瘤现象(图5),其余菌株侵染植物根际后无结瘤现象,表明不同菌株与同一植物匹配存在专一性。

图5 根瘤菌回接百脉根结瘤实验结果
2.6 对峙法分析根瘤菌与假单胞菌的关系
将11种根瘤菌和15种假单胞菌在YMA固体培养基上共培养4 d,观察根瘤菌和假单胞菌菌落生长情况及两者之间的关系,发现根瘤菌与假单胞菌主要出现的关系为抑制、融合共生、无作用关系,其中9种根瘤菌分别与15种假单胞菌有选择性抑制现象,抑制的假单胞菌有鱼假单胞菌(TRM95006)、亚硝酸假单胞菌(TRM95007)、猴假单胞菌(TRM95024)、阿塔卡姆假单胞菌(TRM95028)、氧化砷假单胞菌(TRM95035)、山绿假单胞菌(TRM95052)、青苔假单胞菌(TRM95084)、湖南假单胞菌(TRM95090)、门多萨假单胞菌(TRM95091)、西里西假单胞菌(TRM95100)、恶臭假单胞菌(TRM95121)、温哥华假单胞菌(TRM95122)、普农假单胞菌(TRM95125)、米氏假单胞菌(TRM95140)、恶性假单胞菌(TRM95169),说明根瘤菌与假单胞菌为满足自身生长,两者之间产生特异性的信号或菌株产生某种特殊的代谢产物来维持自身生长不受影响;8种根瘤菌分别与13种假单胞菌选择性发生融合共生关系,根瘤菌有新泻氨基杆菌(TRM95017)、纤维素根瘤菌(TRM95031)、甘薯根瘤菌(TRM95046)、罗氏根瘤菌(TRM95060)、葡萄汁新根瘤菌(RM95085)、附著剑菌(TRM95093)、中慢生根瘤菌(TRM95624)、戈壁中慢生根瘤菌(TRM95240),说明根瘤菌与假单胞菌选择性结合使两者生长的更好。还有部分根瘤菌与假单胞菌之间无特异性信号,结果见表1。
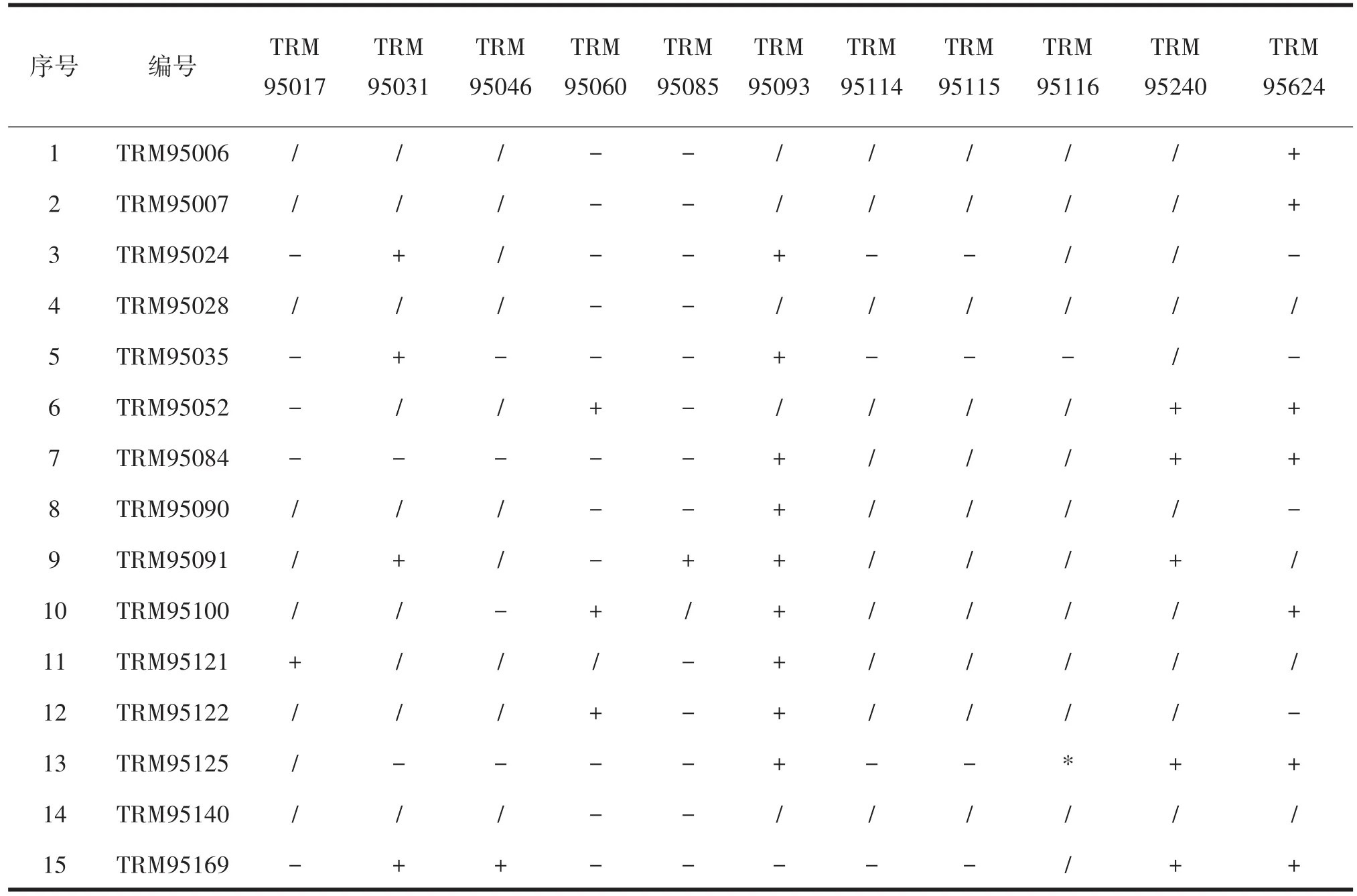
表1 根瘤菌与假单胞菌互作关系
3 讨论与结论
微生物群落结构是其整体功能的基础,对根瘤环境中有着重要的意义。本研究采用免培养法和可培养法对百脉根根瘤内生菌多样性进行了分析,免培养法检测到的优势属为中慢生根瘤菌属、金黄杆菌属和假单胞菌属等,可培养法分离获得的优势菌群为根瘤菌、芽孢杆菌和假单胞菌等,两者在细菌群落组成和分离物丰度上存在差异,原因一是免培养法自身具有优势,灵敏度比较高,检测到的微生物物种数多,二是可培养过程中存在“微生物暗物质(不可培养的微生物)”[23],三是不同细菌对不同培养基具有选择性,可培养法培养基设计的种类有限。针对可培养法的局限性丰富培养基种类和分离手段,优化差异条件,针对性设计分离方案,形成多类型的“增菌培养基”[24],或使用I Chip法原位富集一些不可培养的细菌[25],是可培养法新的策略。因可培养法的局限性,使得免培养法检测到百脉根根瘤内相对丰度较高的中慢生根瘤菌属在可培养分离过程中不是优势属,可能是微生物培养过程中,个别种类的微生物由于数量占优势或生长速度较快,在培养初期大量生长,“淹没”了生长较慢的中慢生根瘤菌。然而免培养法也不能完全代替可培养法,可培养分离获得的芽孢杆菌属在免培养法中未检测到,说明免培养法不能注释所有的微生物,只有将免培养法与可培养法相结合方才能更全面系统地揭示根瘤内细菌的多样性,挖掘潜在应用价值的微生物资源。
目前根瘤菌、芽孢杆菌和假单胞菌在结瘤固氮、促进植物生长及互作关系方面有研究,MEYER S D等[26]从30种豆科植物根瘤中分离鉴定出芽孢杆菌和假单胞菌,其他植物[27]内生菌中假单胞菌属和芽孢杆菌属等为最常见的属,本研究结果与其一致。HAN Q等[21]研究发现蜡样类芽孢杆菌组不仅能特异性地促进快生根瘤菌,抑制慢生根瘤菌的生长,还能缓解盐碱胁迫对快生根瘤菌结瘤的负面影响,增加其在根瘤中的定殖。SUN X L等[28]研究发现施用典型根际益生芽孢杆菌(Bacillus velezensis)不仅可独自促进植物生长,其代谢产物能诱导施式假单胞菌(Pseudomonas stutzeri)的显著富集,促进植物生长并协助植物耐盐性。
本研究通过免培养与可培养相结合方法揭示了百脉根根瘤内生菌多样性,免培养法检测到根瘤内前10个优势属在可培养分离中大部分都有分离到,表明免培养法在一定程度上可以指导可培养法,为后期有目的地分离特定物种奠定了基础。另外获得了1株结瘤根瘤菌和13种与根瘤菌共生的假单胞菌,为生物固氮研究储备了重要的菌种资源。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
河南医学研究(2022年19期)2022-10-19
动物营养学报(2022年8期)2022-09-01
分子诊断与治疗杂志(2022年4期)2022-05-30
新疆农业科学(2022年4期)2022-05-12
中国种业(2022年4期)2022-04-19
中国南方果树(2022年1期)2022-01-28
农技服务(2021年7期)2021-09-24
园艺与种苗(2021年1期)2021-03-16
国际消化病杂志(2021年1期)2021-03-05
