
微藻固碳光合作用强化策略及展望
2022-09-13 03:48毛炜炜尹庆蓉李鹏程宋春风
洁净煤技术 2022年9期
毛炜炜,张 磊,尹庆蓉,李鹏程,胡 毡,宋春风
(1.天津大学 环境科学与工程学院,天津 300350;2.一汽丰田汽车有限公司,天津 300350)
0 引 言
CO2排放量急剧上升导致全球气候变暖,成为人类迄今为止面临的最严峻的环境挑战之一[1]。2022年3月国际能源署(IEA)发表的最新报告显示,截至2021年,全球CO2排放量已增至363亿t,达到历史最高水平[2]。大气中CO2浓度增加引起地表温度持续升高,对自然环境造成不良影响,如海平面上升、冰川融化等,这些变化导致极端天气事件发生、生物物种变化[3]。如何实现CO2减排已成为当前国内外关注的焦点。
CO2捕集利用与封存技术(CCUS)作为一种重要的温室气体减排技术,是我国构建生态文明和实现可持续发展的重要技术,对实现碳达峰、碳中和目标至关重要[4]。目前已有多种碳捕获方法,如化学吸收、吸附、膜分离、微藻固碳法等。从可持续发展角度来看,微藻介导的CCUS技术由于能吸收CO2并将其转化为生物质能源,已成为现阶段资源化利用CO2和开发新能源的热门课题[5-6]。与此同时,利用微藻固碳作为一种经济可行、环保、可持续的固碳技术,应用潜力巨大[7]。
微藻由真核微藻(如硅藻、绿藻等)和原核微藻(如蓝藻等)组成,可以利用太阳能,通过光合作用将CO2转化为蛋白质、淀粉、脂类等生物质。与传统的碳捕获方法相比,微藻固碳主要有以下优势:① 能直接利用太阳能,相比于物理和化学法,节省大量能源。② 微藻固碳过程环境友好,代谢物中无二次污染物。③ 微藻生长周期短(4~10 d),光合效率高,是一般陆生植物的10~50倍[8]。④ 微藻是水生生物,环境适应性强,可在沿海泥滩、盐碱地和沙漠中栽培,不与陆生植物争夺土壤。我国拥有广阔的海洋和泥滩等可培养微藻进行资源化利用,不但能减少大气中CO2,还能带动当地经济发展[9]。⑤ 微藻生物质中50%左右都是碳,其生长可消耗大量CO2,实现固定和转化发电厂或其他工业废气中CO2的目的[10]。且城市废水和工农业废水可作为培养微藻的营养源,降低成本[11-12]。⑥ 微藻生物质含有丰富的脂类和碳水化合物等成分,可生产高附加值生物质产品。这些产品可用于制备食品、动物和水产养殖饲料、化妆品、药品、肥料、保健品、具有特殊用途的生物活性物质和生物燃料(包括生物柴油、生物氢、染料油、乙醇等)[13]。而其转化制备的生物燃料被认为是极具潜力替代石油的生物质资源之一[14]。
微藻捕获CO2的效率主要取决于微藻种类、培养系统和生长条件(CO2浓度、培养基、pH、温度、光照强度、溶解氧)。常用于捕获CO2的微藻种类包括螺旋藻、小球藻、栅藻、葡萄藻、裸藻、绿球藻等。研究表明,在高浓度CO2条件下养殖微藻,会大幅提高微藻的固碳效率和生物质产量,还会增加脂类积累。RODAS-ZULUAGA等[15]发现随CO2体积分数增加(0.03%~20.00%),Scenedesmussp.的生物量和蛋白质随之增加。光是微藻生长和代谢的基本条件之一,为光合作用和一系列生理反应提供动力,然而微藻的光合作用效率并不随光强度或光照时间的增加而增加。通过控制适当的光强度和照射时间,可提高微藻细胞的光利用效率,如12 h∶12 h或18 h∶6 h的光照-黑暗时间。培养基应有足够的营养物质来提供微藻生长,C、N、P和S是微藻细胞的重要成分。此外,培养基成分不同会使微藻产生不同的目标产物,可通过改变微藻培养条件的手段调控。
2050年能源科学发展战略已明确将微藻固碳和能源利用作为中国的潜在发展方向,但利用微藻对CO2进行捕获仍面临一些挑战。笔者从影响微藻光合作用效率的主要因素出发,介绍了微藻的光合作用和固碳机理,回顾微藻固碳的研究现状并探讨了其研究制约因素,总结提高微藻光合效率的策略,包括诱变、驯化、基因工程以及纳米材料的干预等,最后,对未来微藻固碳领域的发展进行展望。
1 微藻光合作用固碳
1.1 光合作用
微藻光合作用利用光能将CO2转化为有机化合物同时释放分子氧,是微藻固定和储存CO2的基础[16]。研究表明,微藻的光合效率高达12%,是固定CO2的首选[17]。除固碳外,微藻还能合成不同生物分子,如蛋白质、碳水化合物和脂质等[18]。通常,光合作用的整体反应用式(1)描述:
(1)
真核微藻的光合作用发生在叶绿体中,但原核微藻蓝藻发生在类囊体中,这是因为原核生物中缺乏叶绿体细胞器[19]。光捕获天线复合物由光捕获色素组成,在光合作用过程中具有多种功能,其捕获光能并转移到光合反应中心以触发有氧光合作用[20]。这些天线系统将吸收的光子能量传递给叶绿素分子,并以叶绿素形式保存能量,从而为光合作用提供动力[21]。微藻细胞中的光合作用分2个阶段进行:光反应和暗反应阶段。光合作用的第1阶段光反应由光驱动,利用光能将氧化态烟酰胺腺嘌呤二核苷酸磷酸(NADP+)和二磷酸腺苷(ADP)转化为储存能量的烟酰胺腺嘌呤二核苷酸磷酸(NADPH)和三磷酸腺苷(ATP)分子[22]。第2阶段暗反应不需光参与,且与第1阶段光反应同时发生,微藻细胞不断消耗光反应过程中产生的NADPH和ATP固定大气中CO2并形成碳水化合物[22]。
光合作用的核心是2个主要的光活性复合物(光系统I(PSI)和光系统II(PSII)),通过激发的叶绿素二聚体将阳光转移到电子传输链中。通过激发叶绿素P680驱动电子传递二聚体进入PSI。此外,PSI通过叶绿素P700转移从PSII接受的电子,叶绿素从光激发的天线叶绿素氧化成强烈还原的NADPH[23]。同时,可通过光磷酸化过程中形成ATP储存光反应收集的能量,每对通过光合电子传递链的电子可形成1.3个ATP分子[24]。
暗反应中,藻类通过卡尔文循环(图1)固定CO2,其中核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)在CO2固定中发挥关键作用[25]。Rubisco是双功能酶,具有催化2种不同反应的能力。① 羧化反应中,利用CO2和核酮糖-1,5-二磷酸(RuBP)形成2个分子的3-磷酸甘油酸(3-PGA)。3-PGA被用于卡尔文循环中RuBP再生,并为微藻进一步碳代谢提供受体。3-PGA通过微藻中1,7-二磷酸景天庚酮糖蛋白(SBPase蛋白)调控。② Rubisco可作为加氧酶引起光呼吸,此时Rubisco利用O2和RuBP产生2-磷酸乙醇酸酯(2-PGA)和3-PGA。2-PGA进一步转化为乙醇酸盐。后者通过C2循环、甘油酸途径或通过脱羧作用代谢[26]。
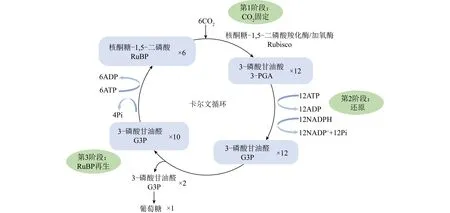
图1 卡尔文循环反应Fig.1 Reactions during the Calvin cycle
1.2 微藻中无机碳的跨膜形式

1.3 微藻种类的影响
微藻种类直接影响CO2固定和生物质产量。典型微藻种类对生物质产量和最大CO2固定率参数的影响以及最佳CO2浓度见表1。
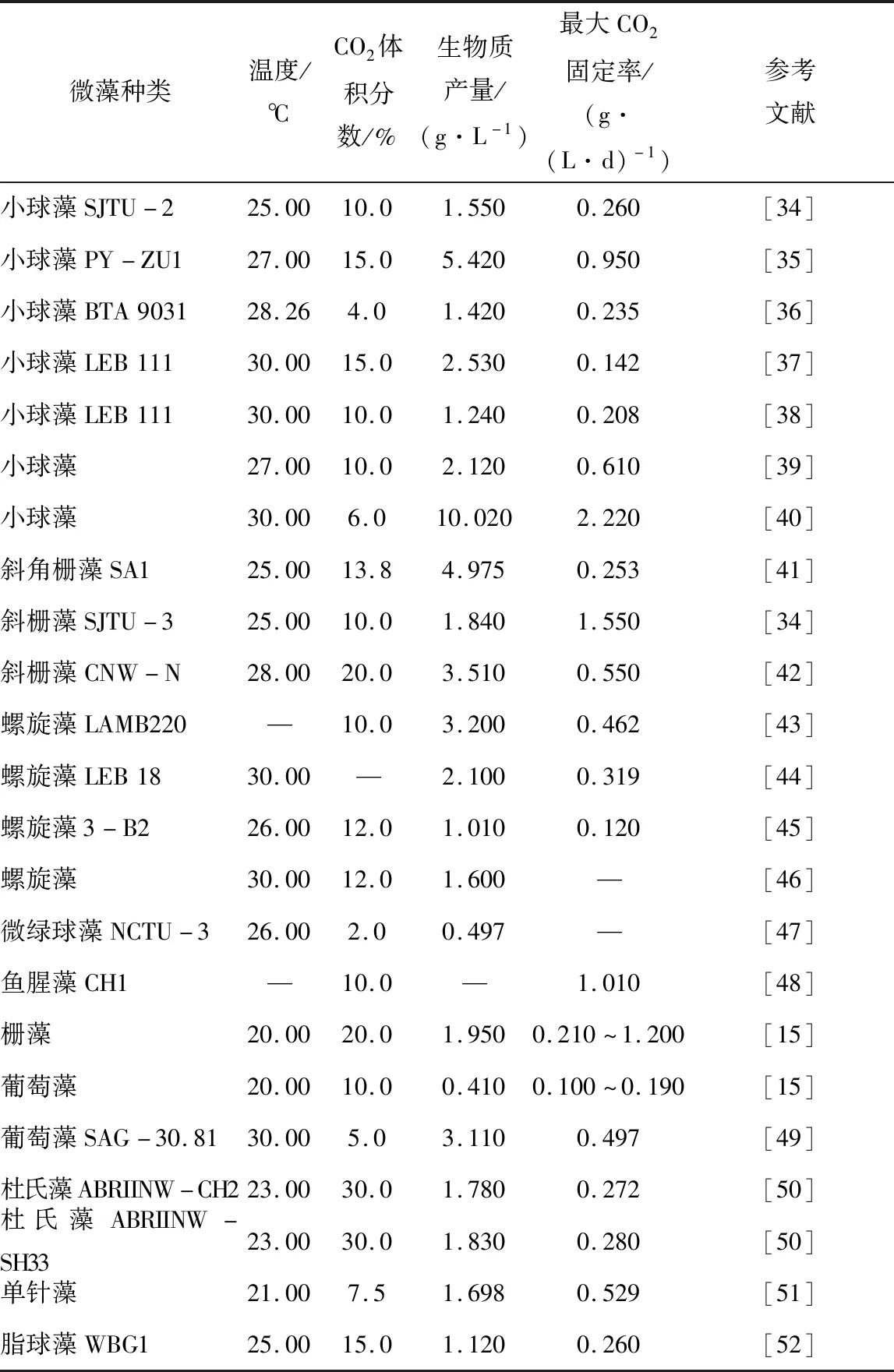
表1 不同种类下微藻生物质产量和最大CO2固定率Table 1 Microalgal biomass yield and maximum CO2 fixation rates under different species
CO2固定和生物质生产性能因微藻种类而异,但由于微藻具有不同的生物学性能并在不同条件下培养,因此数据可能不具有严格的可比性。然而,与其他物种相比,观察到小球藻性能更好,生长速度快、更耐受恶劣环境。其生物质产量和CO2固定率分别高达10.02 g/L和2.22 g/(L·d)[40]。
微藻-CO2生物固定和生物质生产是一个复杂的物理化学过程。实现高CO2固定率和生物质产量的最佳范围或最佳值因微藻物种不同而存在差异。有必要开发高性能物种,在更复杂和恶劣的环境中实现微藻CO2固定和生物质生产最大化。因此,选择、分离和培养生长速度快、光合速率高、环境耐受/适应性强、脂质含量高的微藻物种将是未来研究方向[27]。
2 微藻光合效率提高策略
2.1 随机诱变
2.1.1 化学诱变
化学诱变是突变育种领域一种高效易行的技术,通过化学诱变剂处理生物细胞,以诱发基因突变产生遗传性状的改变。根据育种目标对产生的突变体进行筛选培育,最终得到具有理想特征的新品种,如高CO2固定率、高脂质含量、强耐酸性等,具有很高专一性。目前最广泛使用的化学诱变剂包括甲基磺酸乙酯(EMS)和亚硝基胍(NTG)[53-54]。
KAO等[55]使用EMS诱导小球藻得到MM-2突变体。MM-2在生物量质量浓度为1.2 g/L时,对脱硫沼气(~20% CO2、~70% CH4和H2S体积分数<100×10-6)的CO2捕获效率在阴天约为70%,晴天为80%。研究发现,采用EMS诱变还可增加微藻的生物质产量和脂质积累。TANADUL等[56]使用EMS诱导小球藻,通过除草剂筛选出突变体,相比于野生型,其最高生物质产量和脂类产量分别提高了110%和53%。
NTG是另一种常见的化学诱变剂,LEE等[57]使用N-甲-N′-硝基-N-亚硝基胍进行诱变,成功获得了一种具有耐高温的小球藻GD突变体。该突变体在室外高温条件下具有较高的光合作用效率和生物质产量,是大规模生产乳酸的有效发酵原料。KUO等[58]通过NTG诱变法筛选了一种耐碱的小球藻突变体AT1,结果表明,通过10% CO2的间歇性通气和碱性培养基(pH=10)作为耐碱微藻培养的CO2捕获策略,可以明显提高小球藻突变体AT1的CO2利用效率。
化学诱变可产生高频率的点突变,染色体畸变率相对较低,有利于选择特定的育种方向[59]。但化学试剂对微藻的毒性较大,也存在突变频率低等问题,且使用过程中有一定毒害性。
2.1.2 紫外线诱变
紫外线诱变控制灵活、操作方便,被广泛应用于包括藻类在内的各种生物体的诱变[60]。紫外线辐射能诱导DNA中嘧啶二聚体的形成,包括转变、颠换、移码突变或缺失,从而引起生物体突变[61]。
通过紫外诱变得到的诱变藻株是CO2固定和生产生物燃料的候选藻种,有助于减少全球变暖和缓解能源短缺。LI等[62]利用紫外线诱变技术能耐受高浓度CO2的高效微藻S.obliquusWUST4,并利用其开发的中试规模系统从实际烟气中捕获CO2,CO2去除率可达67%。LIU等[60]利用紫外诱变产生小球藻突变体,相比于野生型,其生物量和脂质均有不同程度增加,生长速率更高。
QI等[63]结合96孔微孔板-紫外诱变技术,筛选出一种斜栅藻突变体SDEC-1M。连续培养后,该突变体表现出高CO2耐受性和高生物质产量。此外,高光转换效率和高碳水化合物含量说明其能将光有效转化为生物乙醇原料。这个创新方法使每个细胞在96孔微孔板的孔中分离诱变,保护了分离藻株纯度,避免了巨大的工作量和感染风险。
紫外线辐射引起的突变为非定向,但可灵活控制使微藻发生突变[64],不需获取微藻的遗传信息。与基因工程相比更易操作;与化学方法相比,紫外诱变可避免二次污染。由于紫外线突变对生物体有显著生物学效应,如具有更好的固碳性、脂质、虾青素等,可获得更高收益[60]。
2.1.3 核诱变
核诱变能通过电离作用改变DNA分子结构,直接导致碱基、脱氧核糖、糖-磷酸连接处的化学键断裂;且在电离的同时与细胞中分子,特别是与水分子相互作用,产生大量自由基破坏DNA结构引起基因突变或重组,从而改变物种的遗传性状[65]。AGARWAL等[66]发现γ射线能在一定程度上刺激Rubisco酶的活性,促进了卡尔文循环中C3化合物的合成,从而提高CO2捕获效率。
60Co和137Cs是最常用于核诱变的辐射源。ZHU等[67]用60Co的γ辐照获得了耐酚且生长迅速的A.platensis突变体,突变体的苯酚降解效率和CO2固定率相比野生型分别提高了34.7%和22.4%。CHENG等[68]采用60Co的γ射线照射螺旋藻以提高CO2固定率和生物质产量,其得到的突变体生物量相比于野生型增加了310%,认为CO2固定率的提高是由于螺旋藻突变体微观结构的变化[68]。突变体的细胞壁孔径增加,这一特性降低了孔阻力,直接促进CO2向细胞中扩散[68]。此外,CHENG等[69]通过γ射线对菱形藻Nitzschiasp.进行了2轮诱变,发现突变体快速生长,培养周期由15 d缩短至12 d。LU等[70]应用137Cs射线照射小球藻,得到的突变体MS700吸收溶解无机碳能力和析氧速率比野生型分别高出27.7%和31.4%。突变体的生物质产量和固碳效率大幅提高,其可能原因是基于叶绿素的闪氧产率升高,故质体醌库中有更多的可用电子受体,光能到化学能的转换效率更高[70]。
2.2 适应性实验室进化
适应性实验室进化(ALE)即驯化,是一种改善微藻菌株特性的有力工具,用于在环境压力下(如高CO2浓度、高盐度、高温等)培养微藻,以获得具有理想表型的微生物进化[71]。借助压力驯化,培养物中大量微生物可通过多周期培养直接在多种基因中平行积累非直观的有益突变[72]。工业烟气的主要成分是CO2,同时还包含NOx、SOx、PM2.5、重金属Hg等多种残留物。YE等[73]采用生命周期法综合评价了微藻固定燃煤电厂和煤化工烟气CO2技术的能量转换特性和环境影响。结果表明,微藻生长速率变化对能源消耗和净CO2排放影响最大。因此更多学者采用驯化微藻提高其在烟气条件下的生长速率和耐受性[59]。
ALE是提高小球藻对高CO2耐受性的有效途径[74]。为获得高CO2耐受性且具有高CO2固定能力的微藻菌株,LI等[74]在10%和20%的CO2条件下进行了31个周期的适应性实验室进化,分别获得了2个进化的小球藻AE10和AE20,均能在30%的CO2下快速生长,且最大生物量浓度和叶绿素含量明显高于原始藻株。WANG等[10]对海洋微拟球藻CCMP1779进行了CO2逐步梯度驯化,在15%的CO2下,驯化后的微藻生物量提高了22.7倍,同时研究了CO2梯度驯化改善微藻特性的机制。研究发现微藻的功能选择性多靶点基因突变由CO2梯度驯化过程引起,从而使光合作用、碳固定和细胞合成能力提高[10]。
除了高浓度CO2,烟气中其他杂质也会抑制微藻生长。ASLAM等[75]研究证明了混合微藻群落可逐渐在燃煤产生的100%未经过滤烟气中(含11%CO2)生长。然而,由于适应高浓度SOx和NOx需较长时间,只有添加更高浓度的磷酸盐缓冲剂确保pH才能维持生长。为进一步研究杂质对适应性实验室进化的影响,CHENG等[76]通过适应性实验室进化(46个周期)获得了一株新的小球藻Cv。证实了该小球藻能在模拟烟气(10% CO2、200×10-6NOx和100×10-6SOx)条件下适应生长,最大CO2固定率为1.2 g/(L·d)[76]。
CO2梯度驯化是工业上培养各种高CO2耐受微藻的理想方法。经过驯化的微藻既能快速生长,又能提高固定CO2的能力。但烟气中存在的SOx和NOx抑制微藻的CO2固定过程,故能在微藻生长过程中保持稳定pH,这对于利用微藻处理高浓度SOx和NOx的烟气至关重要[77]。
2.3 基因工程
近10 a来,微藻基因组学的重大进展为基因工程提供了坚实基础[78]。一些微藻的细胞器基因组已被测序,并且证明了微藻基因型和表型之间的内在关系[79-80]。由于基因编辑技术不断发展,基因组可根据需要进行精确编辑。微藻基因工程有利于改善微藻的光合作用和生物量积累,从而提高CO2捕获率[78]。最大化光合作用效率是目前开发和利用生物能源的最大挑战之一。
光合作用涉及光吸收、能量转换和碳转移等,是一个复杂的物理化学反应过程[72,81]。改善光合作用对于提高生物质生产力和CO2封存效率至关重要。改善微藻光合作用效率的3个主要潜在目标点为:① 提高参与CO2固定的酶效率[82];② 改变能量收集复合物[83];③ 增加CO2同化以减少CO2和能量损失[79,84-85]。
2.3.1 CO2固定中关键酶改进
参与CO2固定途径的关键酶表达调控是提高CO2固定效率的关键手段之一。CO2固定涉及2个关键途径:卡尔文循环(将无机碳固定在有机碳上)和CCM(促进无机碳向卡尔文循环的输送)[86]。
Rubisco是全球碳循环的主要参与者,涉及卡尔文循环中碳固定的第1步,引导CO2进入卡尔文循环[87]。但Rubisco酶的催化反应是CO2固定的限速步骤,因为在正常CO2水平下,Rubisco仅发挥了其25%的催化能力。为增加Rubisco活性,有研究集中在Rubisco基因改造,以提高其速度和选择性[88]。增强Rubisco的催化特性可通过引入点突变进行[89]。FUJIHASHI等[90]根据中温菌(来自植物I型)和超嗜热古菌(III型)Rubiscos的三维结构和氨基酸残基的差异,设计了几个新的点突变增强其在室温条件下的活性。点突变SP8-T289D型在体内活性的改善尤其明显,其生长率几乎是环境温度下含野生型酶菌株的2倍。还可通过修饰Rubisco激活酶基因的表达来调节Rubisco活性。WEI等[82]通过核编码的、与cbbX同源的Rubisco激活酶的过度表达,提高了工业油料微藻Nannochloropsis-oceanica的生物质产量和光合作用效率。在空气水平的CO2下,突变体的生长速率、生物量产量和脂质产量分别提高了约32%、46%和41%,光合作用提高约28%。其中Rubisco大亚基蛋白丰度增加约45%,表明nNoRca样的过度表达通过上调Rubisco水平和活性来增强微藻的光合作用。
此外,CO2光合固定效率不仅取决于Rubisco的羧化能力,还取决于RuBP的再生能力。醛缩酶过表达促进卡尔文循环中的RuBP再生和光系统中的能量转移。YANG等[91-92]产生了表达蓝藻醛缩酶的转基因C.vulgaris细胞。由于醛缩酶的过表达,转基因微藻的CO2固定率和能量转移效率更高。此外,许多研究报道在微藻中过表达SBPase基因和Rubisco激活酶基因会加速卡尔文循环,从而增强了碳固定和光合作用的能力。
2.3.2 能量收集复合体的优化
光照是直接影响微藻的光合活性及其生长动力学的主要因素[96]。微藻的光照辐射增强时,其光转换效率和CO2固定能力提高[97]。然而,约80%的吸收光子可能浪费,从而将光合生产力和太阳能到产品的转换效率降至极低水平[98]。
通过减小集光复合体(LHC)天线尺寸在一定程度上解决这些问题,减轻对太阳光的过度吸收和激发能量的浪费,从而提高光合作用效率,特别是在密集培养中[79]。一些功能性转基因插入和表达到微藻基因组中,如lciA和nabI*,可减小LHC天线尺寸,并改善CO2的固定和光合效率[99]。BECKMANN等[100]使用NAB1的永久活性变体(一种细胞溶质RNA结合蛋白)进行翻译抑制试验,降低了LHC基因的转录表达,导致PSI天线尺寸减小10%~17%。MUSSGNUG等[101]使用RNAi技术下调了莱茵衣藻中整个LHC基因家族,以减少荧光和热量造成的能量损失,从而提高了强光条件下的光合效率。结果表明,在荧光水平降低的情况下存在突变体Stm3LR3,光合量子产率更高,对光抑制的敏感性降低,提高了强光条件下的光合效率。近年来,通过敲除编码LHC蛋白质的基因产生了几种突变体,如基因tla1、tla2和tla3,这些突变体减小了叶绿素天线尺寸。如敲除tla3(CpSRP43)提高了微藻的太阳能转化效率,由于将天线尺寸减小至野生型的40%,从而提高了光合效率[102]。
此外,调整降低非光化学淬灭系数(NPQ)机制也可提高作物生产力[103]。PEROZENI等[104]发现LHCSR3蛋白在莱茵衣藻PSI和PSII的NPQ机制中发挥重要作用。LHCSR3在HSP70/RBCS2启动子的控制下表达,为细胞提供最小的NPQ容量。这种微调的NPQ活性将莱茵衣藻的光合效率从3.4%提高至4.4%~4.7%[105]。最近,SPA1和CUL4基因已确定通过控制莱茵衣藻中PSBS和LHCSR基因的表达响应强光、蓝光和紫外光,从而使NPQ快速恢复[106]。通过正确表达关键的调节NPQ基因提高生物质生产力具有发展潜力。
改善微藻中光合作用效率的另一种策略是引入具有更宽吸收光谱的异源光系统。光合有效辐射(PAR)是指波长为400~700 nm的太阳光部分,可用于启动光合作用。植物和微藻光系统仅能吸收PAR范围的可见光,而约50%的可用太阳辐射落在该范围外[107]。引入非天然色素的生物合成途径可能会增加吸收范围和光合效率。KOH等[108]从衣藻中克隆了叶绿素a加氧酶(CrCAO)基因到微拟球藻Nannochloropsis(只有叶绿素a,缺乏叶绿素b)中,以产生叶绿素b。12 d后,在中光照条件下,该藻细胞数增加26%,细胞干重提高31%,总脂质含量增加8%。此外,研究报道将叶绿素f合酶(ChlF)编码基因引入蓝藻,可使该生物体合成叶绿素f(Chl f),该色素在太阳光谱的远红色区域吸收,扩展了PAR的光谱范围[83]。ChlF可替代D1亚基,形成产生Chl f的改良型PSII复合体[109]。通过在蓝藻Synechocystissp. PCC 6803(其本身不含Chl f色素)中设计嵌合的D1/ChlF蛋白,该工程菌株可合成Chl f色素[109]。此外,基因工程还可用于指导细胞叶绿素或类胡萝卜素向其他色素的酶促转化,以扩展PAR谱[110-111]。
2.3.3 CO2同化作用的增强
研究表明,CO2浓度对微藻碳固定和生物质生产性能的影响复杂。大多数微藻仅在低浓度CO2水平下生长,CO2体积分数超过5%认为对微藻生长有毒,因此亟需在高浓度CO2(20%)下快速生长的微藻[27,112-113]。改善微藻CO2固定效率需提供足够的ATP并重新定向碳通量以生成代谢物。研究假设增加NADPH消耗途径可能会改善光反应和暗反应的耦合,从而提高光合效率和生物质产量[79]。ZHOU等[114]为验证这一假设,在蓝藻Synechocystissp. PCC 6803中构建了NADPH消耗途径。结果表明,NADPH消耗突变体的生物质产量和CO2固定率分别增加了84%和38%。HU等[115]开发了一种组合策略提高微生物固定CO2的效率,将产生ATP的羧化反应与消耗ATP的碳固定途径中的Rubisco分流器结合。该策略提供了足够的ATP以提高CO2固定效率,同时将CO2固定途径重新连接到生物合成的中心代谢途径。自养长聚球藻的CO2固定率和苹果酸产量分别提高110%和260 μmol/L。此外,一种名为“malyl-CoA-glycerate (MCG)”的人工合成途径被基因工程改造到聚球藻中延长PCC7942,使CO2同化加倍[79]。此外,通过使用更有效的羧化酶替代Rubisco或用其他碳固定途径替代卡尔文循环来调节光呼吸途径可提高微藻的光合效率[116-117]。上述结果表明碳固定途径可被设计,且新途径可显著增强碳固定。未来需进一步研究提高光合效率的方法。
2.4 纳米材料的干预
研究表明,纳米材料(NMs)在改善细胞行为方面潜力巨大[118]。NMs被定义为任一外部维度、内部或表面结构处于1~100 nm纳米尺度的材料。工业上常用的NMs有碳基纳米材料(CNMs)、金属氧化物基纳米材料(MONMs)和贵金属基纳米材料(NMNMs)。这些NMs通常与微藻存在不同程度的相互作用[118]。低浓度NMs在提高微藻生物量、脂质含量、改善光合作用等方面应用已有报道[119]。由于低浓度NMs可通过诱导分泌细胞外聚合物(EPS)和增强细胞壁提高对NMs抵抗力[120]。故在微藻培养过程中加入NMs有望提高微藻产量和经济效益。
使用纳米纤维作为物理吸附剂的CO2生物固定研究前景广阔,培养结束时易将微藻生物质与纳米结构分离[123]。VAZ等[124]开发了含有氧化铁纳米颗粒(NPsFe2O3)的聚合物纳米纤维,用于培养小球藻LEB111。NPsFe2O3增加了纳米纤维的比表面积和孔隙率,这些特性对于纳米结构作为吸附材料的应用至关重要。含有纳米纤维的微藻中表现出更高的CO2固定率(216.2 mg/(L·d)),以及更高的生物量和碳水化合物产量,证实了电纺纳米作为CO2物理吸附剂的潜力。VAZ等[124]研究表明,吸附剂纳米纤维增加了小球藻LEB111在室外试验中的CO2生物固定及脂质产量[125]。此外COMITRE等[37]探究了通过更新小球藻LEB111培养物中聚合物纳米纤维来提高微藻固定CO2能力,发现培养的第7天和第15天,纳米纤维的双重更新提高了小球藻的生物质浓度和CO2的生物固定率。
此外,碳基NMs可通过增加光吸收光谱促进微藻生长和光合作用。碳点(CDs)毒性低、可生物降解,ZHANG等[126]采用小球藻作为模型微藻研究CDs的影响,使生物量和生长率分别提高17%和21%。这是由于CDs可被小球藻降解并产生大量CO2,这些CO2被叶绿体固定并增强光合作用。CDs可将Rubisco酶体内活性提高34%。此外,CDs可吸收紫外光并作为抗氧化剂保护小球藻抵抗紫外线辐射的伤害。NGUYEN等[127]研究发现含氨化镁(MgAC)培养基中添加壳聚糖基碳点N(CD)s,有助于绿藻Tetraselmissp.抵消紫外线辐射的不利影响,并将光辐射转化为有利于光合作用的波长,提高了绿藻的生长速率和生物量生产力。氧化石墨烯量子点(GOQDs)被认为可促进PSII能量转移从而增强微藻的光合作用。YANG等[128]将在紫外(380 nm)激发后具有蓝光(465 nm)辐射的氧化石墨烯量子点(GOQDs)引入微藻培养基中,系统中发现CO2固定率提高了20%,脂质积累增加了34%。证明使用GOQDs提高微藻的光合效率潜力巨大,尤其在太阳紫外线较高的环境中,但仍有必要探索更有效的NMs来增强吸收光谱。

在微藻培养中添加NMs具有突出优势,如增强CO2在溶液中扩散/溶解、光吸收光谱和色素含量,但由于添加NMs会对微藻细胞造成氧化应激,增加ROS水平,研究表明高浓度NMs会抑制光合作用,故在低浓度时可改善微藻生长和光合效率[118]。
3 发展前景
目前,随机诱变、适应性实验室进化、基因工程和纳米材料干预对增强微藻固碳潜力巨大。但由于微藻的固碳过程相对复杂,仍存在局限性,需进一步深入探索。
1)工业烟气的主要成分为CO2,同时包含NOx、SOx、PM2.5、重金属Hg等多种残留物。通过逐渐增加CO2浓度筛选耐受更高浓度CO2的微藻菌株是实现有效、经济可行的CO2生物固定的关键。而微藻种类繁多,藻类研究仅关注少数种类,需进一步关注其他潜在微藻。此外,微藻-CO2固定过程中应进行更完整的经济分析和敏感性分析,以获得各产品生物质加工、生命周期、实时场景中的工业应用、市场增值终端产品等。
2)随机诱变被认为是一种有前景的突变育种技术,广泛应用于菌株改良和代谢过程的分子基础研究。化学诱变易操作,但化学试剂有害。与化学方法相比,紫外诱变控制更灵活,可减少二次污染,但毒性较大、突变率低、辐射风险大等,需进一步解决。
3)尽管在微藻培养中添加NMs具有诸多优势,如增强CO2扩散/溶解、增强微藻光吸收光谱及可回收和重复使用NMs等,但仅在低浓度NMs下,微藻的生长和光合性能得以改善,当微藻暴露在高浓度NMs中时可能会限制其光合作用。
4)目前在微藻中掺入NMs的相关研究仅在实验室规模下进行,NMs昂贵的价格和毒性限制了其在工业中的应用。在微藻培养后回收和重复利用NMs是一种降低成本的有效方法。如可从培养基中简单离心分离产生的微藻生物质实现回收。
5)利用基因编辑技术对微藻进行基因修饰比较繁琐,需要复杂的设备和试验条件,价格昂贵,实验周期长。但与其他修饰相比,具有以下优势:可定向操作,根据需求设计基因,打破物种之间的界限。未来有必要确定更多微藻基因组序列,阐明基因型和表型之间的内在关系。通过人工智能算法进行遗传设计,加速微藻基因工程的发展和应用。
4 结 语
可再生能源和碳捕集与利用技术是满足世界能源需求和减少温室气体排放的2个重要解决方案。微藻生物质作为可再生能源,在众多碳利用技术中尤为重要:① 以高光合速率更有效固定温室气体CO2,具有减少烟气中CO2排放的潜力;② 实现碳利用,可转化为高附加值产品(如生物燃料、保健品、化妆品、动物饲料等),实现经济效益。微藻适应恶劣环境、耐受高浓度CO2,并具备生长率和光合效率,是研发成功微藻固碳系统的关键。系统讨论了微藻的光合作用和相关碳固定机制。小球藻、螺旋藻、栅藻、葡萄藻、杜氏藻等微藻是大规模户外种植以减少发电厂烟气中CO2排放的竞争候选者。可使用随机诱变、适应性实验室进化、基因工程和纳米材料的干预等策略来提高微藻的固碳效率。其中,基因工程作为一种新颖而强大的基因技术应用潜力巨大,不仅可截断LHC天线尺寸来提高光合效率,还可提高Rubisco的速度和选择性。此外,纳米材料可增强CO2在溶液中扩散/溶解,显著提高PSII中的相对电子传输速率以及微藻中ROS水平,从而改善对类胡萝卜素的一般光合作用。
猜你喜欢
中国岩溶(2022年3期)2022-11-30
南方农业(2022年12期)2022-11-23
西南农业学报(2022年8期)2022-10-28
科学技术创新(2022年28期)2022-10-21
亚热带农业研究(2022年1期)2022-08-08
海洋湖沼通报(2022年2期)2022-05-08
古今农业(2022年1期)2022-05-05
知识就是力量(2021年6期)2021-07-09
农业科技通讯(2021年1期)2021-03-06
中国农业科技导报(2020年3期)2020-03-15
