
水稻香味基因Badh2的功能和效应分析
2023-12-02 12:10刘之熙闵军刘三雄刘利成胡敏李咏谊
湖南农业科学 2023年10期
刘之熙 闵军 刘三雄 刘利成 胡敏 李咏谊

摘 要:香稻是一种具有芳香气味的特种稻资源,具有较高的营养价值和经济价值。通过克隆香味基因Badh2启动子区插入序列badh2-p,构建启动子区插入突变表达载体pCAMBIA 1300-badh2-pi,转化水稻品种C5。采用qRT-PCR分析Badh2基因的表达量,同时利用GC-IMS联用技术测定香味物质2-AP和其他挥发性有机物的含量。结果表明:转基因株系叶片中Badh2基因的表达水平都升高了,其中有2株的Badh2基因表达水平极显著升高;转基因株系叶片中大部分挥发性化合物的含量高于野生型C5,2-AP浓度与Badh2基因的相对表达量水平成正相关。
关键词:水稻;Badh2基因;启动子区插入突变;香味;分子机制
中图分类号:S511文献标识码:A文章编号:1006-060X(2023)10-0001-06
Function and effect analysis of Badh2 Gene in rice
LIU Zhi-xi, MIN Jun, LIU San-xiong, LIU Li-cheng, HU Min, LI Yong-yi
(Hunan Rice Research Institute, Key Laboratory of Indica Rice Genetics and Breeding in the Middle and Lower Reaches of Yangtze River Valley, Ministry of Agriculture, Changsha 410125, PRC)
Abstract:Fragrant rice is a kind of special rice resource with aromatic smell, which has high nutritional and economic values. By establishing promoter insertion mutation of Badh2 gene, analyzing Badh2 gene expression, and determining content difference of 2-AP and other volatile organic compound, the relationship between promoter insertion mutation of Badh2 gene and aroma formation, and the molecular mechanism of rice aroma formation were explored. The results showed that the expression levels of Badh2 gene in the leaves of transgenic strains increased, with two of them showing extremely significant increases; the content of most volatile compounds in the leaves of transgenic strains was higher than that of wild-type C5, and the concentration of 2-AP was positively correlated with the relative expression level of Badh2 gene.
Key words:rice; Badh2 gene; promoter insertion mutation; fragrance; molecular mechanism
收稿日期:2022-12-20
基金項目:湖南省自然科学基金青年基金项目(2019JJ50334);湖南省农业科学院科技创新资金项目(2018QN06)
作者简介:刘之熙(1981—),女,山西长治市人,副研究员,主要从事水稻分子育种和种子质量检测技术研究。
水稻(Oryza sativa L.)是世界上最重要的粮食作物之一,全世界有近 30 亿人口以水稻为主食[1]。香米不仅在蒸煮后清香可口,更具有很高的营养价值。同时香米在国际市场上广受欢迎,价格比非香大米高2倍以上,这也为香稻研究提供了广阔的市场前景。
香稻含有多种挥发性化合物,有学者发现直接与水稻香味相关的挥发性物质为2-乙酰1-吡咯啉(2-acetyl-1-pyrroline,2-A'P)[2-3]。由于甜菜碱醛脱氢酶基因Badh2功能的缺失,翻译提前终止而产生无功能的BADH2蛋白,中断了2-AP合成底物γ-氨基丁醛的代谢,使其转而生成了香味物质2-AP[4]。Badh2基因位于8号染色体上[5],由 15 个外显子和 14 个内含子组成,常见的突变类型为第7外显子8 bp缺失和3个SNP差异导致的移码突变,以及第2外显子上7 bp缺失造成的翻译提前终止[6-7]。随着香味基因的克隆和香稻资源的收集,共发现有20余种badh2的等位基因[8-11]。除了编码区 SNPs 或 InDels 引起非同义突变或移码框突变而导致无功能的BADH2蛋白产生的变异类型外,还有一些Badh2基因的变异发生在内含子区、启动子区及5'UTR区,携带这些变异类型的水稻品种大多散发独特的香气。
笔者前期利用二代测序技术对3个香稻品种XW13、YZX及XW17进行全基因组测序,发现3个香稻品种的Badh2基因序列一致,在编码区无明显变异的情况下,启动子区距离ATG -1487 bp处有一个8 bp的插入突变,这种变异类型在已测序并公开发表的76份香稻资源的Badh2基因序列中也有发现,命名为 badh2-p[12]。但该变异类型如何影响Badh2基因的表达,从而使水稻产生香味的遗传机制还未见报道。研究通过克隆badh2-p,构建启动子区插入序列与非香品种编码区序列的双元表达载体转化水稻品种C5,分析Badh2基因的表达差异并测定香味物质2-AP和其它挥发性有机物成分的差异,分析该位点变异对香味形成的遗传效应。
1 材料与方法
1.1 试验材料
转基因受体C5由湖南省水稻研究所水稻轻简安全高效遗传改良创新团队提供。经测序验证,C5中Badh2基因的变异类型为第7外显子存在8 bp缺失,即badh2-E7。
1.2 badh2-p的克隆及载体的构建
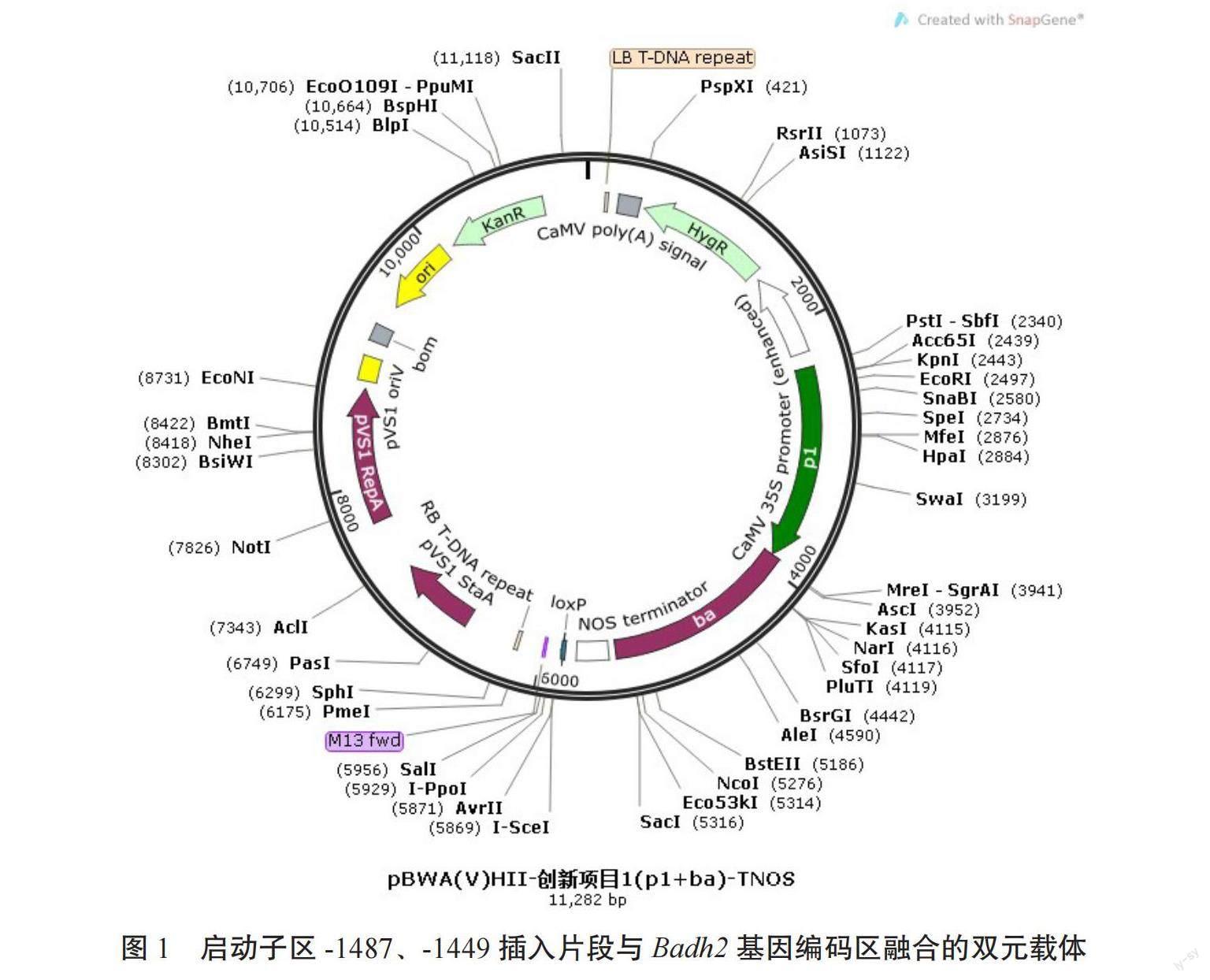
试验采用CTAB法提取香稻品种XW13叶片DNA,并以其为参考序列设计Badh2基因启动子区和编码区扩增引物,p1(+):5'- CGATGGTCTCACAACGAAGGCTATTTTAATTTTATT ATTGTTAGATTTTTTTTGCCTAA-3';p1(-):5'- CGATGGTCTCATGGTGGAGTGGAGCGTTGAGG TGGAAAGCGATAGGTATCTTTCTATCT-3',ba(+):5'-CGATGGTCTCAACCAATGGCCACGGCGATCCCGCAGCGGCAGCTCTTCGTCGCCGGCGA-3';ba (-):5-'CAGTGGTCTCATACATTACAGCTTGGAAGGGGATTTGTACCATCCCCACGGCTCATCGG-3'。扩增片段通过同源重组连接到植物表达载体pCAMBIA 1300上,构建启动子区插入突变植物表达载体pCAMBIA 1300-badh2-pi。
1.3 T1代转基因植株鉴定
将经过验证的表达载体转化到农杆菌感受态细胞EHA105中,浸染并转化C5愈伤组织,通过潮霉素抗性基因特异引物筛选T0代转基因阳性植株。取阳性株系和野生型各20粒种子种植,在苗期采用CTAB法提取基因组DNA,利用潮霉素抗性基因特异引物对T1代转基因植株进行筛选。hyg(280)-F: 5'-ACGGTGTCGTCCATCACAGTTTGCC-3';hyg(280)-R:5'-TTCCGGAAGTGCTTGACATTGGGGA-3'。PCR反应体系(10 ?L)为:2×Taq Master Mix 5 ?L、 正反向引物各0.2 ?L、DNA 1 ?L和H2O 3.6 ?L。PCR反应条件为:94℃ 4 min; 94℃ 15 s,55℃ 15 s,72℃ 30 s, 35个循环;72℃ 5 min。PCR产物在4%的琼脂糖凝胶上进行电泳分离,Gelred核酸染料染色后在紫外灯下拍照。
1.4 Badh2基因的表达量分析
利用RNA-easy Isolation Reagent提取试剂盒分别提取T1代转基因与野生型植株齐穗期叶片的总RNA,采用HiScript III All-in-one RT SuperMix Perfect for qPCR合成试剂盒反转录合成第1链cDNA,然后利用荧光定量 PCR分析 Badh2基因在转基因和野生型植株中的表达量。以Actin 基因作为内参基因。qRT-PCR 反应体系如下:cDNA 模板 3 μL,2×SYBR qPCR Mix (TOYOBO) 10 μL,正反引物(10 μmol/L)各 1 μL,dd H2O 补足至 20 μL。PCR 扩增程序如下:95℃下预变性 3 min,95 ℃ 10 s,60 ℃ 30 s,72 ℃ 20 s,45 个循环。利用公式 2-ΔΔCT方法计算基因的相对表达量。
1.5 挥发性化合物分析测定
根据qRT-PCR结果,在齐穗期选取T1代转基因植株中Badh2基因表达量极显著上升的单株和野生型单株,采用GC-IMS联用技术测定其中挥发性有机物成分的含量。各剪取0.5 g叶片置于20 mL顶空瓶中,60℃孵育15 min,密封后備用,每个样品做3 个平行。
GC-IMS测定条件:采用自动顶空进样;顶空孵化温度60 ℃;孵化时间15 min;孵化转速500 r/min;进样针温度85 ℃;进样体积500 μL,不分流模式;色谱柱MXT-5(15 m×0.53 mm,1 μm);柱温60 ℃;分析时间20 min;载气为高纯N2(纯度≥99.99%);柱流速初始为2 mL/min,保持2 min,2~10 min柱流速线性增加至10 mL/min,10~20 min柱流速线性增加至100 mL/min。
1.6 数据处理
采用仪器自带分析软件包括LAV(Laboratory Analytical Viewer)和3款插件(Reporter插件、Gallery Plot插件、Dynamic PCA插件)以及GC×IMS Library Search进行挥发性化合物分析;采用SPSS 25.0进行显著性分析,显著性差异为P<0.05,Origin 2020软件绘图。
2 结果与分析
2.1 植物表达载体的构建及遗传转化
利用CTAB法提取香稻品种XW13叶片DNA,以其为参考序列设计Badh2基因启动子区和编码区扩增引物,扩增片段通过同源重组连接到植物表达载体pCAMBIA 1300上,构建启动子区插入突变植物表达载体pCAMBIA 1300-badh2-pi(图1)。利用农杆菌介导法转化C5愈伤组织,通过潮霉素抗性基因特异引物筛选T0代转基因阳性植株,获得转基因T0代阳性植株31株(图2)。
2.2 T1代转基因株系中香味基因Badh2 相对表达量的检测
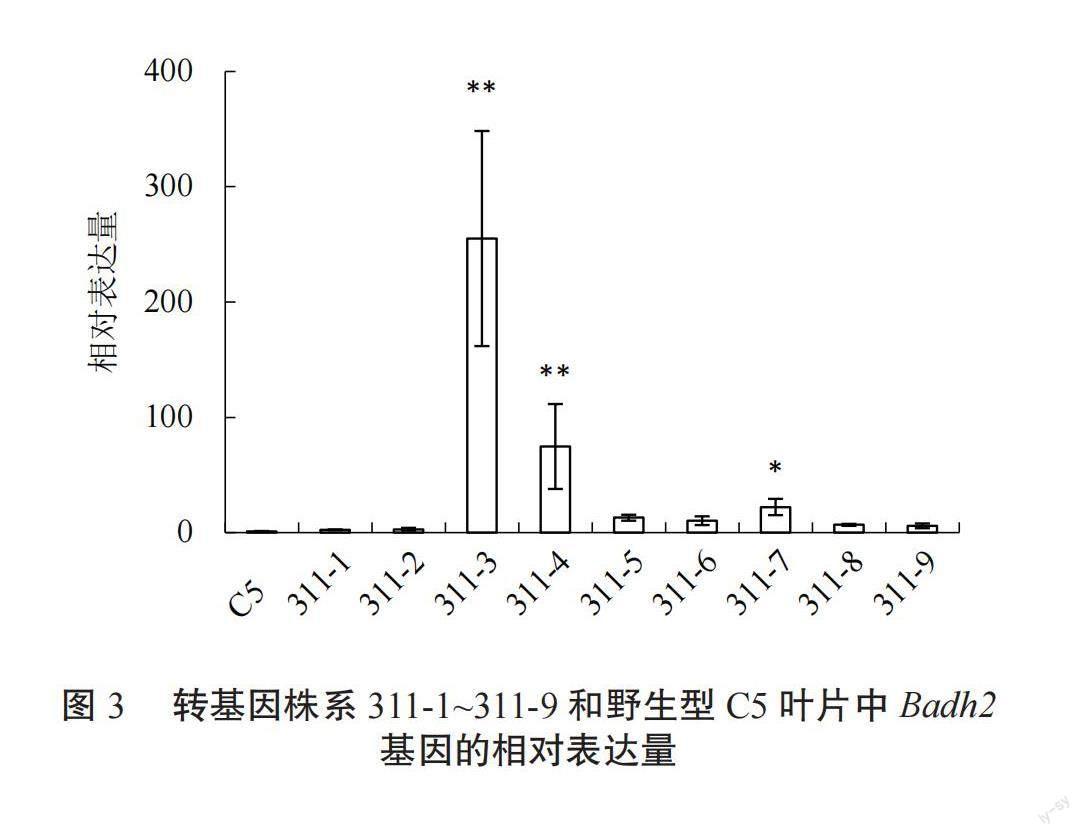
T0代转基因阳性植株自交获得T1代,在苗期利用潮霉素抗性基因特异引物对T1代转基因植株进行筛选。将阳性单株移栽,齐穗期分别提取T1代转基因与野生型植株叶片的总RNA,利用荧光定量 PCR进行Badh2基因总mRNA表达分析。结果显示,31株转基因株系叶片中Badh2基因表达水平均有升高,其中2株转基因株系Badh2基因表达水平极显著升高,分别为:311-3,311-4;4株转基因株系Badh2基因表达水平显著升高,分别为:311-7,311-18,311-26,311-29(图3~图5)。
2.3 轉基因株系挥发性化合物GC-IMS分析
选取Badh2基因表达量极显著上升的单株311-3、311-4,及野生型C5单株齐穗期的叶片,采用GC-IMS仪器LAV分析软件中的Reporter插件程序对不同转基因株系和野生型叶片中挥发性化合物进行三维对比,结果见图6。图6中每一个峰都代表
一种挥发性化合物,以点的颜色和面积表示含量高低,点的颜色越深、面积越大表示该物质含量越高,红色点表示物质含量较高,而白色点表示含量较低。从色彩角度观察3D谱图,不同转基因株系较为相似但又存在不同,很难直观区分。
为了便于比较不同转基因株系叶片中挥发性化合物的差异,以野生型C5指纹图谱作为参比,其他转基因株系图谱扣除参比,若挥发性化合物含量相同则扣除背景后为白色,高于参比为红色,低于参比为蓝色,如图7所示。不同转基因株系的挥发性化合物含量存在差异,311-3样品蓝色部分和红色部分较311-4样品明显较多,说明311-3和311-4样品中各有一部分挥发性物质含量高于野生型C5,一部分低于野生型C5,且311-3和311-4样品的挥发性物质含量存在明显差异。
2.4 转基因株系叶片中挥发性化合物的指纹图谱分析
为了更直观反映转基因株系叶片中挥发性化合物的变化规律及相对含量差异,每个样品重复3 次测定,采用Gallery Plot插件绘制挥发性化合物指纹谱图。图中每行代表一个样品的挥发性化合物,颜色深浅代表挥发性化合物的含量高低。由图8可知,不同转基因株系的挥发性化合物分布不同,各自有特征峰区域同时也存在共同区域。野生型C5和转基因株系之间的差异显著,红框所示物质在各自样本中含量相对较高。其中,香稻典型性香气成分—2-AP(爆玉米花、茉莉花香)在311-3中浓度最高,在311-4中浓度次之,在C5中浓度最低,这与Badh2基因在3个单株中的相对表达量水平成正相关。311-3中异戊醛(具有苹果香气)、1-戊烯-3-醇(水果香味)、反-2-庚烯醛(具青草香气)、庚醛(水果香味)、2-戊基呋喃(具豆香、果香等香气)、2-庚酮(梨香)等含量较高;311-4中2-甲基丁醛(咖啡、可可香气)、2-丁酮(甜醋、似橙皮香气)、丁醛、乙偶姻(呈奶油香气)、2-乙基呋喃(甜香和咖啡似香气)、2-乙酰基呋喃(有杏仁、坚果、焦糖似香气)、苯甲醛(苦杏仁气味)、异丁酸异丁酯(呈菠萝、葡萄皮香气)、水杨酸甲酯(具有冬青叶香味)等含量较高;C5中反-2-辛烯醛(呈脂肪和肉类香气)、反-2-戊烯醛(呈土豆和豌豆似香气)、正戊醇(略有奶油香气)、顺-2-戊烯醇(绿茶陈化风味)等挥发性化合物含量最高。2,4-庚二烯醛(水果香味)、丙酮(似薄荷香气)、2-己烯醇等挥发性物质在不同株系中含量接近。由图8可知,转基因株系大部分挥发性化合物含量高于野生型C5,且311-3、311-4中部分挥发性物质含量增加较为明显。
2.5 转基因株系叶片挥发性化合物的主成分分析
主成分分析是多元统计分析中一种无标签的数据降维方法,并能最大限度保留样本原始信息。采用Dynamic PCA插件对转基因株系和野生型C5挥发性化合物进行主成分分析,可以更直观地判别区分并可视化不同株系挥发性化合物的差异。由图9可知,野生型C5和2个转基因株系均能够较好地分离。其中,野生型C5和311-3之间的距离较远,表明两者之间挥发性化合物存在较大差异。而野生型C5和311-4距离较近,表明二者之间的挥发性化合物差异较小。主成分结果表明,GC-IMS分析结果能够较好地判别和区分不同的株系。
3 讨 论
前人对控制香味性状的甜菜碱醛脱氢酶基因(Badh2)的研究主要集中在编码区和内含子区碱基的插入、缺失或突变,对启动子区的变异研究较少。本文通过建立启动子区插入片段突变转化体系,分析其与野生型Badh2基因的表达水平、香味物质2-AP的含量差异及其它挥发性有机物成分的差异,发现该启动子区插入突变会引起Badh2基因RNA水平显著上升,2-AP含量和大部分挥发性化合物含量也明显提高。究其原因,可能是因为转基因株系中2-AP代谢通路上某些酶(鸟氨酸转氨酶、吡咯啉-5-羧酸合成酶)含量增加,导致谷氨酸-脯氨酸相互转化过程中间物1-吡咯啉-5-羧酸的积累,能够生成1-吡咯啉,最终与乙酰基团结合生成2-AP,从而导致2-AP含量和大部分挥发性化合物含量出现了明显的上升。具体的调控机理与分子机制,目前正在深入的分析研究中。
参考文献:
[1] HU C Y, SHI J X,QUAN S, et a1. Metabolic variation between japonica and indica rice cultivars as revealed by non-targeted metabolomics[J]. Scientific Reports,2014,4(5067):1-10.
[2] BUTTERY R G, LING L C, JULIANO B O,et al. Cooked rice aroma and 2-acetyl-1-pyrroline [J]. Journal of Agricultural and Food Chemistry,1983,31(4):823-826.
[3] TSUZUKI E, SHIMOKAWA E. Inheritance of aroma in rice[J]. Euphytica,1990,46(2):157-159.
[4] CHEN S H,YANG Y,SHI W W,et a1. Badh2,encoding betaine aldehyde dehydrogenase,inhibits the biosynthesis of 2-acetyl-1-pyrroline a major component in rice fragrance[J]. The Plant Cell,2008,20(7):1850-1861.
[5] AHN S N,BOLLICH C N,TANKSLEY S D. RFLP tagging of a gene for aroma in rice[J]. Theoretical and Applied Genetics,1992,84(7/8):825-828.
[6] BRADBURY L M T,FITZGERALD T L,HENRY R J,et a1. The gene for fragrance in rice[J]. Plant Biotechnology Journal,2005,3(3):363-370.
[7] SHI W W, YANG Y, CHEN S H, et al. Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties[J]. Molecular Breeding, 2008, 22(2): 185-192.
[8] GOLESTAN HASHIEMI F S,RAFII M Y,ISMAIL M R,et al. Biochemical,genetic and molecular advances of fragrance characteristics in rice[J]. Critical Reviews in Plant Sciences,2013,32(6):445-457.
[9] ARAWATHI Y,SINGH R,SINGH A K,et al. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.)[J]. Molecular Breeding,2008,21:49-65.
[10] KOVACH M J, CALINGACION M N, FITZGERALD M A, et al. The origin and evolution of fragrance in rice (Oryza sativa L.)[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(34): 14444-14449.
[11] SHAO G N,TANG S Q,CHEN M L,et a1. Haplotype variation at Badh2,the gene determining fragrance in rice[J]. Genomics,2013,101(2):157-162.
[12] 劉之熙,朱克永,闵 军,等. 基于基因组测序揭示香稻Badh2基因的变异信息[J]. 湖南农业科学,2019(8):1-4.
(责任编辑:高国赋)
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27
军事文摘(2021年22期)2021-11-26
文苑(2020年6期)2020-06-22
文苑(2019年22期)2019-12-07
中国中药杂志(2016年21期)2017-02-16
云南中医中药杂志(2016年9期)2016-11-29
儿童故事画报·智力大王(2015年12期)2016-01-23
中国钱币(2015年6期)2015-11-18
现代仪器与医疗(2015年3期)2015-07-06
幼儿时代·故事妈妈(2004年3期)2004-03-17
