
应用转录组分析不同施肥对玉米穗位叶中糖类、脂类和蛋白质代谢的影响
2023-12-29 01:37朱凯丽刘立功刘树堂
西南农业学报 2023年10期
曾 黎,朱凯丽,刘立功,刘树堂,李 军
(1. 青岛农业大学农学院, 山东 青岛 266109;2. 北京市农林科学院蔬菜研究所,北京 100097;3. 青岛农业大学资源与环境学院, 山东 青岛 266109)
【研究意义】氮(N)、磷(P)和钾(K)是植物生长发育必需的大量元素,N和K分别占植物干重的2%和10%,而活细胞中P浓度约在毫摩尔范围内[1-3]。生产上N、P和K肥的合理配施不仅有利于植物的生长发育还有助于作物增产[4-7]。因此,从分子水平上解析NPK配施对三大营养元素间的互作效应以及糖类、脂类和蛋白质代谢的影响,对揭示肥料高效利用和作物品质改良的调控机制具有重要意义。【前人研究进展】南镇武等[8]、段有强[9]和周录英等[10]研究发现,土壤中施加氮肥有利于提高冬小麦和花生籽粒蛋白质含量;Almodares等[11]和Shen等[12]研究发现,施加钾肥增加了高粱和梨果实的可溶性糖含量。周录英等[10]研究发现,磷肥对提高籽仁蛋白质和脂肪含量效果明显,且合理的氮磷钾配施可增加花生籽仁的赖氨酸、蛋氨酸和油酸、亚油酸含量,提高油酸/亚油酸比值,从而改善花生营养品质;Oloyede等[13]报道,少施或不施NPK肥有利于提高南瓜果实中抗氧化酚类物质的积累。这些研究表明,除增加作物产量外,NPK肥配施还对作物的营养品质产生较大影响。近年来,RNA-Seq技术已发展成为分析植物基因表达谱的强有力工具。许多关于N、P或K营养匮乏的转录组分析已在玉米[14-17]、水稻[18-19]和高粱[20]中进行研究,发现了许多参与N、P或K吸收和代谢相关的差异表达基因。然而,这些研究都是在缺乏某种特定营养元素且大多数是在水培条件而非在田间条件下进行的。Xu等[21]认为这种情形产生的原因在于建设长期定位施肥试验点存在一定的困难。聂胜委等[22]认为,长期定位施肥试验比常规田间试验在评价不同施肥制度对作物生长发育和产量等因子效应方面数据更加可靠、解释力更强。【本研究切入点】目前有关田间条件下,施加NPK肥对玉米叶片糖类、脂类和蛋白质代谢相关基因的表达模式与分子调控机制报道较少。【拟解决的关键问题】本研究利用高通量RNA-Seq测序结果,解析N、K和P肥配施条件下糖类、脂类和蛋白质代谢相关基因的表达模式,揭示不同肥料配施对三大有机物质代谢相关基因表达的分子调控机制,为提高玉米叶片光合利用率提供依据。
1 材料与方法
1.1 试验材料
夏玉米品种为“鲁豫16号”,种植于青岛农业大学(原莱阳农学院)莱阳长期定位试验站。该站建于1978年,用于评价长期施肥效率。有关试验站的详细描述参见Song等[23]。
本试验设置5个处理:不施肥(CK)、氮肥(N)、氮+磷肥(NP)、氮+钾肥(NK)和氮+磷+钾肥(NPK)(表1)。3次重复,随机排列,共15个小区,每个小区面积为33 m2。于灌浆期选取3片穗位叶/小区的上半部位,液氮冷冻,-80 ℃保存。

表1 试验田每年的施肥量Table 1 Fertilization rates in the experiment field per year (kg/hm2)
1.2 RNA提取、cDNA文库构建和Illumina测序
叶片总RNA提取和转录组测序工作委托武汉博越致和生物科技有限公司完成。总RNA提取后,分别用Qubit®RNA分析试剂盒(Life technologies, USA)和Labchip GXII Touch HT核酸分析仪(PerkinElmer, USA)测量RNA浓度和质量。质检合格后,用KAPATM单链RNA文库制备试剂盒(Illumina, USA)构建测序文库。然后在Illumina HiseqX平台上进行测序,获得原始数据(Raw data)。
1.3 RNA-Seq加工和基因功能注释
利用FastQC v0.11.5和Trimmomatic v0.36软件将原始数据进行过滤,获得高质量的序列数据(Clean reads)。以玉米B73基因组(gcf_00000 5005.2 _b73_refgen_v4_genome.fa)为参考,用Hisat2软件对Clean reads进行序列比对,获取在参考基因组上的位置信息以及测序样本特有的序列特征信息。计算每千个碱基的转录每百万映射读取的fragments(FPKM值)。用DESeq2软件检测基因的表达水平,以P<0.05,|log2FC (Fold change)|>1为筛选标准,获得不同处理下的差异表达基因(DEGs)。参照玉米B73基因组数据库信息,用Gene Ontology数据库进行查找,从而得到基因的功能注释。
1.4 差异基因功能富集分析
基于GO Database数据库(http://geneontology.org)和KEGG Database数据库(http://www.genome.jp/kegg),利用R语言的clusterprofiler软件包对DEGs进行GO和KEGG富集分析。为控制假发现率(FDR),计算q值以做校正,以P≤0.05为阈值,满足此条件的GO和KEGG通路定义为在差异基因中显著富集的通路。
1.5 统计分析
用SPSS 18.0和Excel 2010对数据进行统计分析和作图。
2 结果与分析
2.1 不同施肥处理下差异表达基因的GO功能富集分析
利用GO数据库,发现参与糖类、脂类和蛋白质代谢的DEGs在BP、CC和MF三大部分均有富集。尽管富集的条目相同,但在不同组合处理下DEGs数目各异(图1)。在所有组合处理中,NPvsCK组合在BP、CC和MF富集的DEGs最多,而NvsCK富集的DEGs最少。在BP中,DEGs在多糖代谢过程、脂质生物合成规程和胞内氨基酸代谢过程均有较高富集;在CC中,DEGs主要富集在叶绿体部分;而在MF中,DEGs主要富集在作用于糖基键的水解酶活性和蛋白丝氨酸/苏氨酸激酶活性。
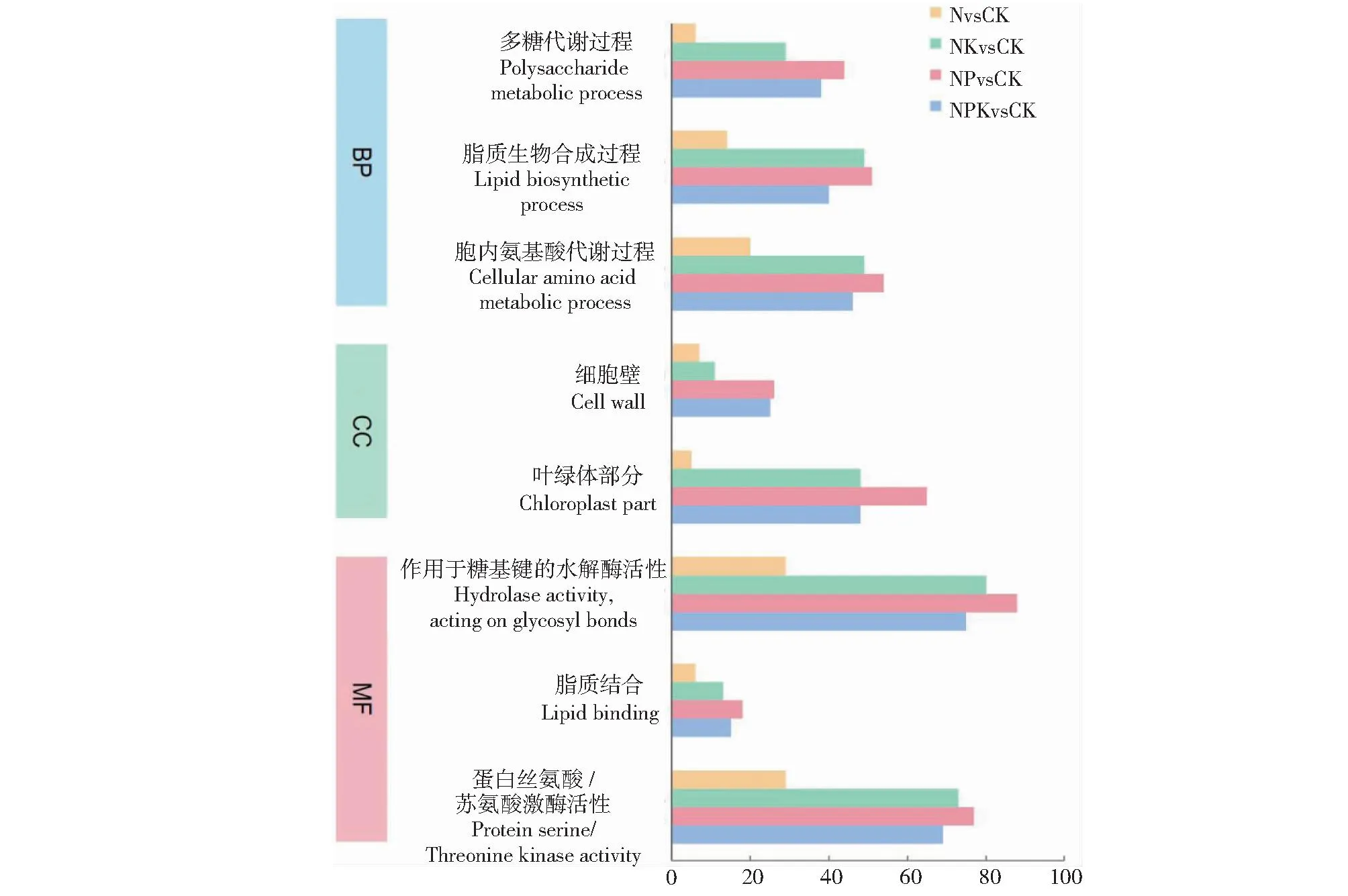
图1 差异表达基因GO富集分析Fig.1 GO enrichment analysis of differential expressed genes
2.2 不同施肥处理下差异表达基因的KEGG通路富集分析
由表2可以看出,4个处理组合中共同的显著性富集通路主要包括碳代谢、淀粉和蔗糖代谢、脂质代谢、氨基酸合成和丙氨酸、天门冬氨酸、谷氨酸代谢途径富集。在碳代谢、淀粉和蔗糖代谢和氨基酸合成途径中富集的DEGs数目较多。其中,NKvsCK、NPvsCK和NPKvsCK组合在这些代谢通路中富集的DEGs数目远远高于NvsCK组合。

表2 差异表达基因KEGG通路富集Table 2 KEGG enrichment of the differential expressed genes
2.3 不同施肥处理对玉米叶片糖类代谢相关DEGs的影响
以P<0.05和|log2FC|>1为筛选标准,发现在不同施肥处理下有82个DEGs参与玉米叶片糖类代谢的显著调控(图2)。除约12%的DEGs功能未知外,其余的DEGs分别参与蔗糖代谢、淀粉代谢、纤维素代谢、海藻糖代谢和糖分运输。其中,|log2FC|>2的DEGs有24个(表3),这些DEGs在NvsCK、NKvsCK、NPvsCK和NPKvsCK组合中的数量分别是6、17、20和14,以NPvsCK组合最多,NvsCK组合最少,说明氮和/或钾、磷肥配施诱导了更多基因参与玉米穗位叶糖代谢相关基因的表达。

图2 不同肥料处理下玉米叶片糖类代谢相关的DEGs数量(|log2FC|>1)Fig.2 The number of DEGs related to sugar metabolism in maize ear leaf under different comparisons of fertilizer treatments (|log2FC|>1)
AGPL2编码葡萄糖-1-磷酸腺苷转移酶大亚基2,参与淀粉的合成代谢,在所有处理组合中均显著上调。GRMZM5G803981和GRMZM2G025833编码产物是β-淀粉酶,参与淀粉的降解代谢,在NKvsCK、NPvsCK和NPKvsCK组合中均显著下调,但在NvsCK组合中却未见差异。GRMZM2G051806和GRMZM2G086845编码产物参与蔗糖的代谢,除在NvsCK组合中未见差异外,在NKvsCK、NPvsCK和NPKvsCK组合中均显著下调,其中在NPvsCK组合中下调幅度更大。GRMZM2G016890和GRMZM2G066162编码产物参与纤维素代谢,在NvsCK和NPvsCK组合中均显著上调,但在NKvsCK和NPKvsCK组合中却未见差异。GRMZM2G008226编码产物是α-海藻糖磷酸合成酶9,参与海藻糖代谢,在NvsCK、NKvsCK、NPvsCK和NPKvsCK组合中均显著上调。除TPS7在NvsCK组合中上调外,编码与海藻糖代谢相关的基因如GRMZM6G738249等均显著下调。发现约有33%的DEGs参与糖类物质的运输。其中,有3个基因显著上调,分别是在NvsCK、NKvsCK和NPvsCK组合中检测到的编码糖转运体的基因GRMZM2G063824,在NKvsCK、NPvs CK和NPKvsCK组合中检测到编码蔗糖转运蛋白BoSUT1的基因GRMZM2G081589,在NPKvsCK组合中检测到的编码糖转运蛋白sweet17-like的基因GRMZM2G106462。其余的编码与蔗糖转运蛋白相关的基因GRMZM2G077546、GRMZM5G839593和GRMZM6G056015在NKvsCK、NPvsCK和NPKvsCK组合中均显著下调,但在NvsCK组合中未见差异。
2.4 不同施肥处理对玉米叶片脂类代谢相关DEGs的影响
以P<0.05和|log2FC|>1为筛选标准,发现在不同施肥处理下有29个DEGs参与玉米叶片脂类代谢的显著调控(图3)。除约14%的DEGs功能未知外,其余的DEGs分别参与脂肪酸(含饱和、不饱和脂肪酸)生物合成、脂肪酸降解和脂肪酸延长。其中,|log2FC|>2的DEGs有6个(表4),这些DEGs在NvsCK、NKvsCK、NPvsCK和NPKvsCK组合中的数量分别是1、4、5和5,以NPvsCK和NPKvsCK组合最多,NvsCK组合最少。可见,单纯氮处理组合调控的DEGs数量明显少于其他处理组合。
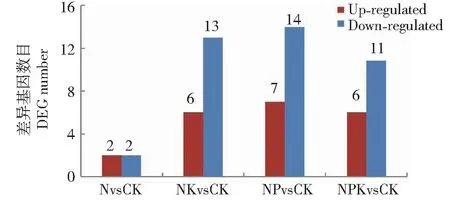
图3 不同施肥处理下玉米叶片脂肪酸代谢相关的DEGs数量(|log2FC|>1)Fig.3 The number of DEGs related to fatty acid metabolism in maize ear leaf under different comparisons of fertilizer treatments (|log2FC|>1)
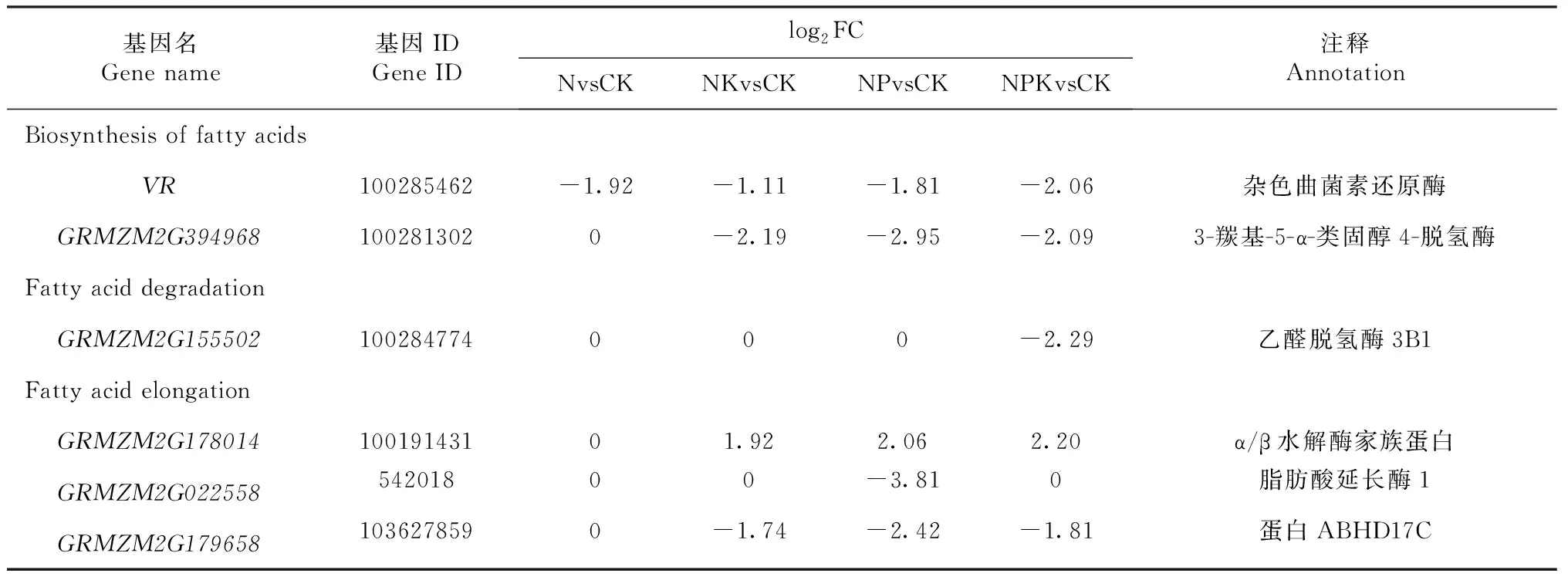
表4 不同处理组合中玉米叶片脂肪酸代谢相关的DEGs列表(|log2FC|>2)Table 4 DEGs related to fatty acid metabolism in maize ear leaf under various combinations of fertilizer treatments (|log2FC|>2)
VR编码产物是杂色曲菌素还原酶,参与脂肪酸的生物合成,在所有处理组合中均显著下调,以NPKvsCK组合下调幅度最大。参与编码类固醇代谢的3-羰基-5-α-类固醇4-脱氢酶和脂肪酸延长的蛋白ABHD17C的基因GRMZM2G394968和GRMZM2G179658,在NKvsCK、NPvsCK和NPKvsCK组合中均显著下调,但在NvsCK组合中却未见差异。GRMZM2G178014编码产物是α/β水解酶家族蛋白,参与脂肪酸的延长,在NKvsCK、NPvsCK和NPKvsCK组合中均显著上调,但在NvsCK组合中未见差异。
2.5 不同施肥处理对玉米叶片蛋白质氨基酸代谢相关DEGs的影响
以P<0.05和|log2FC|>1为筛选标准,发现在不同施肥处理下有84个DEGs参与蛋白质代谢的显著调控,其中有19个DEGs功能未知(图4)。其中,|log2FC|>2的DEGs有17个(表5),这些DEGs在NvsCK、NKvsCK、NPvsCK和NPKvsCK组合中的数量分别是9、14、13和14,以NKvsCK和NPKvsCK组合最多,NvsCK组合最少。可见,单纯氮处理组合调控的DEGs数量少于其他处理组合。

图4 不同施肥处理下玉米叶片氨基酸代谢相关的DEGs数量(|log2FC|>1)Fig.4 The number of DEGs related to amino acid metabolism in maize ear leafunderdifferent comparisons of fertilizer treatments (|log2FC|>1)

表5 不同处理组合中玉米叶片氨基酸代谢相关的DEGs (|log2FC|>2)Table 5 DEGs related to amino acid metabolism in maize ear leaf under various combinations of fertilizer treatments (|log2FC|>2)
与脯氨酸、天冬酰胺、组氨酸代谢相关的基因GRMZM2G061777、GRMZM2G028535、GRMZM2G078472和RPI2在所有处理组合中均显著上调,而与异亮氨酸、苏氨酸代谢相关的基因GRMZM2G153536、GRMZM2G147268、GRMZM5G871471显著下调。与丙氨酸、谷氨酸、色氨酸代谢相关的基因GRMZM2G124963、GRMZM2G375064和GRMZM2G138382在NKvsCK、NPvsCK和NPKvsCK组合中显著上调,但在NvsCK组合中未见差异。
3 讨 论
3.1 氮和/或钾、磷肥配施显著影响与糖代谢相关基因的转录水平
灌浆期玉米叶片旺盛的光合作用是籽粒糖分积累的主要来源[24]。转录组分析结果表明,灌浆期玉米穗位叶有82个参与糖代谢显著调控的DEGs,这与该时期叶片较高的光合效率相一致。基因功能注>释分析发现,这些DEGs富集主要在碳代谢、淀粉和糖代谢途径上,尽管富集的条目相同,但在不同肥料处理下DEGs数目差异显著,如NKvsCK、NPvsCK和NPKvsCK组合下的DEGs数量是NvsCK组合的2~3倍,说明氮和/或钾、磷肥配施显著影响了与糖代谢相关基因的转录水平,提高了光合产物由“源”到“库”的运输和积累,增加了作物产量[25]。
淀粉是植物光合作用主要的存贮物质。植物在白天合成淀粉但在夜间降解,这部分淀粉被称为“过渡性淀粉”[26],可为植物生长提供能量和碳源[27]。ADPG焦磷酸化酶(AGP)是淀粉合成前体-ADPG生成的限速酶,AGPL2是编码AGP大亚基的基因,在所有处理组合中均显著上调,表明施肥有利于玉米叶片淀粉的快速合成与积累。许多降解酶如淀粉磷酸酶、α-淀粉酶和β-淀粉酶,都参与淀粉的分解。已发现在拟南芥基因组中有9个编码β-淀粉酶(BAM)的基因,将淀粉水解成麦芽糖[28]。本研究结果表明,GRMZM5G803981和GRMZM2G025833基因在NKvsCK、NPvsCK和NPKvsCK组合中均显著下调,但在NvsCK组合中未见差异。说明施加钾和/或磷肥降低了灌浆期玉米叶片的淀粉水解,有利于淀粉积累。
海藻糖是一种非还原性双糖,由海藻糖-6-磷酸(T6P)经磷酸海藻糖合成酶(TPS)和磷酸海藻糖磷酸酶TPP催化合成[29],在低等植物中充当应激保护剂,但在高等植物中该作用被蔗糖所取代[30]。本研究发现,除GRMZM2G008226基因外,大部分参与海藻糖代谢基因表达下调,说明施加钾和/或磷肥明显抑制了海藻糖的生物合成。
在拟南芥中,AtSUCs和AtSWEETs基因编码蛋白负责蔗糖的转运[31]。本研究发现,在不同肥料处理下有21个DEGs参与了糖分转运,这与灌浆期同化产物的快速合成相吻合。在NKvsCK、NPvsCK和NPKvsCK组合中编码蔗糖转运蛋白BoSUT1的基因(GRMZM2G081589)和NPKvsCK组合中编码类糖转运蛋白sweet17的基因(GRMZM2G106462)显著上调;另一个编码碳水化合物转运蛋白的基因(GRMZM2G081589)在NvsCK、NKvsCK、NPvsCK处理组合中表达显著上调,但在NPK处理中表达差异不显著。说明不同肥料配施影响了糖物质的运输,而关于这些基因的功能尚待进一步研究。
3.2 氮和/或钾、磷肥配施显著影响与脂肪酸代谢相关基因的转录水平
农业生产上,施加无机肥料不仅能影响植物的糖代谢,而且还能影响植物的脂肪组成[10,12,32]。在脂肪酸生物合成过程中,乙酰辅酶A羧化酶(ACC)是负责将乙酰辅酶A催化生成丙二酰辅酶A的限速酶[33];脂肪酸合成酶(FAS)通过将丙二酸基转移到延伸链上,催化生成C16:00和C18:0-ACP;脂肪酸脱氢酶(FAD)催化产生不饱和脂肪酸;脂肪酸延长酶(FAE)催化产生长链脂肪酸。有研究表明,脂肪酸伸长酶1(FAE1)的突变改变了甘蓝型油菜的脂肪酸组成[34]。在本研究中,编码脂肪酸延长酶1的基因(GRMZM2G022558)在NPvsCK组合中显著下调,但在NvsCK、NKvsCK和NPKvsCK组合中却未见差异,说明脂肪酸链的延长在玉米穗叶中受到磷肥的高度调控。
编码3-羰基-5-α-类固醇4-脱氢酶1的基因(GRMZM2G394968)在NKvsCK、NPvsCK和NPKvsCK组合中显著下调,而编码α/β-水解酶家族蛋白的基因(GRMZM2G178014)则在这些处理组合中表达显著上调。表明氮和/或钾、磷肥配施显著影响了这些DEGs的表达水平。
3.3 氮和/或钾、磷肥配施显著影响与蛋白质氨基酸代谢相关基因的转录水平
在植物中,氨基酸是重要的氮存储化合物[35]。本研究发现,灌浆期玉米叶片中共有84个DEGs基因受到调控,表明该时期氨基酸的生物合成过程异常活跃。与其他处理组合相比,NvsCK组合的DEGs数量减少了2~3倍,说明氮和/或钾、磷肥配施显著提高了玉米叶片中氨基酸代谢基因的表达水平,进而对籽粒品质产生影响。谷氨酰胺合成(GOGAT)和谷氨酸合成酶(GS)在植物中将铵态氮同化为氨基酸过程中起着重要作用[36]。在本试验中,研究发现NKvsCK、NPvsCK和NPKvsCK组合中谷氨酸合成基因(GRMZM2G375064)显著上调,说明添加钾、磷肥促进氮肥由硝态氮向氨基酸的同化。已有研究表明,脯氨酸可以充当渗透保护剂,对干旱或盐等非生物逆境进行响应[37]。在本研究中,2个与脯氨酸生物合成相关的基因(GRMZM2G061777和GRMZM2G028535)在所有处理组合中均显著上调,这与GO富集分析结果一致,即在NKvsCK、NPvsCK和NPKvsCK组合中富集了许多参与非生物胁迫应答的基因。天冬酰胺在许多植物器官如萌发的种子、衰老器官等的氮循环、存储和运输中发挥着重要作用[38]。编码天冬酰胺合成酶4的GRMZM2G078472在所有组合处理中均显著上调,而编码天冬酰胺合成酶3的GRMZM2G053669仅在NvsCK和NPKvsCK组合中显著上调。酪氨酸、苯丙氨酸和色氨酸属于芳香族氨基酸(AAA),它们参与植物激素、色素和生物聚合物等众多第二代谢物的生物合成[39]。与色氨酸生物合成有关的基因(GRMZM2G138382)仅在NKvsCK、NPvsCK和NPKvsCK组合中显著上调,说明氮和/或钾、磷肥配施促进色氨酸的生物合成。
4 结 论
氮和/或钾、磷肥配施显著影响与玉米叶片糖类、脂类和蛋白质氨基酸代谢相关基因的转录水平,鉴定出一些参与三大有机物质代谢的关键基因,为未来通过控制肥料配施来提高玉米叶片光合利用率提供了分子依据。
猜你喜欢
军事文摘(2023年18期)2023-10-31
中国化肥信息(2022年2期)2022-04-19
作文周刊·小学四年级版(2022年8期)2022-03-11
中国化肥信息(2021年10期)2022-01-19
中国生殖健康(2020年5期)2021-01-18
中国化肥信息(2020年7期)2020-03-19
小学生必读(低年级版)(2019年5期)2019-08-30
中国生殖健康(2018年5期)2018-11-06
中国化肥信息(2018年2期)2018-08-23
食品与生活(2017年12期)2018-01-09
