
基于代谢组学探讨犊牛失明原因
2023-12-29 01:37王建东唐玉林郭亚男高海慧侯鹏霞郭延生
西南农业学报 2023年10期
王建东,唐玉林,郭亚男,高海慧,侯鹏霞,于 洋,郭延生
(1.宁夏农林科学院动物科学研究所,银川 750002;2.宁夏大学农学院,银川 750021)
【研究意义】宁夏六盘山一带的肉牛养殖属于传统产业,但同时存在一家一户的粗放养殖模式,饲料单一,不添加微量元素和维生素等添加剂,犊牛失明发病率在5%左右,一般不会引起死亡,如果产犊及时发现,通过即时注射维生素A会减轻发病率。尚青山[1]对化隆县新生犊牛发生维生素缺乏症的病例研究发现,近年来犊牛先天失明发病率不断攀升,且完全治愈率较低,给当地养殖户造成了经济损失。目前研究认为犊牛失明的主要原因为机体维生素A缺乏[2-4],维生素A参与视色素的代谢演变过程,促进视紫质的再生成,还参与骨骼形成,缺乏会导致夜盲症乃至目盲症的发生,以及因颈椎骨的变形,颅腔拥塞或脑疝,导致神经受损性失明[5-6]。【前人研究进展】引起犊牛维生素A缺乏症的原因主要有2种:一是母体妊娠期间进食劣质饲料,且未进行有效的维生素补充;二是妊娠奶牛机体胃肠、肝脏功能不全,无法对脂溶性维生素进行有效吸收,母体缺乏维生素A进而导致胎儿先天缺乏[7]。研究发现,给常年饲喂干玉米秸秆,很少吃新鲜植物饲料,日粮几乎没有维生素添加剂的妊娠奶牛,连续饲喂5个多月不添加维生素的低劣青贮,新生犊牛出现了维生素A缺乏症[8]。也有人认为犊牛失明还与基因突变有关。Fujimura等[9]对26例先天性失明犊牛观察研究发现,失明犊牛出生一段时间后,视神经可能会萎缩,其认为这是由一种隐性基因引起的遗传性疾病。另外,李启飞等[10]采用氢核磁共振代谢组学方法,对1岁以下维生素A缺乏患儿与健康儿童的尿液进行代谢产物分析,发现差异代谢产物与肠道微生态平衡、消化系统疾病、呼吸系统疾病、免疫相关疾病及能量代谢、生长发育密切相关,说明代谢组学分析在维生素A缺乏早期筛查中具有潜在应用价值。【本研究切入点】目前,犊牛失明没有完全确定的病因,且研究主要集中在病理学观察[11]、原因分析、营养预防和药物治疗上[12],对其进一步的代谢产物和代谢通路的变化研究较少。【拟解决的关键问题】本文通过对同一饲喂条件下,失明犊牛及其母亲、正常犊牛进行代谢组学分析,研究失明犊牛代谢产物和代谢通路与其母亲及正常犊牛之间的异同,为进一步探究犊牛失明原因,降低犊牛先天失明的发生率,减少养殖行业经济损失提供科学依据。
1 材料与方法
1.1 试验动物与试验设计
在宁夏地区某牛场,随机选择同圈舍出生日期接近(7±2日龄)、视力正常、初生重≥40 kg正常犊牛3头,初生重≥40 kg、无意识撞物、眼前晃手不会眨眼、眼球较为突出、瞳孔散大呈蓝色的失明犊牛(7±2日龄)及其母亲(2胎以上)3对,试验分为3组:失明犊牛组(VaS)、失明犊牛母亲组(VsL)、正常犊牛组(CS),采用围栏饲养方式犊牛跟随母亲母乳喂养,产后母牛采用表1的饲料配方进行饲喂,自由饮水。

表1 产后母牛饲粮组成及营养水平(干物质基础)Table 1 Diet composition and nutritional level of postpartum cows (DM basis) (%)
1.2 样品采集
在晨饲3 h后用负压肝素钠管于犊牛颈静脉采血,失明犊牛母亲采用尾根静脉采血。血样于3000 r/min离心15 min。分装血浆于离心管中,液氮速冻5 min,-80 ℃保存。
1.3 代谢组学测定
1.3.1 样品前处理 将100 μL样品与400 μL冷甲醇乙腈(v/v,1∶1)涡流彻底混合,混合物在冰浴中超声处理1 h后,在-20 ℃下孵育1 h,最后在4 ℃下以14 000 r/min的速度离心20 min,收集上清液并在真空LC-MS分析下干燥。将干燥的提取物溶于50%乙腈中,用一次性0.22 μm乙酸纤维素过滤每个样品,并转移到2 mL HPLC瓶中,在-80 ℃储存。
1.3.2 色谱分离 整个分析过程中样品置于4 ℃自动进样器中,样品采用SHIMADZU-LC30超高效液相色谱系统(UHPLC),使用ACQUITY UPLC®HSS T3(2.1 mm×100.00 mm, 1.8 μm)(Waters, Milford, MA, USA)色谱柱。其中进样量6 μL,柱温40 ℃,流速0.3 mL/min;色谱流动相A:0.1%甲酸水溶液,B:乙腈;色谱梯度洗脱程序如下:0~2 min,B为0%;2~6 min,B从0%线性变化至48%;6~10 min,B从48%线性变化至100%;10~12 min,B维持在100%;12.0~12.1 min,B从100%线性变化至0%;12.1~15.0 min,B维持在0%。
1.3.3 质谱采集 每例样品分别采用电喷雾电离(ESI)进行正离子(+)和负离子(-)模式检测。样品经UPLC分离后用QE Plus质谱仪(Thermo Scientific)进行质谱分析,使用HESI源进行离子化,其离子化条件如下:喷雾电压:3.8 kv(+)和3.2 kv(-);毛细管温度:320(±)℃;鞘气:30(±)arb;辅助气体:5(±)arb;探头加热器温度:350(±)℃;S-lens RF水平:50%。
质谱采集时间:15 min。母离子扫描范围:70~1050 m/z,一级质谱分辨率:70 000(质荷比=200),AGC target:3e6,一级 Maximum IT:100 ms。二级质谱分析按照下列方法采集:每次全扫描(full scan)后触发采集10个最高强度母离子的二级质谱图谱(MS2 scan),二级质谱分辨率:17 500(质荷比=200),AGC target: 1e5,二级 Maximum IT:50 ms,MS2 Activation Type: HCD,Isolation window:2 m/z,Normalized collision energy(Setpped):20,30,40。
1.4 数据预处理
原始数据采用MSDIAL软件进行峰对齐、保留时间校正和提取峰面积。代谢物结构鉴定采用精确质量数匹配(质量偏差Mass tolerance<20 g/kg)和二级谱图匹配(质量偏差Mass tolerance<0.02 Da)的方式,检索HMDB、MassBank等公共数据库及拜谱本地自建的代谢物标准品库。
对提取得到的数据删除组内缺失值>50%的离子峰不参与后续统计分析;对正负离子数据分别进行总峰面积归一化,整合正负离子峰并应用R软件进行模式识别,数据经Unit variance scaling(UV)预处理后,进行后续数据分析。
2 结果与分析
2.1 样本质控分析
采用正离子模式(ESI+)和负离子模式(ESI-) ESI检测,每5个样品检测一个质量控制(QC)样品,以验证系统的可靠性。将质控样品的全离子色谱(TIC)光谱叠加比较,结果表明,质谱仪的响应强度和保留时间重叠(图1),表明该方法具有较好的稳定性和较低的由仪器误差引起的变化。峰间分离良好,说明色谱和质谱条件适合本研究样品的测定。
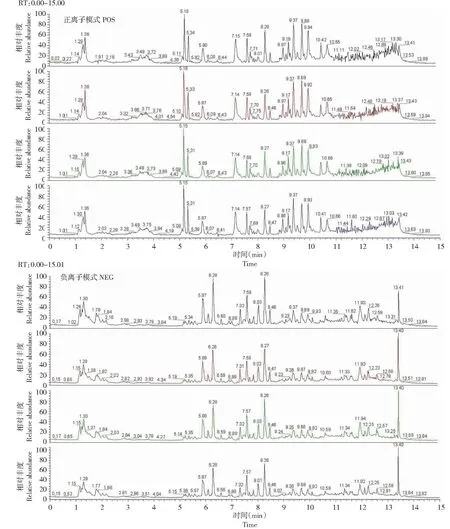
图1 正负离子模式下全离子色谱(TIC)光谱Fig.1 Total ion chromatography (TIC) spectrum in positive and negative ion mode
2.2 OPLS-DA分析
正交偏最小二乘判别分析(OPLS-DA)运用偏最小二乘回归建立代谢物表达量与样品类别之间的关系模型,来实现对样品类别的预测[13]。由OPLS-DA得分图(图2)和模型验证得分(表2)可知,OPLS-DA模型参数R2Y>0.9、Q2>0.5,说明OPLS-DA模型没有发生过拟合,模型稳定可靠。
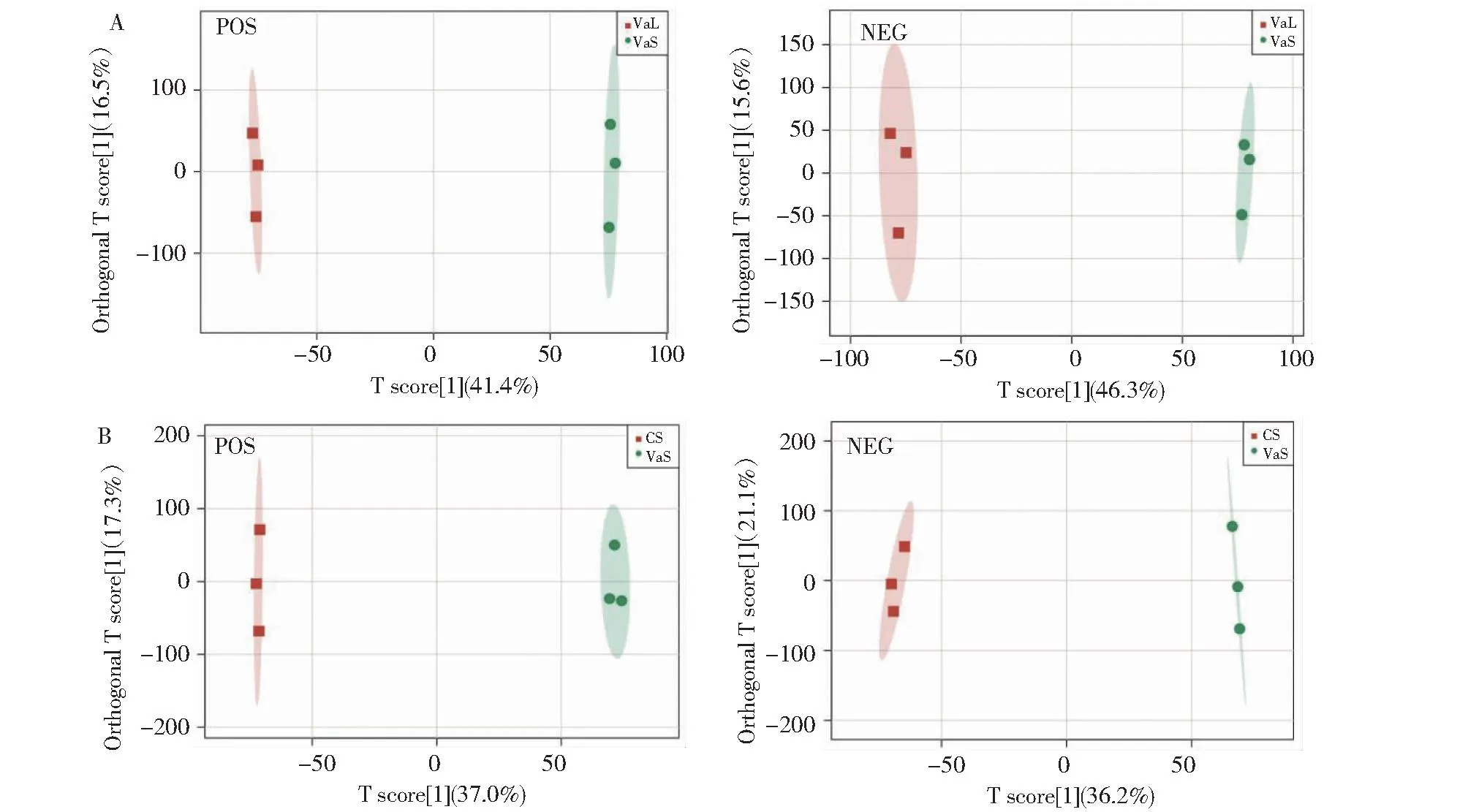
A:VaL.vs.VaS为正负离子模式OPLS-DA得分图;B:VaS.vs.CS为正负离子模式OPLS-DA得分图。A: VaL.vs.VaS is positive and negative ion mode OPLS-DA score graph; B: VaS.vs.CS is positive and negative ion mode OPLS-DA score graph.图2 OPLS-DA得分图Fig.2 OPLS-DA score chart

表2 比对组的OPLS-DA模型评价参数Table 2 Evaluation parameters of the OPLS-DA model for the comparison group
2.3 差异代谢物筛选
2.3.1 单变量统计分析 本研究共注释到667种代谢产物,其中正离子模式下399种,负离子模式下268种。将OPLS-DA的VIP值>1和单变量统计分析P<0.05作为显著性差异代谢物的标准,VaL.vs.VaS比较组共筛选出256种差异代谢物,其中正离子模式下144种,负离子模式下112种(图3-A)。VaS.vs.CS比较组共筛选出177种差异代谢物,其中正离子模式下108种,负离子模式下69种(图3-B)。

A:VaL.vs.VaS为正负离子模式火山图;B:VaS.vs.CS为正负离子模式火山图。A: VaL.vs.VaS is positive and negative ion mode volcano map; B: VaS.vs.CS is positive and negative ion mode volcano map.图3 正负离子模式火山图Fig.3 Positive and negative ion mode volcano map
2.3.2 失明犊牛与其母亲之间差异代谢物筛选 为进一步直观展示样本之间的关系和不同样本之间代谢物的表达差异,本研究对所有代谢物表达量呈显著差异的进行层次聚类(Hierarchical clustering),根据VIP值对top50差异代谢物表达量进行可视化分析。由图4可知,其中VaS.vs.VaL比较组上调的代谢物有14个,包括2-氮己环酮(2-Piperidone)、D-肉碱(D-Carnitine)、环(脯氨酸-亮氨酸)二肽[Cyclo(Leu-Pro)]、鼠李糖(Rhamnose)、脯氨酸-羟脯氨酸(Proline-Hydroxyproline)、黄单酸(Xanthomonic acid)等。表达量下调的代谢物有36个,包括4-羟基喹啉(4-hydroxyquinoline)、鞣花酸(Ellagic acid)、氢化可的松(Hydrocortisone)、甘胆酸(Glycohyocholic acid)、视紫红质(Rhodopin)、甘氨胆酸(Glycocholic acid)、二十烷酸(Icosanoic acid)、油酸甘油酯(Monoolein)等。
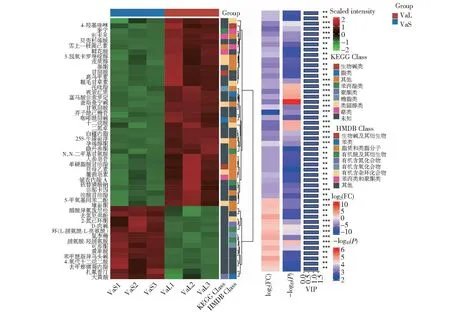
横坐标表示样本名称及分组,纵坐标表示差异代谢物。右侧的树状结构表示差异代谢物之间的相似度聚类关系,分枝越短关系越大,后面矩形依次为相对表达量、FC、P-value和VIP,颜色从绿、蓝到红表示代谢物的表达丰度从低到高,差异倍数由低到高,**代表P-value<0.005,***代表P-value<0.001。下同。The horizontal coordinates indicate the sample names and groupings, and the vertical coordinates indicate the differential metabolites. The tree structure on the right side indicates the similarity clustering relationship between the differential metabolites, the shorter the branches the greater the relationship, followed by rectangles in order of relative expression, FC, P-value and VIP, the color from green, blue to red indicates the expression abundance of metabolites from low to high, the difference ploidy from low to high,** indicates P-value<0.005, and *** indicates P-value<0.001.The same as below.图4 失明犊牛与其母亲之间差异代谢物聚类Fig.4 Differential metabolite clustering between blind calves and their mothers
2.3.3 失明犊牛与正常犊牛之间差异代谢物筛选 由图5可知,VaS.vs.CS比较组上调的代谢物有32个,包括4-吡哆醇酸(4-Pyridoxic acid)、3-(4-羟基苯基)乳酸[3-(4-Hydroxyphenyl)lactate]、鹅去氧胆酸(Chenodeoxycholic acid)、木质纤维素酸(Lignoceric acid)、脱氧胆酸(Deoxycholic acid)、LTC4、亮肽酶素(Leupeptin)、脯氨酸-羟脯氨酸(Proline-Hydroxyproline)、肾磷脂酸(Nephromopsic acid)等。表达量下调的代谢物有18个,包括L-谷氨酸(L-Glutamic acid)、亚油酸(Linoleic acid)、N-(2-呋喃基)甘氨酸[N-(2-Furoyl)Glycine]、棕榈酰乙酰胺(Palmitoyl ethanolamide)、5-羟甲基尿苷(5-Hydroxymethyluridine)、鼠李糖(Rhamnose)、脂肪酸(FA 18:3+1O)、黄嘌呤(Xanthosine)、多酚(APolyphyllin A)等。

图5 失明犊牛与正常犊牛之间差异代谢物聚类Fig.5 Differential metabolite clustering between blind and normal calves
2.4 差异代谢物功能分析
2.4.1 失明犊牛与其母亲之间差异代谢物功能分析 依据代谢物的结构与功能,对比较组筛选得到的差异代谢物进行分类统计(图6)。VaS组与VaL组相比,差异代谢物注释到KEGG数据库物质主要种类为类固醇(Steroids,18.84%)、生物碱(Alkaloids,14.49%)、萜类化合物(Terpenoids,11.59%)、多肽类(Peptides,8.70%)和PK聚酮肽(PK Polykeides,8.70%)。由图7可知,VaS组与VaL组相比,P<0.05差异代谢通路共有30个,矫正后P<0.05的差异代谢通路共有10个,主要包括胆汁分泌(Bile secretion)、初级胆汁酸生物合成(Primary bile acid biosynthesis)、戊糖磷酸途径(Pentose phosphate pathway)、维生素B6代谢(Vitamin B6 metabolism)、组氨酸代谢(Histidine metabolism)、氨基酸的生物合成(Biosynthesis of amino acids)等(表3),其中胆汁分泌代谢通路分布差异代谢物最多,与其母亲相比,失明犊牛有4个代谢物显著上调,9个代谢产物显著下调。
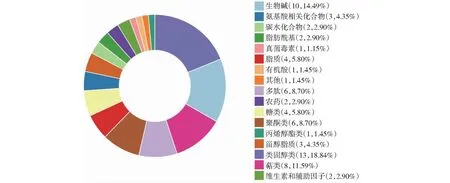
不同颜色表示KEGG一级分类,没有注释到分类信息的代谢物未被统计;图例标明分类名称、差异代谢物数量以及饼图中所占百分比。下同。Different colors indicate KEGG level 1 classification, and metabolites without annotated classification information are not counted; Legend indicates the classification name, number of differential metabolites, and percentage of the pie chart. The same as below.图6 失明犊牛与其母亲之间差异代谢物分类Fig.6 Classification of differential metabolites between blinded calves and their mothers

将pathway 归属到level 1分类,每类中的pathway从上至下-log10(P-value)依次降低,即P-value依次升高,显著性依次降低。圆圈大小表示count, 即注释到该通路中的差异代谢物数量;圆圈颜色对应校正后的P-value,由红到蓝越显著。下同。The pathway is assigned to level 1 classification, and the pathway in each category is decreased from top to bottom-log10 (P-value), i.e., P-value increases and significance decreases in order. The circle size indicates count, i.e. the number of differential metabolites annotated to the pathway; The circle color corresponds to the corrected P-value, from red to blue the more significant. The same as below.图7 失明犊牛与其母亲之间差异代谢物显著通路Fig.7 Significant pathways of differential metabolites between blinded calves and their mothers

表3 失明犊牛与其母亲之间差异代谢物分析Table 3 Differential metabolite analysis between blind calves and their mothers
2.4.2 失明犊牛与正常犊牛之间差异代谢物功能分析 依据代谢物的结构与功能,对比较组筛选得到的差异代谢物进行分类统计(图8)。VaS组与CS组相比,差异代谢物注释到KEGG数据库物质主要种类为生物碱(Alkaloids,14.89%)、脂类(Lipids,14.89%)、萜类化合物(Terpenoids,14.89%)、类固醇(Steroids,10.64%)、多肽类(Peptides,10.64%)。由图9可知,VaS组与CS组相比,P<0.05差异代谢通路共有12个,矫正后P<0.05的差异代谢通路共有16个,主要包括胆汁分泌(Bile secretion)、ABC运输工具(ABC transporters)、精氨酸生物合成(Arginine biosynthesis)、谷胱甘肽代谢(Glutathione metabolism)、氨基酸的生物合成(Biosynthesis of amino acids)等(表4),其中胆汁分泌代谢通路分布差异代谢物最多,与正常犊牛相比,失明犊牛有5个代谢物显著上调,4个代谢产物显著下调。
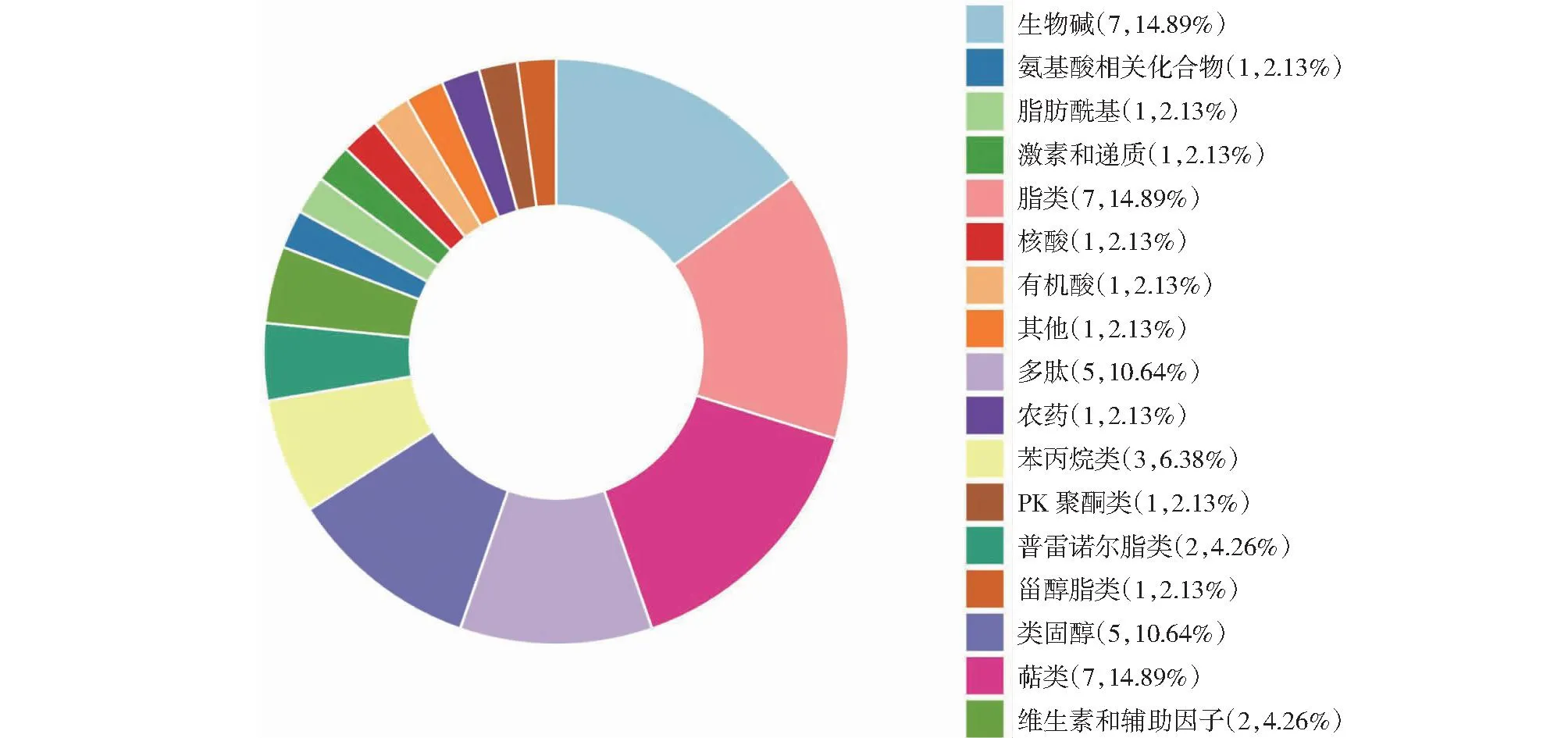
图8 失明犊牛与正常犊牛之间差异代谢物分类Fig.8 Classification of differential metabolites between blinded and normal calves
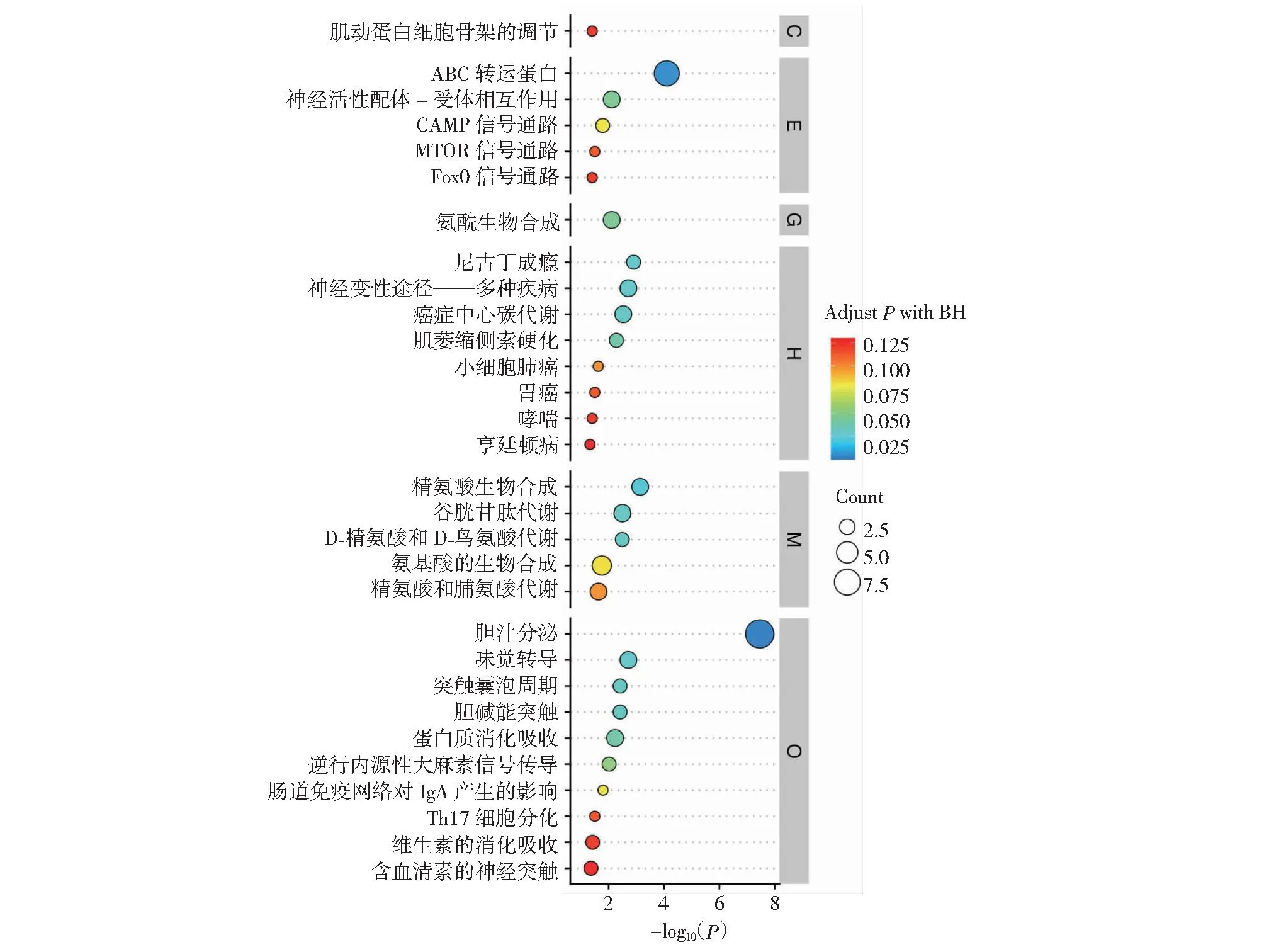
图9 失明犊牛与正常犊牛之间差异代谢物显著通路Fig.9 Significant pathways of differential metabolites between blind and normal calves

表4 失明犊牛与正常犊牛之间差异代谢物分析Table 4 Analysis of differential metabolites between blind and normal calves
3 讨 论
研究报道,视紫红质基因突变是导致视网膜色素变性最常见的原因,占常染色体显性遗传视网膜色素变性的25%~30%[14-15],已发现的视紫红质基因突变多达150余种。视紫红质(RHO)基因编码视紫红质蛋白,RHO蛋白含348个氨基酸,其C端第310~349位氨基酸可与11-顺-视黄醛形成功能性发色,当光子照射到RHO蛋白时,11-顺-视黄醛异构化,在视杆细胞中启动光转导级联反应,在将光能转换为电能的过程中起到关键性作用[16-17]。RHO基因是导致视网膜色素变性的突变基因之一。视紫红质基因突变可引起内质网应激、蛋白聚集、膜受体异常激活,从而导致视网膜色素变性。具体病理过程是视杆细胞先进行性变性,视锥细胞进而受累,最终导致视杆和视锥细胞及色素上皮丧失功能[18-19]。患病多发生于儿童、青少年,表现主要为进行性视野缺损视力下降、视网膜骨细胞样色素沉着和夜盲。徐四川等[20]报道称视紫红质蛋白是一个跨膜蛋白,视黄醛(RET)在该蛋白中的活性结合位点涉及到视觉过程机理,且与牛的一些眼科疾病有关。综上可知,机体在早期生长阶段存在着因视紫红质基因突变导致视网膜色素变性,进而引发视力受损的可能。在本研究中,失明犊牛代谢产物中的视紫红质相对含量极显著低于其母亲,且维生素A的相对含量在两者之间未呈显著差异。表明犊牛失明可能与机体视紫红质异常有关,导致光转导级联反应无法正常启动,进而影响正常视物。因为维生素A与视黄醛之间的转化存在可逆反应,但二者相对含量在失明犊牛与其母亲体内不存在显著性差异,进一步推测视紫红质的缺乏可能与基因突变存在一定关系,但具体的原因和机理有待进一步验证。
动物中的视黄醇酯和植物中的维生素A经胃蛋白酶的作用从食物中释出,在胃内几乎不被吸收,在小肠中经胆汁和胰脂酶的作用消化分解,由肠粘膜吸收。贺秀媛等[21]对维生素A缺乏症的犊牛器官进行镜检,发现维生素A缺乏会对肉犊牛的上述器官的组织结构产生不同程度的影响。同样,胆汁淤积性肝病为儿童常见肝脏疾病,胆汁调节肠道对脂肪和脂溶性维生素的吸收,因而在发生胆汁淤积时,常存在脂肪和FSVs(维生素A、D、E和K)的吸收缺陷[22]。对新生患有胆汁淤积性肝病的婴儿人为补充生理所需要的维生素A、D时并不能满足其对脂溶性维生素的需求,且不同的喂养方式对脂溶性维生素的吸收并无影响[23-24]。当肝脏和肠道有慢性疾病、饲料中缺乏维生素A或其前体胡萝卜素时也能发生维生素A缺乏症[5]。本研究发现,各比较组维生素A水平不存在显著性差异的条件下,失明犊牛和其母亲、正常犊牛相比,胆汁分泌代谢通路均呈显著性差异,其也是包含差异代谢物最多的代谢通路。另外,失明犊牛和其母亲相比,胆汁分泌初级胆汁酸生物合成代谢通路也存在显著性差异。由此认为,犊牛失明可能是机体自身存在着肝脏方面的疾病,影响胆汁的正常分泌,导致正常摄入的脂溶性维生素A不能被吸收利用,进而影响犊牛视力。
4 结 论
在机体维生素A相对含量没有显著性差异的条件下,失明犊牛和其母亲、正常犊牛相比,胆汁分泌代谢通路均呈显著性差异,其也是包含差异代谢物最多的代谢通路。另外,失明犊牛和其母亲相比,胆汁分泌初级胆汁酸生物合成代谢通路也呈显著性差异。表明犊牛失明可能与胆汁无法正常分泌,导致正常摄入的脂溶性维生素A不能被吸收利用有关。另外,失明犊牛代谢产物中的视紫红质相对含量极显著低于其母亲,且维生素A的相对含量在两者之间未呈显著性差异,表明犊牛视紫红质蛋白缺乏,可能是导致其失明的另一原因。
猜你喜欢
现代农村科技(2023年3期)2023-04-14
现代临床医学(2022年4期)2022-09-29
浙江林业科技(2022年1期)2022-02-20
科学与生活(2021年3期)2021-11-10
兽医导刊(2016年12期)2016-05-17
兽医导刊(2016年12期)2016-05-17
分析测试学报(2015年7期)2016-01-13
重庆工商大学学报(自然科学版)(2015年10期)2015-12-28
质谱学报(2015年5期)2015-03-01
当代畜禽养殖业(2014年5期)2014-08-31
