
中国亚热带杉木人工林不同林分发育阶段的群落构建机制
2024-01-23 05:45鲁旭东董禹然毛岭峰
南京林业大学学报(自然科学版) 2024年1期
鲁旭东,董禹然,李 垚,毛岭峰
(南京林业大学生态与环境学院,江苏 南京 210037)
杉木(Cunninghamialanceolata)人工林作为中国亚热带区域最为重要的人工林生态系统之一,分布面积广泛,占亚热带人工林总面积的60%~80%[1],对我国木材生产、林业经济发展具有重要战略意义。杉木人工林具有生长快、对气候土壤环境要求较高等特点[2]。大量杉木人工林的物种调查发现,其群落物种多样性随着林龄、胸径、密度的不同呈现出巨大的差异,而这一差异形成的机制目前还鲜有探索,尤其缺乏基于大尺度的研究。
群落构建的动态变化过程一直是群落生态学的一个重要主题,预测群落构建过程中的轨迹仍然是一个挑战[3]。在群落林分发育进程中生态过程的相对重要性会发生变化,即早期在群落中占据主导作用的生态过程可能逐渐不再重要[4]。因此,系统研究不同林分发育阶段群落的构建机制十分必要。群落构建过程可以通过检验群落的系统发育结构来实现:即通过将共存物种与随机抽样区域物种库生成的零模型期望进行比较[5-6]。群落系统发育结构表现为3种基本形式:①随机,其中观测值与零模型期望重叠,表明扩散和随机作用是群落构建的主导力量;②系统发育聚集,表明环境过滤是控制群落构建的主导力量,即同域分布的物种对栖息地环境的适应性是决定物种共存的主要力量;③系统发育分散,表明生物间相互作用是主导力量,即相似物种之间的拮抗相互作用导致一些分类群被排除在外,留下一个由远亲组成的群落。
对两种天然林(中国亚热带常绿阔叶林和热带森林)群落林分发育过程的研究发现,随机过程占主导作用[7-8]。但一项关于松树人工林恢复的研究发现群落构建的确定性过程更重要[9]。对于热带地区次生林的研究表明,环境过滤在群落构建中的起决定性作用[10];也有针对亚髙山草甸群落的研究发现,局域尺度内群落的构建过程不受环境影响,生物过滤作用占主导[11]。造成这些结果差异的原因可能是群落林分发育过程的早期和后期有不同的群落构建机制[12]。例如,在瑞典厄兰岛草原恢复过程中,早期表现为环境过滤,后期表现为竞争排斥[13];在新热带区3个不同林分发育阶段的实验结果表明,在发育过程中群落的净亲缘关系指数逐渐下降,即亲缘关系越来越远[14];在新几内亚的热带森林研究中发现,林分发育的早期阶段,群落系统发育结构发生由随机到聚集再到发散的变化[15]。以上研究结果都支持早期环境过滤起主要作用,后期竞争排斥作用更明显这一观点。然而,也有少数研究显示出相反的模式。如在西班牙松树重新造林过程的研究中,林分发育的早期阶段表现为随机到发散最后又趋向随机的变化[16]。因此,群落林分发育过程中生态过程的变化规律可能不统一,不同生态系统可能呈现不同的动态变化趋势。
目前,对杉木人工林的研究还主要停留在物种多样性的调查,对林分发育过程中群落构建机制动态变化的研究尚有空间。基于前期对人工林群落林分发育的研究发现:在早期阶段,群落乔木层胸径小、郁闭度低,物种要忍受高强度光照、干旱以及贫瘠土壤,因此生境过滤作用更有可能是主导群落构建的因素[17-19];在后期过程中,为了竞争相同的资源,生物间的相互作用在定居和繁殖中的作用可能会变得越来越重要[19]。因此,假设在杉木人工林发育的早期阶段,生境过滤作用使得群落系统发育结构聚集,而在林分发育的后期阶段,物种之间的竞争作用使得亲缘关系变远,群落系统发育结构发散。
了解杉木人工林群落林分发育过程中的构建机制,可以为杉木人工林在经营过程中提升局域生物多样性的稳定提供理论基础,提升生态系统服务功能。因此,本研究采用空间代替时间的方法构建时间序列,通过文献检索和野外抽样调查,收集亚热带地区林龄跨度较大(3~100 a)的143个杉木人工纯林的样方数据,以期分析中国亚热带杉木人工林群落在不同林分发育阶段群落构建主要生态过程的动态变化。
1 材料与方法
1.1 样方收集
杉木样方数据由公共数据库与实地调查两部分组成。首先从中国国家知识基础设施库(CNKI,https://www.cnki.net)中检索了所有同行评审的出版物,使用的关键字和术语如下:“地名+杉木+群落/样方”(例如:福建+杉木+群落/样方);实地调查使用了2020年于贵州省实地调查获取的杉木人工林样方数据进行补充。以下标准适用于筛选加入本研究的样方:①有完整的物种组成数据;②有具体的林龄。本研究共获取杉木人工林样方139个(表1),主要包括通过18篇文献的检索获得福建样方51个(幼龄林12个、中龄林16个、近熟林5个、成熟林5个、过熟林13个);通过10篇文献的检索获得广西样方21个(幼龄林3个、中龄林9个、近熟林9个);通过7篇文献的检索获得湖南样方24个(幼龄林6个、中龄林12个、近熟林3个、成熟林3个)。同时通过27篇文献共获得具有平均胸径数据的样方86个(胸径范围3~38 cm)。

表1 基于文献的我国各省/区杉木人工林样方数
1.2 数据处理
首先,用R 4.1.2中plantlist包[20]校对每个样方中的物种名称,对照TPL的植物名录系统进行数据标准化,修订或者剔除一些学名变动以及变种或亚种,共获得1 119种植物作为本研究的总物种库;然后,使用R软件的V.Phylomaker包[21],根据核对好的物种库,以Smith等[22]的全球维管植物系统发育史的巨型树(GBOTB.extended.tre)为骨架提取属或科级最大簇的根和基节点信息,通过“phylo.maker”函数中的场景3(scenarios=“S3”)构建新的系统发育树;最后,利用R软件的ape包[23]里“multi2di”函数对系统发育树多分枝进行随机分解,最终形成具有二分枝结构的系统发育树(图1)。

图1 中国亚热带杉木人工林总种库的系统发育树Fig. 1 Phylogenetic tree of the general seed bank of Chinese fir plantation forests of Chinese subtropical
本研究采用净种间亲缘关系指数(net relatedness index,NRI)和净最近种间亲缘关系指数(nearest taxon index,NTI)测定群落的系统发育结构。除此之外,以省界划分区域,分别作为区域物种库构建各区域系统发育树,以检验在研究区域内环境异质性和地理屏障等可能阻碍物种的传播、定植和存活的因素,以及对NTI和NRI的计算是否有显著影响。结合系统发育树,利用picante程序包[22]计算各个样方中所有物种的谱系距离(SESMPD和SESMNTD),通过picante程序包提供的零模型进行随机化,将每个样方的物种名从物种库中随机抽取999次,获取该样方中物种在随机零模型下的SESMPD和SESMNTD的分布。SESMPD等于负NRI,SESMNTD等于负NTI。NRI和NTI大于0时,表示与零模型相比群落系统发育聚集;NRI和NTI 小于 0时,表示与零模型相比群落系统发育发散;NRI和NTI 为0时,表示与零模型相比群落系统发育随机。一般认为,NRI对系统发育整体的聚集性或均匀性较为敏感,NTI对靠近进化树末端的均匀性和聚集性更为敏感。NRI侧重于从整体上描述群落中物种形成的系统发育结构,NTI侧重最近邻体的种间亲缘关系[8]。NRI(式中记为INR)和NTI(式中记为INT)的计算公式如下:
式中:DMPobserved和DMNTobserved分别为群落中种间平均遗传距离和最近种间平均遗传距离的观察值,E(DMPnull)和E(DMNTnull)为零模型平均期望值(n=999),Ds(DMPnull)和Ds(DMNTnull)为零模型的标准差值。
将林龄、胸径作为自变量,对自变量数据进行分组(林龄5 a为1组,杉木人工林为速生物种,5 a为1个龄级,超过35 a后为过熟林;胸径2 cm为1组),将其所对应的系统发育结构指数求平均值作为标准化数据,使用对数回归拟合散点图呈现杉木人工林群落在林分发育过程中的系统发育结构变化趋势。通过群落系统发育结构随林龄变化趋势以及随胸径的变化趋势两个角度探究群落生长发育过程中系统发育结构的变化趋势。数据分析和回归曲线的绘制使用R中ggplot2、ggpmisc、ggpubr软件包完成。
2 结果与分析
2.1 杉木人工林系统发育结构随林龄的变化趋势
分析发现,无论是使用总种库还是使用各区域种库,随着林龄的增大,杉木人工林植物群落的NRI和NTI均表现为先降低后平缓的趋势。根据拟合结果显示,在幼龄林前期阶段NRI >0,群落系统发育聚集,幼龄林后期期阶段NRI趋向于0,在中龄林及以后阶段NRI <0,群落系统发育发散。表明在群落整体水平上,杉木人工林植物群落随着林龄的增大,系统发育结构由初期阶段表现为聚集、在幼龄林后期转为随机过程最终逐渐趋向于发散。而近熟林阶段之前NTI >0,群落系统发育聚集,近熟林阶段NTI慢慢趋向于0,在成熟林及以后阶段NTI <0,群落系统发育发散。在进化树末端水平上,杉木人工林植物群落随着林龄的增大,系统发育结构由近熟林阶段之前表现为聚集、在近熟林阶段转为随机过程及最终逐渐趋向于发散(图2)。

图2 总种库和区域种库植物群落系统发育结构随林龄的变化趋势Fig. 2 Trends in the phylogenetic structure of plant communities with planting age in total and regional species pool
2.2 杉木人工林系统发育结构随胸径的变化趋势
分析发现,无论是使用总种库还是使用各区域种库,随着胸径的增大,杉木人工林植物群落的NRI和NTI均表现为先降低后平缓的趋势。根据拟合结果显示,NRI在生长前期胸径低于7~8 cm时大于0,群落系统发育聚集;在胸径7~8 cm慢慢趋向于0;胸径超过7~8 cm后NRI <0,群落系统发育发散。表明在群落整体水平上,杉木人工林植物群落随着胸径的增大,系统发育结构由初期阶段表现为聚集在胸径7~8 cm转为随机过程,最终逐渐趋向于发散。NTI在胸径达到20 cm之前大于0,群落系统发育聚集;在20 cm慢慢趋向于0,超过20 cm后NTI <0,群落系统发育发散。在进化树末端水平上,杉木人工林植物群落随着胸径的增大,系统发育结构在胸径20 cm之前表现为聚集,在20 cm转为随机过程,最终逐渐趋向于发散(图3)。
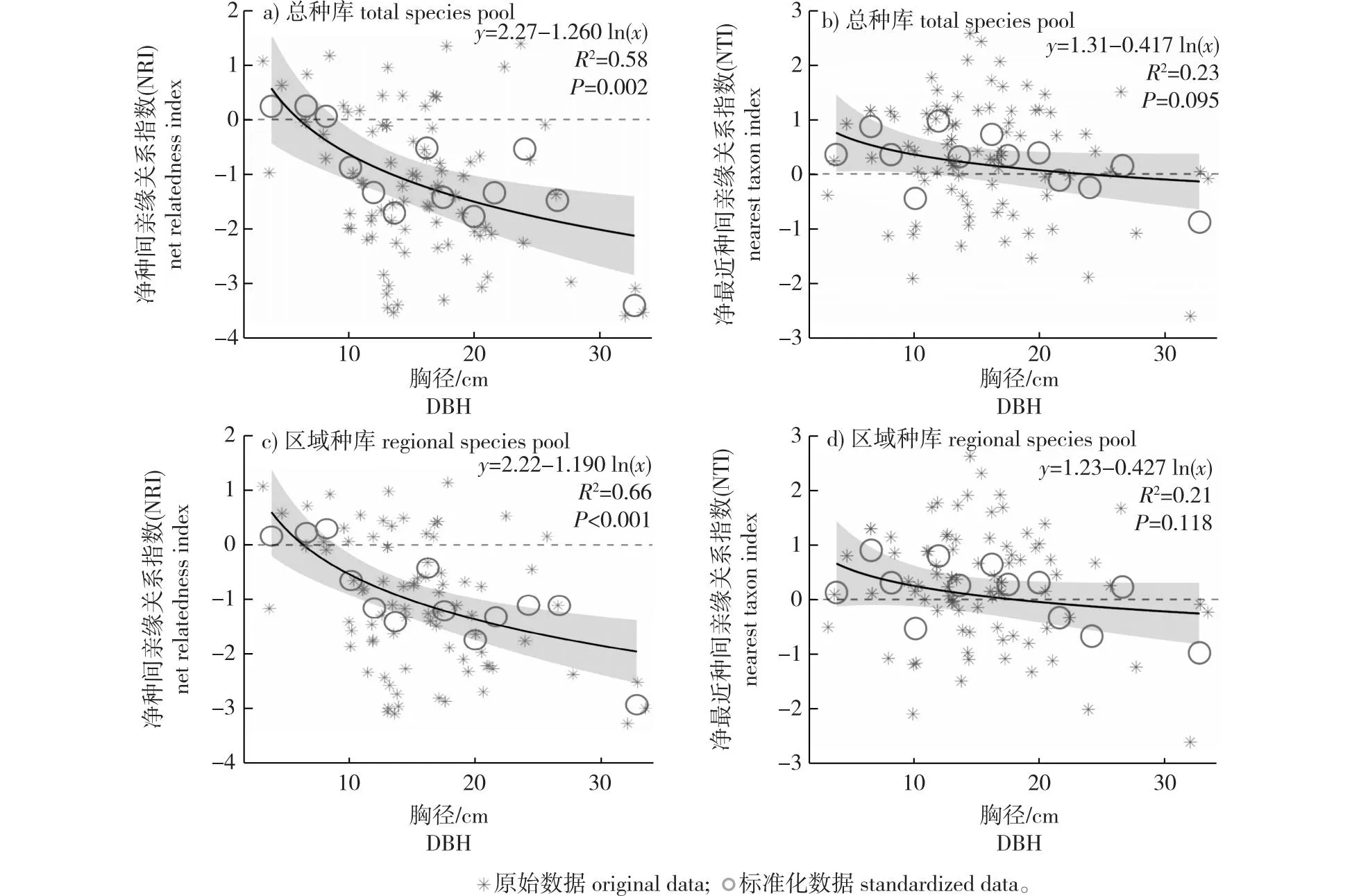
图3 总种库和区域种库植物群落系统发育结构随胸径的变化趋势Fig. 3 Trends in the phylogenetic structure of plant communities with DBH in total and regional seed bank
3 讨 论
杉木人工林植物群落的NRI和NTI随着林分生长发育而下降,这意味着植物群落生长发育过程中,系统发育结构存在由聚集向发散的方向发展的趋势。基于被子植物系统发育生态位保守性的假设,系统发育聚集源于非生物环境过滤作用,系统发育发散源于生物间相互作用[5-6,24-25]。本研究结果证明了非生物环境过滤作用在早期植物群落构建中可能占主导地位,然而随着林分发育的进行,经过一段随机过程后,生物间相互作用在群落构建中可能越来越重要[6,14,18]。
总体上,杉木人工林随着林龄的增长,群落的系统发育结构有从聚集向发散变化的趋势。NRI和NTI与样方年龄的回归曲线均具有相似的趋势。杉木人工林植物群落早期阶段,土壤种子库的发芽通常是决定初始群落结构的最重要过程,早期以快速生长的物种为主[26-27]。然而这时环境恶劣,林下先锋物种的定植需要忍受光高强度照、干旱以及贫瘠土壤,导致种子库中的一些喜阴喜湿喜肥植物难以适应局域群落的小气候环境条件生存,这时的林下物种占据相似的生态环境,具有生态相似性属于亲缘关系近的物种,这是生境过滤的结果。因此在早期阶段的群落构建中环境过滤作用占主导地位。随着林分逐渐成熟,物种丰富度高,尤其是通过扩散作用进入局域群落的物种激增,必然会产生相同生态位的物种去竞争相同的资源,从而使亲缘关系接近的物种不能共存,导致系统发育结构发散。这个阶段群落中环境资源丰富,趋向于一个基本保持稳定且相对充足的状态[26],生物相互作用在群落构建过程中变得重要。前期定植的先锋物种逐渐适应了栖息地自身所处生态位,然而随着生物扩散作用以及当地物种因为环境适宜而迅速定植,导致环境资源以及生态位的稳定会被打破,物种为了竞争相同的资源,各种内种间的相互作用愈来愈剧烈。生物间的相互作用在群落构建中变得越来越重要[19]。林分过熟后,郁闭度逐渐饱和,下层不耐阴的先锋物种逐渐死去,导致间隙形成,并且森林在光照可用性方面变得更加异质[26]。因此,环境过滤作用逐渐发挥作用,致使系统发育结构下降趋势变缓,此时生物相互作用依旧占据主导地位,但是趋向于一个稳定的水平。
但值得注意的是,尽管NRI和NTI总体趋势一致,但其变化的速率有明显的差异。在中龄林和近熟林阶段,NRI和NTI下降,但NRI <0,NTI >0,表明在这个阶段群落整体水平上表现为发散,而在进化树末端上表现为聚集。一方面,在这个阶段NRI和NTI的值较小趋近于0,确定性作用较弱,系统发育结构趋向于随机。这是可能因为杉木群落的局域小气候环境随着林龄的增加越来越适宜,杉木属于速生针叶林,随着林龄的增加上层结构变得复杂,对林下环境起到遮光保湿作用,土壤养分随着时间的积累增加[18-19]。适宜的环境导致生物多样性迅速增加,一部分来自本土物种逐渐适应局域群落的环境条件而定植,属于确定性过程;还有一部分是由于区域物种池的生物扩散作用[28-29],是随机性过程。生物扩散作用将局域群落动态与区域过程联系起来,是理解群落多样性格局不可或缺的过程[30]。扩散使得物种有机会进入不同的生境环境,并最终在其适合度较高的生境中定殖;局域群落中前期灭绝的物种,可通过扩散过程重新入侵并在群落中长期维持。当大量来自一个区域物种池的物种进入一个局域群落时,由于这些物种的定植机会相等,这个阶段中性过程在群落构建中起着主导作用[31]。另一方面,NRI更快速地向发散转变,而NTI的变化速度相对缓慢。这表明,在林分发育过程中,群落整体水平上的系统发育关系随着林龄的增加更快转向发散,而在进化树末端相对更不明显。
总体上,杉木人工林随着胸径的增长,群落的系统发育结构同样有从聚集向发散变化的趋势,并且NRI和NTI与林龄的回归曲线均具有特别相似的趋势,侧面佐证了植物群落生长发育过程中,系统发育结构存在由聚集向发散的方向发展的趋势。起始阶段胸径较小,群落环境恶劣,环境过滤作用占主导地位;随着胸径生长发育,森林结构越来越复杂,促进物种丰富度的增加,生物间的相互作用在群落构建中变得越来越重要[14,26-27]。人工林的性状会直接影响群落构建,而林龄和胸径相关性较高,本研究中杉木人工林的林龄和胸径相关系数为0.8,所以从两个视角验证了在植物群落生长发育过程中,系统发育结构存在由聚集向发散发展的趋势。
本研究对杉木人工林演替过程中群落构建动态的研究可以为杉木人工林的经营、提升局域生物多样性的稳定以及提升生态系统服务功能提供可靠的理论基础。对于刚种植的杉木人工林,杉木胸径较小,在营林过程中应当尽可能地通过施肥、浇水等措施去改善初期的环境条件,在促进杉木生长发育的同时,为物种多样性的增加提供条件。在中间阶段处于区域向杉木局域群落扩散作用的高峰阶段,保持局域群落的相对稳定更能促进杉木人工林实现生态服务与木材供应的双赢,因此不建议在这个阶段过度营林。对于过熟林,生物多样性水平高,物种间相互作用过程复杂,应当专注于其生物多样性服务价值而非经济价值,适当采取一些间伐、疏伐手段,改善林分郁闭度,促进林下生物间的相互作用,可以更好地发挥生态系统服务功能。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
防护林科技(2020年10期)2020-12-22
林业调查规划(2020年3期)2020-06-03
电子测试(2018年18期)2018-11-14
防护林科技(2016年9期)2016-09-27
防护林科技(2016年5期)2016-09-05
中国医学装备(2015年10期)2015-12-29
电测与仪表(2015年7期)2015-04-09
