
星斗山天然次生林优势种的生态位和种间联结动态
2024-02-22 11:35徐来仙艾训儒
林业科学研究 2024年1期
何 江,徐来仙,艾训儒
(1. 中国林业科学研究院林业科技信息研究所 北京 100091;2. 湖北民族大学林学园艺学院,湖北恩施 445000;3. 广西大学林学院,广西 南宁 530004)
生态位和种间联结是生态学的一个重要研究领域[1-2]。生态位指物种对环境资源利用能力及物种之间的相互作用[3],对维持物种稳定共存和促进群落演替具有重要作用[4-5]。种间联结作为反映物种空间分布的主要指标,不仅能体现物种间的竞争、共存、独立等依赖关系[6-7],还能揭示群落结构、演替和发展动态[8]。优势种对群落结构与环境的形成具有明显控制作用,它们通常个体数量多、生活能力较强和生物量高[9]。对比研究群落不同时期内优势种生态位和种间联结的动态变化,有助于深入了解群落组成、结构、功能以及主要物种间相互作用的变化[7,10],揭示群落稳定性、动态规律与演替趋势[11-12],并侧面反映群落对气候变化的响应[13]。目前国内外学者对优势种生态位和种间联结的探讨主要基于静态尺度,忽略动态尺度;现有动态研究极少,且仅局限于珍稀物种群落[14]、藤本植物[15]和喀斯特次生林[10],缺乏常绿落叶阔叶混交林的相关动态研究。
常绿落叶阔叶混交林是我国特有的天然林类型,也是亚热带山地森林的主体[11-16]。星斗山国家级自然保护区保存较完整的常绿落叶阔叶混交的天然次生林[16-17],物种丰富、群落结构复杂,且积极调节区域小气候和维持生物多样性水平。目前星斗山国家级自然保护区的研究主要涉及植物功能性状[17]、植物区系[18]、群落结构和物种多样性等[16],缺少优势种生态位和种间联结动态报道。鉴于此,本研究基于湖北星斗山国家级自然保护区固定监测样地的两次调查数据,分析重要值超过2%且排名前10 名物种的生态位和种间关系的动态变化,揭示常绿落叶阔叶混交的天然林次生林优势种对环境的适应状况、种间关系及变化规律,以期为亚热带天然次生林的生物多样性保护、森林可持续经营和管理提供参考。
1 研究区域概况
研究区为星斗山国家级自然保护区,位于鄂西南利川市、恩施市、咸丰县3 地交汇处,保护区总面积68 339 hm2,由东部星斗山(29°57'~30°10' N,108°57'~109°27' E)和西部小河(30°04'~30°14' N,108°31'~108°48' E)组成。地势呈现中部高四周低,海拔为815.0~1 751.2 m,土壤为黄壤(800 m 以下)、黄棕壤(800~1 500 m 之间)和棕壤(1 500 m 以上)。星斗山属于亚热带山地季风湿润性气候,以冬温夏热和降水丰沛为特点,年均气温12.7 ℃,最高温和最低温分别为35.4 ℃和-5.4 ℃,年均降水量1 287.11 mm,年日照时间1 298.9 h,无霜期255 d,年蒸发量1 109.2 mm。亚热带常绿落叶阔叶混交林作为典型植被类型[16]。
2 研究方法
2.1 样地设置与调查
2013 年夏季,在星斗山国家级自然保护区核心区常绿落叶阔叶混交的天然次生林典型分布地段(三县场附近),建立48 个永久固定监测样地(20 m × 20 m)。样地分布位置如图1 所示。样地设置严格遵守热带森林科学中心(Center of Tropical Forest Sciences,CTFS)的样地建立标准和调查技术规范[19]。样地信息包括经纬度、坡度、坡向、坡位和海拔(见表1),物种调查指标包括胸径(diameter at breast height,DBH)≥1 cm 木本植物的物种名、DBH、树高、相对坐标,并在DBH为1.3 m 处用红色油漆标记与挂牌;共调查到55 科117 属212 种、18 259 株,平均胸径4.64 cm,平均树高5.99 m。2018 年夏季,首轮复查样地;依据2013 年调查挂牌和树木坐标,重新测量每株木本植物的DBH、树高及生长状况(枯立、断头、倒伏、死亡等);同时,对新增的DBH ≥ 1 cm 木本植物进行标记挂牌(1 486株),记录其DBH、树高、相对坐标和生长状况;总调查到54 科110 属200 种、16 279 株,平均胸径5.37 cm,平均树高7.72 m。

图1 样地分布位置Fig. 1 Distribution position of sample plots
2.2 重要值计算
物种重要值反映物种对资源利用和适应能力[11],公式如下[20]:
式中,IV为某个物种重要值,Ra为相对密度,Rf为相对频度,Rd为相对显著度。
2.3 生态位特征
(1)生态位宽度[21]:
式中,Bi为物种i的生态位宽度,值阈为[0,logr];r为资源等级数(用样方总数代替,每个样地视为1 个资源位);nij为物种i在资源位j上的优势度(用物种重要值代替);Ni为物种i在所有资源位中重要值的总和。
(2)生态位重叠指数[22]:
式中,Oih表示物种i和物种h之间的生态重叠指数,值阈为[0, 1],且Oih=Ohi;Ni和Nh分别表示物种i和物种h在所有资源位中重要值的总和;nij和nhj分别为物种i和物种h在资源位j中的重要值。
2.4 总体联结性检验
采用方差比率法计算全部物种的总体联结性,W统计量检测总体关联性是否显著,具体公式如下[23]:
式中,VR为全部物种的总体关联系数;N为总样地数;Tj为样地j中物种数量;t为样地中物种平均数;S为总物种数;ni为物种i 存在的样地数。在独立性零假设条件下,VR期望值为1 时原假设成立,表示所有物种间无关联、相互独立;当VR>1,所有物种种间为正联结;当VR<1 时,负联结。W为W统计量,若表示物种间总体关联不显著(P>0.05),反之为显著联结(P<0.05)[24]。
2.5 种间联结性分析
(1)χ2检验值:
基于重要值排名前十的优势种在样方中是否出现(0 为未出现,1 为出现),构建种对间2 ×2 联列表;求每一种对的a、b、c、d值,其中a为2 物种同时出现的样地数,b和c仅有1 物种出现的样地数,d为2 物种都未出现的样地数。因为取样具有非连续性,用Yates连续校正公式计算χ2检验值[24]。
式中,χ2为χ2检验值;N为总样地数。ad>bc,两物种间为正联结;ad<bc,两物种间为负联结;ad=bc,相互独立。若χ2>6.635(P<0.01),两个物种呈现极显著联结;若3.841 ≤χ2≤6.635(0.01≤P≤0.05),两物种间显著联结;若χ2< 3.841(P>0.05),种对间无联结。
(2)联结系数[25]:联结系数可补充χ2检验值和反映两物种间的联结程度。
式中,AC为联结系数,-1≤AC≤1;AC越趋近1,两物种间正联结越强;AC越趋近-1,两物种间负相关越强;AC= 0,两物种之间无关联。
(3)Jaccard 指数[26]:Jaccard 指数能较准确判断物种间正联结强度,避免d值和a值偏高或偏低造成的偏差。
式中,JA为Jaccard 指数,0≤JA≤1;JA越趋向于1,两物种间关联程度越大;JA为0 时,两物种之间相互独立。
(4)Spearman 秩相关系数[24]:Spearman相关性分析适用各种数据,弥补χ2检验值结果、更敏感反映种间关系。
式中,rs(i,k)表示物种i和物种k在样地j中的Spearman 秩相关系数,取值范围为[-1, 1],正值表示正相关,负值为负相关;N为总样地数;xij与xkj分别为物种i和物种k在样地j中重要值;xi与xk分别为物种i和物种k在所有样地中的重要值平均值。
采用R4.1.3 软件的“vegan 包”计算物种重要值,“spaa 包”测定生态位和种间联结指标,“corrplot 包”分析相关性。
3 结果与分析
3.1 优势种重要值变化
如表2 所示,5 年间物种重要值大于2%且排名前10 名的物种组成差异较小,仅有1 个物种发生变化。2013年有7 个物种的生活型为乔木,1 个物种为灌木,2 个物种为灌木或乔木;2018 年时乔木、灌木、灌木或乔木3 种生活型的物种数分别为5、1、4。亮叶桦(Betula luminiferaH.Winkl.)是2013 年物种重要值排名第10 的物种,而 2018 年并未进入前 10 名; 长蕊杜鹃(Rhododendron stamineumFranch.) 属于2018 年新增的排名前十物种,物种重要值为2.80%。水榆花楸(Sorbus alnifolia(Sieb. et Zucc.) K. Koch)的物种重要值均最高,翅柃(Eurya alataKobuski)次之。光叶水青冈(Fagus lucidaRehd. et Wils.)、 包果柯(Lithocarpus cleistocarpus(Seemen) Rehderet E. H. Wilson)、 多脉青冈(Cyclobalanopsis multinervisW. C. Cheng et T. Hong)、鹅耳枥(Carpinus turczaninowiiHance) 和锥栗(Castanea henryi (Skan) Rehd. et Wils.)的重要值排序波动,多脉青冈由第5 名降为第7 名,鹅耳枥由第7 名降为第9 名,锥栗由第9 名降为第10 名(减少5.16%),光叶水青冈和包果柯排名互换、重要值都减少。齿缘吊钟花(Enkianthus serrulatus(Wils.) Schneid.)和交让木(Daphniphyllum macropodumMiq.)重要值排名未变,分别比2013 年降低了13.73%、增长了1.36%。综上,本研究选择重要值超过2%且排名前10 名物种进行后续生态位和联结性分析。
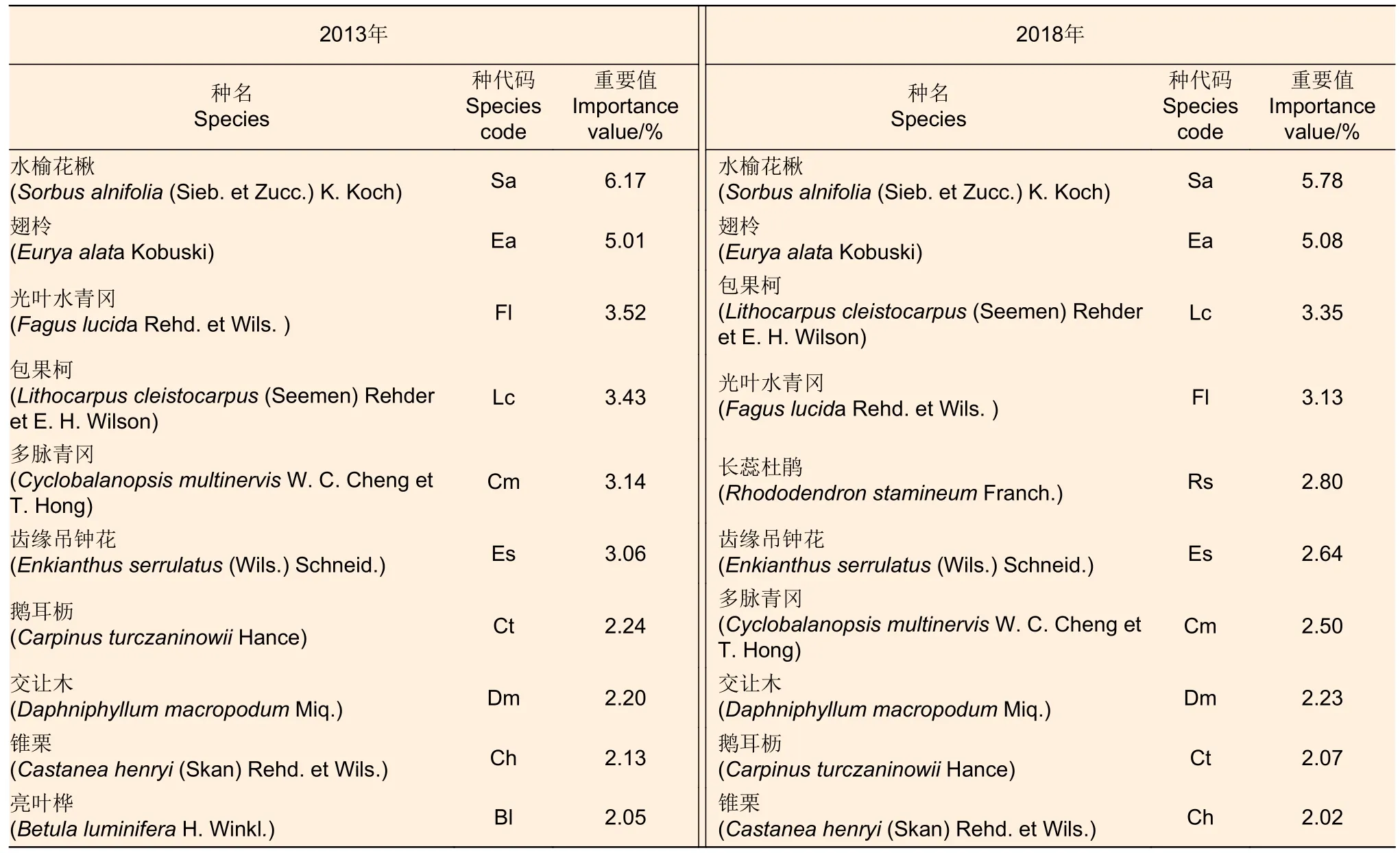
表2 2013—2018 年优势种重要值的变化Table 2 Changes in importance values of dominant species from 2013 to 2018
3.2 优势种的生态位特征变化
由图2 可知,两次调查中翅柃生态位宽度均最高,2018 年生态位宽度比2013 年提高1.21%;交让木和水榆花楸的生态位宽度位居前四,皆大于22,但生态位宽度随时间变化逐渐变窄。2018 年长蕊杜鹃的生态位宽度上升到9.33,多脉青冈和包果柯的生态位宽度分别比2013 年时增加23.07%、5.69%。亮叶桦生态位宽度在2013 年排第二(26.05),但2018 年显著降低。锥栗、鹅耳枥、齿缘吊钟花、光叶水青冈的生态位宽度呈下降趋势,比2013 年时减少4.53%、4.45%、4.61%和4.35%。齿缘吊钟花、包果柯、光叶水青冈、长蕊杜鹃的生态位宽度皆小于10,占据较窄生态位。生态位宽度与物种重要值的排序并不完全相同,两次调查中水榆花楸、翅柃的物种重要性排名前2,但翅柃生态位宽度均位居第1 名,水榆花楸排名第4(2013 年)、第3(2018 年);交让木生态位宽度排名前3 名、重要值却排第7 名。2013 年和2018 年的锥栗物种重要值分别排名第9 名、第10 名,而生态位宽度都位居第5 名。
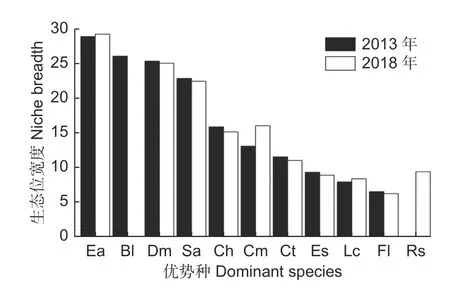
图2 2013—2018 年优势种的生态位宽度变化Fig. 2 Variation of niche breath of dominant species from 2013 to 2018
优势种对之间的生态位重叠指数存在变化(见图3),平均值由2013 年的0.30 减少到2018 年的0.29,群落整体竞争随时间推移而下降。2013年水榆花楸-亮叶桦的生态位重叠指数最高(0.60),交让木-亮叶桦、交让木-水榆花楸的生态位重叠指数次之(0.54);而2018 年交让木-水榆花楸生态位重叠指数最高(0.54),其次是翅柃-多脉青冈(0.53)、交让木-翅柃(0.52)。生态位重叠指数大于0.5 时,2013 年、2018 年种对数分别为6 对、3 对。大部分种对数的生态位重叠指数在0.2 和0.5 之间,由2013 年26 对增加到2018 年30 对,分别占总对数的57.78%和66.67%。生态重叠指数小于0.2 时,2018 年种对数比2013 年(13 对,占28.89%)减少1 对。锥栗-光叶水青冈、光叶水青冈-齿缘吊钟花的生态位重叠指数在2013 年最低(0.05),但在2018 年时锥栗-光叶水青冈上升到0.06、光叶水青冈-齿缘吊钟花未变化。亮叶桦与6 个优势种(水榆花楸、交让木、锥栗、翅柃、齿缘吊钟花和多脉青冈)的生态位重叠指数较高,竞争较激烈;长蕊杜鹃与其他物种的生态位重叠指数未超过0.5。

图3 2013—2018 年优势种生态位重叠指数变化Fig. 3 Changes in niche overlap index of dominant species from 2013 to 2018
3.3 优势种的总体联结性变化
2013 年和2018 年的优势种总体关联系数分别为1.59、1.88,皆大于1,说明整体呈现正联结,且正联结趋于增强。2013 年和2018 年的W统计量分别为76.16、90.17,W统计量不在χ20.95(48)=33.10 和χ20.05(48)=65.17 范围内,说明物种间总体联结性表现为显著正关联,且显著正关联性逐渐增加。
3.4 优势种的种间关联性变化
优势种χ2检验结果如图4 所示,正联结种对数大于负联结种对数,正联结种对数随着时间推移而增加。2018 年的正联结种对数为33(占总对数的73.33%),比2013 年增加6.45%;2018 年的负联结有11 对(占总对数的24.44%),比2013年减少15.38%。正负比由2013 年的2.38 提高到2018 年的3.00。2013 年和2018 年均只有1 对呈无联结,分别为水榆花楸-包果柯、长蕊杜鹃-光叶水青冈。2013 年有5 对呈现显著或极显著,占总对数的11.11%。水榆花楸-鹅耳枥呈极显著负联结、水榆花楸-锥栗呈极显著正联结,锥栗-齿缘吊钟花、水榆花楸-齿缘吊钟均呈显著正联结,水榆花楸-光叶水青冈呈显著负联结。2018 年显著或极显著种对数比2013 年减少1 对,且不存在显著或极显著负联结;光叶水青冈-鹅耳枥、长蕊杜鹃-齿缘吊钟花呈极显著正联结,锥栗-水榆花楸、长蕊杜鹃-锥栗呈显著正联结。五年间,水榆花楸与越来越多物种(翅柃、鹅耳枥、亮叶桦)呈现负联结,交让木与所有物种皆为正联结,翅柃仅与水榆花楸呈负联结。亮叶桦与包果柯、齿缘吊钟花、鹅耳枥、光叶水青冈、水榆花楸呈负联结,长蕊杜鹃与包果柯、齿缘吊钟花、翅柃、交让木、水榆花楸、锥栗呈正联结。

图4 2013—2018 年优势种的χ2 检验半矩阵图Fig. 4 Semi-matrix of χ2 test for dominant species from 2013 to 2018
由联结系数结果可知(见图5),当联结系数≥0.6,两次调查均有10 种对呈很强正联结。两次调查均显示翅柃-多脉青冈、翅柃-鹅耳枥、翅柃-交让木、翅柃-光叶水青冈、交让木-锥栗的联系系数为1,且随着时间推移而联结系数为1 的种对数增多(2018 年达9 对)。当0.2≤联系系数<0.6 时,种间关系表现为正联结,但联系较弱;相比于2013 年,2018 年新增3 对(长蕊杜鹃-齿缘吊钟花、长蕊杜鹃-水榆花楸、长蕊杜鹃-锥栗)的联结系数在[0.2, 0.6)之间,占总对数的22.22%。2013 年和2018 年分别有19 对、17 对种间联结系数位于(-0.2, 0.2)之间,分别占总种对数的42.22%、37.78%;随着群落发展,关系极弱甚至相互独立的种对数趋向减少。当联系系数≤-0.6 时,种间关系存在很强负联结;2013 年有5 对(亮叶桦-齿缘吊钟花、水榆花楸-翅柃、水榆花楸-鹅耳枥、水榆花楸-亮叶桦、水榆花楸-光叶水青冈),而2018 年仅剩2 对(水榆花楸-翅柃、水榆花楸-鹅耳枥)。2018 年联系系数在(-0.6,-0.2]的种对数为6 对,比2013 年增加50%,五年间呈现负联结、但联结性较弱的种对数随群落发展呈增加趋势。
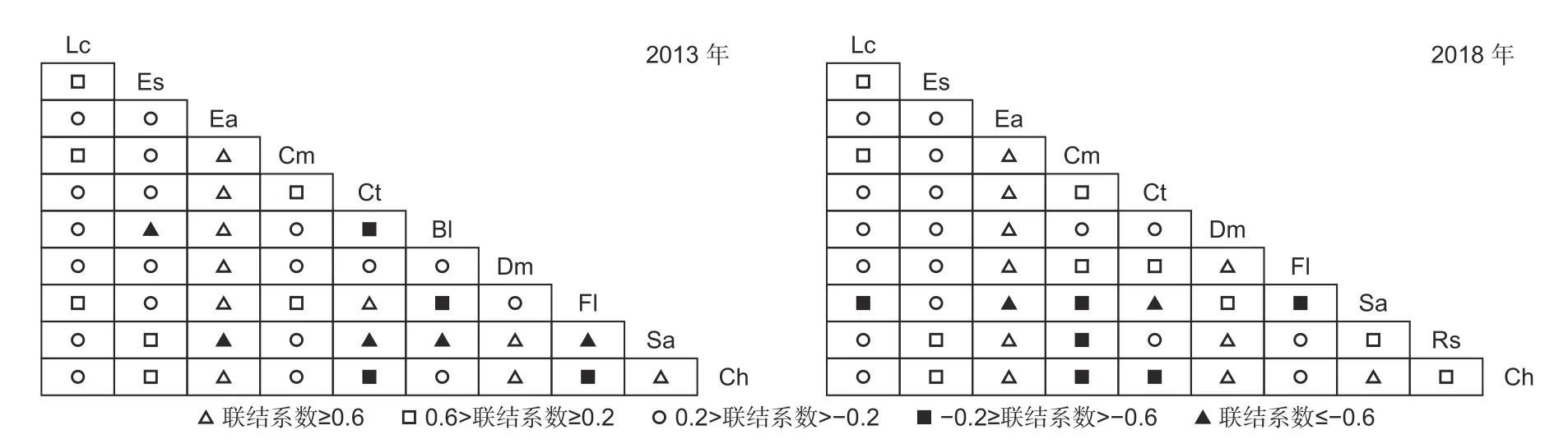
图5 2013—2018 年优势种的联结系数半矩阵图Fig. 5 Semi-matrix diagram of association coefficient of dominant species from 2013 to 2018
优势种的Jaccard 指数结果见图6。当Jaccard 指数≥0.6,两物种间存在较强联结性,两者同时出现的概率较大;2013 年有19 对,占总种对数42.22%,2018 年比2013 年减少15.79%。2018 年、2013 年分别为2 对(翅柃-交让木、交让木-水榆花楸)、5 对(翅柃-交让木、翅柃-亮叶桦、翅柃-水榆花楸、交让木-亮叶桦、交让木-水榆花楸)的Jaccard 指数均高于0.8,翅柃-交让木均达最大值。当0.6>Jaccard 指数≥0.2 时,物种之间存在较弱联结性,共同出现几率较低。2018 年有29 对的种间联结性较弱,占种对数的64.44%,比2013 年增加16.00%。2013—2018 年间,鹅耳枥与所有物种的Jaccard 指数均在[0.2, 0.6)之间,种间联结较弱;水榆花楸、交让木与其他大部分物种的Jaccard 指数高于或等于0.6,种间联结性较强。亮叶桦与翅柃、多脉青冈、交让木、水榆花楸的Jaccard 指数高于0.6,长蕊杜鹃与齿缘吊钟花、翅柃、交让木、水榆花楸的Jaccard 指数高于0.6。只有2013 年锥栗-光叶水青冈的Jaccard指数小于0.2,物种间联结性极弱,而2018 年时其Jaccard 指数达0.2。

图6 2013—2018 年优势种的Jaccard 指数半矩阵图Fig. 6 Semi-matrix diagram of Jaccard index of dominant species from 2013 to 2018
3.5 优势种的种间相关性变化
Spearman 相关系数结果表明(图7),2013年和2018 年的正相关对数均大于负相关对数,显著或极显著正相关对数高于显著或极显著负相关对数。2018 年的正相关对数为29 对(占总种对数的64.44%),2018 年比2013 年增加3.57%。正负比值由2013 年的1.75 上升到2018 年的1.93。2013 年负相关对数为16 对,2018 年比2013 年减少1 对。2013 年时水榆花楸-翅柃为无相关、交让木-包果柯呈正相关(0.06),而2018 年时分别表现为正相关(0.03)、无相关。2018 年的显著或极显著相关对数(14 对)比2013 年(16 对)减少12.50%。2013 年4 对呈极显著正相关(光叶水青冈-鹅耳枥、光叶水青冈-多脉青冈、锥栗-齿缘吊钟花、锥栗-水榆花楸),而2018 年有6 对(锥栗-齿缘吊钟花、锥栗-长蕊杜鹃、长蕊杜鹃-齿缘吊钟花、长蕊杜鹃-水榆花楸、水榆花楸-交让木、光叶水青冈-多脉青冈)。2018 年显著或极显著负相关对数分别比2013 年减少80%和50%。两次调查中,水榆花楸-鹅耳枥呈极显著负相关,水榆花楸-光叶水青冈呈显著负相关。亮叶桦与包果柯、多脉青冈、鹅耳枥、光叶水青冈呈负相关,长蕊杜鹃与多脉青冈、鹅耳枥、光叶水青冈呈负相关。
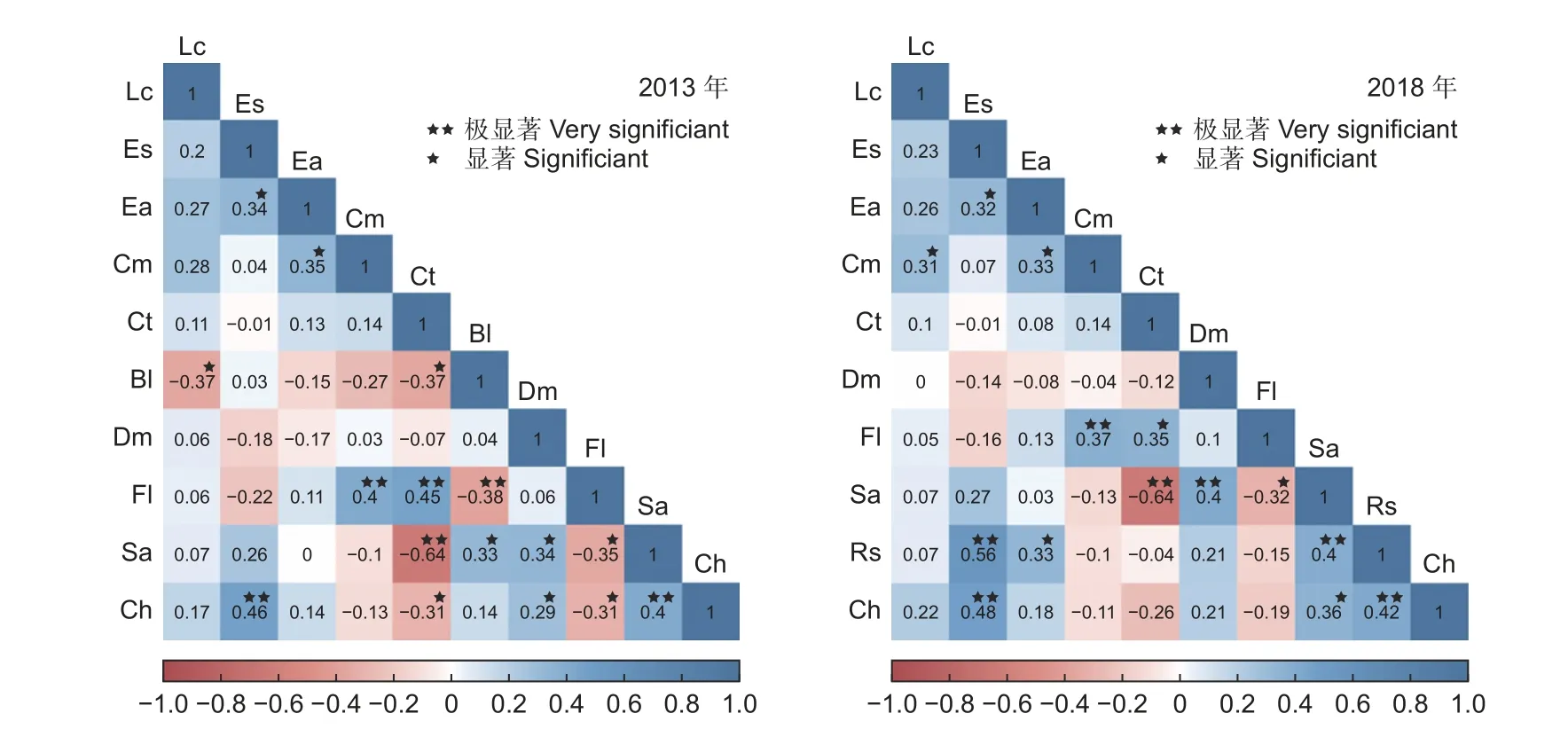
图7 2013—2018 年优势种的Spearman 相关系数及显著性Fig. 7 Correlation coefficients and significance of Spearman of dominant species from 2013 to 2018
4 讨论
4.1 优势种的生态位动态变化
物种组成是反映群落基本特征的重要指标之一。2013—2018 年间星斗山国家级自然保护区天然次生林总体物种组成变化较小,物种株数和科属种数都减少。物种组成变化与群落演替状态、受干扰情况、调查时期间隔及群落偶见种等有关[27]。由于本研究区位于自然保护区核心区,因此森林很少受人为干扰、呈自然更替状态;加之调查间隔期为5 年,群落总体动态变化有限。研究发现死亡个体以阳性落叶树种偏多,且多为较小径阶的灌木、亚乔木,可能因为伴随群落发展,森林结构逐渐复杂、郁闭度较高,导致生长力较弱小的阳性树种缺乏充足阳光和空间资源、逐步自然死亡。这与罗西[13]研究结果相似。物种重要值能体现物种在群落中的地位和作用,判断物种的优势度[28]。重要值排名前10 名的物种组成差异较小,仅有1 个物种发生变化,这表明短期间内群落不会发生明显剧烈变化[29]。长蕊杜鹃作为2018 年新增的物种重要值排名前十名的物种(第5),亮叶桦却未进2018年前十名。可能因为群落演替后生物多样性组成逐渐丰富、群落结构日益复杂和物种间竞争日趋激烈,导致喜阳性树种亮叶桦在生长、更新、竞争中处于相对劣势地位;然而,郁闭度程度较高的林分条件有利于喜阴植物长蕊杜鹃生存和更新。2013 年和2018 年水榆花楸的物种重要值均最高,相对显著度均排名第1,相对密度排名前4,但相对频度排名第6、第7,2018 年重要值呈轻微下降。这说明水榆花楸占据绝对优势地位[30],侧面反映水榆花楸为集群分布物种,且集群分布地还有其他优势物种存在。研究发现,仅翅柃、交让木、长蕊杜鹃的重要值呈小幅度上升,其它优势种的重要值呈不同程度降低,表明星斗山天然次生林中大部分优势物种间倾向均衡制约。另外,胡晓燕等[31]、姚兰等[16]、陈斯等[30]研究显示星斗山国家级自然保护区的主要优势种为交让木、多脉青冈、光叶水青冈、翅柃、亮叶桦、川陕鹅耳枥、鹅耳栎、 包果柯、 灯台树( Cornus controversa Hemsley)、 杉木( Cunninghamia lanceolata(Lamb.) Hook.)、楠木(Phoebe zhennan S. Lee et F. N. Wei)等,这与本研究排名前十名优势种大部分相似、小部分物种(楠木、灯台树、杉木)存在差异,可能因为本研究只选择了物种重要值超过2%且排名前10 名的优势种,从而造成少量重要物种被忽略。今后,有待进一步扩大研究对象和拓宽研究期限,更全面客观反映群落演替中主要物种组成、相互关系和适应环境能力等变化规律。
生态位理论解释当前群落结构建立机制[32]。2013—2018 年间翅柃的生态位宽度均最高,且逐渐变宽。加之翅柃重要值均排名第2,表明翅柃占据一定优势地位,具有较强竞争力。因为翅柃属于灌木,具有较强的耐阴和抗寒能力。交让木和水榆花楸的生态位宽度位居前四,但生态位宽度随群落发展而逐渐变窄。说明这2 个物种对星斗山自然环境条件的适应性较强,可利用资源幅度较广,但随生境条件改变,适宜能力有所减弱。长蕊杜鹃、多脉青冈和包果柯均属于常绿树种,其生态位宽度较窄,但生态位宽度随群落发展逐渐变宽,说明这3 个优势种对不断变化的环境资源利用能力不断提高,也证实部分优势常绿树种生态位宽度随群落恢复呈上升趋势[11]。随群落逐渐发展,生态位宽度下降的优势种数量是上升的优势种数量的1.75 倍,因为生境环境改变,导致物种对资源选择有所改变,加剧相似生态位物种之间竞争[13,33]。本研究中优势种的重要值排序与生态位宽度排序并不完全相同,这与闫东锋等[20]的研究结果相同。锥栗物种重要值靠后(第9 名、第10 名),生态位宽度排名前5,可能因物种生物学特性不同,应对环境变化的方式各异[34],也表明星斗山国家级自然保护区天然次生林的资源未被完全充分利用。
生态位重叠指数能反映不同物种之间对资源利用的相似程度和竞争关系[35]。该群落优势种的生态位重叠程度随群落发展呈小幅下降。可能因为随着群落演替、生境条件变化,中生性种类植物在群落中生长迅速并占据一定优势,从而排除部分竞争力较弱的树种,树种生态位进一步分化,导致总体上物种间关系缓和[11]。生态位越宽,物种对环境的适应力越强,物种之间也存在较高的生态位重叠[32,36]。研究发现,翅柃与其他大部分优势种的生态位重叠指数较高,以翅柃-多脉青冈、翅柃-交让木之间竞争最大。生态位宽度均最窄的光叶水青冈与其他优势种的生态位重叠程度相对较低,可能因为生态位宽度小的物种对资源需求存在差异。在两次调查中,交让木-水榆花楸的生态位重叠指数均很高(前2 名),两者对资源竞争较大,物种生态位趋于泛化[10]。另外,亮叶桦与水榆花楸、交让木、锥栗、翅柃、齿缘吊钟花、多脉青冈的生态位重叠指数较高、生态习性相似,竞争激烈。大多数种对数的生态位重叠指数在0.2 和0.5 之间,且种对数随着群落演替发展呈现上升趋势,说明大部分物种对生境需求具有一定相似性、互补性。
4.2 优势种的种间联结动态变化
总体联结性揭示群落演替阶段、群落结构和物种组成的稳定性[36-37]。当群落趋向于顶级演替时,群落越趋向正联结,群落结构也越完整,且物种间充分利用资源环境,相互促进生长[38-39]。2013—2018年星斗山国家级自然保护区天然次生林优势种的总体联结性均呈现出显著正关联,且显著正关联性逐渐增加。同时,正联结种对数大于负联结种对数,且正联结种对数随群落演替而增加,侧面反映出该群落物种间的生态习性相似,对生境资源需求具有互补性。
本研究综合采用χ2检验、联结系数和Jaccard指数判定群落种对联结性,以更准确阐明物种间的关联程度[40]。2013—2018 年间优势种的χ2检验、联结系数和Jaccard 指数基本一致:正联结种对数大于负联结种对数,不显著联结种对数也占有重要比例,且正负比随群落演替而逐渐增加。这表明星斗山国家级自然保护区天然次生林处于自然演替的早期至中期阶段,种间联结松散、不紧密[10]。翅柃-交让木一直表现为很强正联结,因为它们均耐阴、抗寒,属于灌木或小乔木。亮叶桦与水榆花楸、齿缘吊钟花等5 个物种的χ2检验结果为负联结,与齿缘吊钟花、水榆花楸的联结系数均小于-0.6,与水榆花楸等4 物种的Jaccard指数较高。这说明亮叶桦与齿缘吊钟花、水榆花楸等是竞争或互斥关系,与其他正联结物种(翅柃等)可能为互利或为无害关系。可能因为亮叶桦与齿缘吊钟花、水榆花楸等物种对环境资源的需求相近,故因争夺生存资源而互斥,从而在一定程度上解释了第二次调查时亮叶桦物种重要值未属于前十名的原因。χ2检验结果显示,长蕊杜鹃与翅柃、交让木等6 物种呈现正联结,与齿缘吊钟花、锥栗呈极显著或显著正联结;长蕊杜鹃与翅柃、交让木的联结系数达到1,与齿缘吊钟花、翅柃、交让木等4 物种的Jaccard 指数大于等于0.6。这说明长蕊杜鹃与齿缘吊钟花、翅柃、交让木等多数物种存在较强的互利互助关系。
Spearman 秩相关系数能更灵敏、准确反映群落中物种间关联程度[2],能弥补χ2检验值的不足[24]。Spearman 结果与χ2检验、联结系数、Jaccard 指数和总体关联性结果基本类似,这些指标优势互补、共同反映出星斗山国家级自然保护区天然次生林的真实现状。鉴于计算多个种间联结指标的内容繁杂、结果相似,建议今后加入综合性方法(如赋权重等)进行研究,以更直观、清晰地呈现主要物种的种间关系。水榆花楸-翅柃由2013 年的无相关转变为2018 年的正相关;交让木-包果柯却由正相关转变为无相关。这说明植物种间关联性不仅与自身生物学特性密切相关,还受生存环境等多种因素(地形、土壤、海拔、坡度、光照、水分、温度等)的影响[2]。鹅耳枥-水榆花楸的负相关程度最强,因为两者属于落叶乔木,喜光,耐阴,耐寒,耐干旱瘠薄,生物学特性相似;它们因竞争有限资源而相互排斥,也反映了物种间的生态位分离[41]。
5 结论
2013—2018 年星斗山国家级自然保护区天然次生林总体物种组成变化较小,多数优势种的物种重要值随群落发展而降低。两次固定样地调查期内大部分物种对资源利用能力随群落发展呈下降趋势、生态位宽度随群落发展逐渐变窄,多数物种对生境需求具有一定相似性、互补性;随群落发展,总体联结性呈现越来越强的显著正关联,正相关对数和正负比值呈小幅度增加,负相关对数逐渐减少,不显著联结对数占有重要比例;水榆花楸和翅柃优势显著、竞争力极强,它们与越来越多物种呈现出较强负联结或正联结。总体上,种间联结松散、不紧密,群落处于自然演替的早期至中期阶段。建议及时关注群落结构变化,注重保护环境特化物种与珍稀物种资源,防止优势种扩张导致群落特有种减少。此外,环境因子对植物生长和物种间关系具有重要影响,今后还需要进一步研究环境因子的变化(如土壤、气温等),并结合植物生理生化、遗传基因、长期动态监测等全面探讨群落演替过程中种间关系的变化规律和形成机制。
猜你喜欢
植物研究(2023年5期)2023-09-09
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
数学物理学报(2022年2期)2022-04-26
中国野生植物资源(2022年3期)2022-04-19
新世纪智能(数学备考)(2021年9期)2021-11-24
新世纪智能(数学备考)(2020年9期)2021-01-04
贵州农业科学(2020年1期)2020-03-25
植物资源与环境学报(2019年3期)2019-09-12
中学生数理化·高一版(2018年10期)2018-11-08
西南农业学报(2016年5期)2016-05-17
