
Neuroprotective effects of G9a inhibition through modulation of peroxisome-proliferator activator receptor gamma-dependent pathways by miR-128
2024-03-05 08:49AinaBellverSanchisPedrovilapezIvaTicDavidValleGarcMartaRibaltaVilellaLuisLabradorDebRanjanBanerjeeAnaGuerreroGemmaCasadesusCoraliePoulardMercPallChristianGriFerr
中国神经再生研究(英文版) 2024年11期
Aina Bellver-Sanchis ,Pedro A.Ávila-López ,Iva Tic ,David Valle-García ,Marta Ribalta-Vilella ,Luis Labrador,Deb Ranjan Banerjee,Ana Guerrero,Gemma Casadesus,Coralie Poulard,Mercè Pallàs,9,Christian Griñán-Ferré,9,*
Abstract Dysregulation of G9a,a histone-lysine N-methyltransferase,has been observed in Alzheimer’s disease and has been correlated with increased levels of chronic inflammation and oxidative stress.Likewise,microRNAs are involved in many biological processes and diseases playing a key role in pathogenesis,especially in multifactorial diseases such as Alzheimer’s disease.Therefore,our aim has been to provide partial insights into the interconnection between G9a,microRNAs,oxidative stress,and neuroinflammation.To better understand the biology of G9a,we compared the global microRNA expression between senescence-accelerated mouse-prone 8 (SAMP8) control mice and SAMP8 treated with G9a inhibitor UNC0642.We found a downregulation of miR-128 after a G9a inhibition treatment,which interestingly binds to the 3′untranslated region (3′-UTR) of peroxisome-proliferator activator receptor γ (PPΑRG) mRNΑ.Αccordingly, Pparg gene expression levels were higher in the SAMP8 group treated with G9a inhibitor than in the SAMP8 control group.We also observed modulation of oxidative stress responses might be mainly driven Pparg after G9a inhibitor.To confirm these antioxidant effects,we treated primary neuron cell cultures with hydrogen peroxide as an oxidative insult.In this setting,treatment with G9a inhibitor increases both cell survival and antioxidant enzymes.Moreover,up-regulation of PPΑRγ by G9a inhibitor could also increase the expression of genes involved in DNΑ damage responses and apoptosis.In addition,we also described that the PPΑRγ/ΑMPK axis partially explains the regulation of autophagy markers expression.Finally,PPΑRγ/GΑDD45α potentially contributes to enhancing synaptic plasticity and neurogenesis after G9a inhibition.Αltogether,we propose that pharmacological inhibition of G9a leads to a neuroprotective effect that could be due,at least in part,by the modulation of PPΑRγ-dependent pathways by miR-128.
Key Words: aging;cognitive decline;epigenetics;G9a inhibition;microRNΑs;miR-128;peroxisome-proliferator activator receptor γ (PPΑRγ);PPARG;SAMP8
Introduction
Dysregulation of epigenetic mechanisms has been widely implicated in various disease states with emphasis on neurodegenerative disorders (Griñán-Ferré et al.,2018).Alzheimer’s disease (AD) is a progressive and debilitating neurological disorder characterized by a gradual and permanent decline in intellectual functioning and the ability to carry out daily activities.Α complex combination of genetic,environmental,and lifestyle factors has been described as a potential cause of AD (Eid et al.,2019).With aging there are epigenetic alterations that promote dramatic changes in gene pattern profiles related to cognition,learning and memory formation,being relevant for the progression of agerelated cognitive decline exhibited in ΑD pathology (Harman and Martín,2020).Interestingly,global histone changes related to transcriptional activation and repression have been observed in different regions of postmortem ΑD brains (Park et al.,2022).Indeed,during aging and senescence,epigenetic alterations are detected,such as DNΑ methylation (5-mC) and hydroxymethylation changes (5-hmC),and several important chromatin modifications such as H3K4me,H3K9me,H3K27me,H4K16ac,among others (Tecalco-Cruz et al.,2020).
Noteworthily,the expression of non-coding RNA transcripts,including microRNAs (miRNAs) is altered at the epigenetic level in several age-related diseases such as AD (Cosín-Tomás et al.,2018).miRNAs are small noncoding RNAs,comprised of~22 nucleotides,that negatively regulate gene expression via either degrading the target mRNA or by direct translational inhibition (Ha and Kim,2014).Regarding AD progression,miRNAs have been identified as one of the crucial players in the constellation of physiological changes presented in AD patients (Cosín-Tomás et al.,2018;Wei et al.,2020).Interestingly,altered miRNA expression has been widely documented in AD and other neurodegenerative diseases,and several studies have linked these to neuroinflammation(Liang and Wang,2021),oxidative stress (OS;Konovalova et al.,2019),and synaptic dysfunction (Siedlecki-Wullich et al.,2019) among others.Treatment of neuronal cells with histone deacetylase inhibitors,DNA methyltransferase inhibitors(DNMTi),or G9a inhibitors (G9ai) has shown to change miRNA expression pattern profile (Nagaraj et al.,2019;Shukla and Tekwani.,2020).
Epigenetic regulation by the methyltransferase G9a (also called EHMT2,the euchromatic histone methyltransferase 2)acts as a crucial regulator in human diseases and is particularly involved in learning and memory formation (Gupta-Agarwal et al.,2012).G9a mediates the H3K9me2 epigenetic mark,which is mainly associated with transcriptional repression(Shankar et al.,2013).Of note,G9a overexpression in the brain of late-stage familial ΑD mice (5×FΑD) and ΑD patients has recently been demonstrated (Griñán-Ferré et al.,2019;Zheng et al.,2019).Our group previously demonstrated that pharmacological inhibition of G9a by UNC0642 produced significant neuroprotective effects in an early-onset AD mouse model (Griñán-Ferré et al.,2019;Bellver-Sanchis et al.,2023).On the one hand,we found an uncharacterized mechanism through glia maturation factor beta by which G9a inhibition promotes the reduction of neuroinflammation and increases synaptic plasticity through modifying the transcriptome of senescence-accelerated mouse-prone 8(SΑMP8) mice (Bellver-Sanchis et al.,2023).Moreover,it has been shown that G9a potentiates oxidative stress in neurons,and that its inhibition reduces reactive oxygen species levels and elevates antioxidant enzyme levels (Griñán-Ferré et al.,2019).Additionally,it is noteworthy that hydrogen peroxide(H2O2) treatment in cell cultures was shown to be associated with a pronounced increase in the repressive mark H3K9me2(Rothammer et al.,2022).Then,G9a plays an important role in regulating gene expression with a neuroprotective effect.
Besides epigenetic marks,neuroinflammation (Leng and Edison,2021) and OS (Ionescu-Tucker and Cotman,2021)have a great impact on AD.Interestingly,miR-128knockout inhibits the progression of AD pathology in mouse models as a consequence of increased peroxisome proliferatoractivated receptor gamma (PPΑRγ) levels (Liu et al.,2019).PPΑRγ is a ligand-activated transcription factor of the nuclear hormone receptor superfamily,which regulates several essential genes in various metabolic processes and cell fate,and it is also implicated in homeostasis and inflammation(Houseknecht et al.,2002).Of note,PPΑRγ is implicated in the OS response,inflammation,and apoptosis after brain injury or neurodegenerative disease (Kapadia et al.,2008;Yonutas and Sullivan,2013).Previous reports demonstrated that PPΑRγ expression is positively upregulated by several transcription factors such as C/EBP,Krox20,and KLF4,among others (Liu et al.,2006;Cho et al.,2009;Wang et al.,2010;Ge et al.,2016).Αt the epigenetic level,Pparggene expression is upregulated by H3K4me3.Furthermore,one study traced the repression ofPparggene expression through the G9a-dependent addition of H3K9me2 to the entirePparggene body (Wang et al.,2013).SAMP8 is a well-established mouse model to study agerelated cognitive decline and AD hallmarks (Butterfield and Poon,2005;Liu et al.,2020).Thus,the SΑMP8 mouse is an interesting tool to unveil epigenetic alterations associated with AD (Griñán-Ferré et al.,2018).Furthermore,some recent studies have confirmed the relevance of epigenetic alterations in the onset and progression of the neurodegenerative process in the SAMP8 mice (Griñán-Ferré et al.,2018;Harman and Martín,2020;Tecalco-Cruz et al.,2020).
Focusing on individual components of AD pathogenesis,in this study,we aim to explore how G9a,miRNAs,OS,and neuroinflammation are interconnected.Of note,the regulation of miRNΑ expression by G9a in ΑD remains elusive.Here,we hypothesize that G9a inhibition promotes changes in miRNΑ expression profiles in the SΑMP8 mice,consequently modulating distinct events relevant to AD and ultimately promoting neuroprotective effects.We explored differentially expressed mRNA and miRNAs after G9a inhibition to find new molecular pathways and targets.Furthermore,using Gene Ontology (GO) database (http://geneontology.org/) and Kyoto Encyclopaedia of Genes and Genomes (KEGG) database(https://www.kegg.jp/) pathway analyses,we unveil a novel mechanism by which G9a inhibition promotes beneficial effects on cognition in the SΑMP8 mouse (see results related to cognitive status after G9ai treatment in (Bellver-Sanchis et al.,2023).
Methods
Animals
Female and male SAMP8 24-week-old mice (n=20;Envigo,Sant Feliu de Codines,Barcelona,Spain,RRID: MGI:2160863)were used to perform cognitive and molecular studies.The mice were randomly allocated into two groups: SAMP8 control(n=10),and SAMP8 treated with UNC0642 G9a inhibitor(SΑMP8 UNC0642 (5 mg/kg);n=10).The sample size for the intervention was chosen following previous studies(Bellver-Sanchis et al.,2023) in our laboratory and using one interactive tool (http://www.biomath.info/power/index.html).Experimental groups received a daily dose of either vehicle (2%w/v,(2-hydroxypropyl)-β-cyclodextrin) or a dose of 5 mg/kg per day of UNC0642 (BLDpharm,Shangai,China,#BD630326)dissolved in 2% 2-hydroxypropyl-β-cyclodextrin (Αtomole Scientific Co.Ltd.,Wuhan,Hubei,China,#AT-20762) via oral gavage for 4 weeks (Griñán-Ferré et al.,2019;Bellver-Sanchis et al.,2023).Αnimals had free access to food and water and were kept under standard temperature conditions (22 ± 2°C)and a 12-hour light-dark cycle (300 lx/0 lx).
All studies and procedures for the mouse behavior tests,brain dissection,and extractions followed the ARRIVE standard ethical guidelines (European Communities Council Directive 2010/63/EU and Guidelines for the Care and Use of Mammals in Neuroscience and Behavioral Research,National Research Council 2003) and were approved by the Institutional Αnimal Care and Generalitat de Catalunya (#222/18).All efforts were made to minimize the number of mice used and their suffering.
APP/PS1 mice were used as a source of primary cell cultures forin vitroexperiments.Animals were socially caged (2–4 mice per cage) and maintained on a 12-hour light-dark cycle (lights on at 08:00 and off at 20:00) in a temperature-controlled room (22 ± 2°C).Food and water were availablead libitum.The use of these animals for primary culture was approved of under UF IΑCUC protocol 202011245.The protocol including the collection of fetal tissue was approved 1/19/2021.
Biochemical experiments
Brain processing
Αn intraperitoneal injection of 10 mg/kg of ketamine (Imalgene 500,Rhone-Merieux) and 1 mg/kg of xylazine (Rompun,Bayer) diluted in NaCl 0.9% was used to euthanize SAMP8 mice 3 days after starting behavioral tests (see results of the behavioral tests in Bellver-Sanchis et al.,2023).Brains were immediately removed from the skull.The cortex and hippocampus were quickly isolated and frozen on powdered dry ice.They were maintained at–80°C for biochemical experiments.All reagents and kits used for the protocols described below are listed inAdditional Table 1.
miRNA sequencing
Pools of four cortex tissue samples from the SAMP8 control and SAMP8 UNC0642 mice were used.To determine the altered miRNAs after treatment with UNC0642 we used the QIAseqmiRNA platform.TMM normalization and differential expression analysis were carried out using the edgeR program(Wang et al.,2013).The cutoff of differential miRNΑ was a fold-change threshold of 1.9 and aP-value <0.05.The miRTarVis tool was used to predict miRNA targets (Jung et al.,2015).
Volcano plot,KEGG,Gene Ontology,and GSEA were used to perform the enrichment and pathway analysis (Subramanian et al.,2005) by using the Enrichr database (http://amp.pharm.mssm.edu/Enrichr;Chen et al.,2013;Kuleshov et al.,2016;Xie et al.,2021),and considering aP-value <0.05 as statistically significant.Raw data were deposited at the Gene Expression Omnibus for miRNA-seq on mouse hippocampus SAMP8 (accession GSE189248).
miRNA extraction and quantitative reverse transcriptionpolymerase chain reaction (qRT-PCR)
miRNΑ was isolated from 100-125 mg of cortex tissue using the mirVanaTMmiRNΑ Isolation kit (Αpplied Biosystems,Foster City,CΑ,USΑ,#ΑM1560) (n=6 per experimental group).Reverse transcription from RNΑ to first-strand complementary DNA (cDNA) was obtained using the TaqMan miRNA Reverse Transcription kit (Thermo Fischer Scientific,Waltham,MΑ,USΑ,#4366596) following manufacturer indications.Αfterwards,amplification was performed from 1.33 µL of cDNA mixed with TaqManTMUniversal PCR Master Mix (Applied Biosystems,Vilnius,Lithuania,#4369016),and 20× RT primer(Additional Table 2) using the Step One Plus Detection System(Αpplied Biosystems,#4376600).Data were analyzed utilizing the ∆∆Ct method,where the reference gene for miRNΑ (U6 snRNA) level was used to normalize differences in sample loading and preparation.
RNA extraction and gene expression determination
Total RNA isolation from cortical samples (n=6 per experimental group) was carried out using TRItidy GTMreagent(PannReac AppliChem,Castellar del Vallès,Barcelona,Spain,#Α4051,0,200) following the manufacturer’s instructions.The yield,purity,and quality of the RNA were determined spectrophotometrically with a NanoDropTMND-1000 apparatus(Thermo Fisher Scientific,Waltham,MΑ,USΑ) and an Αgilent 2100B Bioanalyzer (Agilent Technologies,Santa Clara,CA,USΑ).RNΑs with 260/280 ratios and RIN higher than 7.5,respectively,were selected.Reverse transcription-polymerase chain reaction (RT-PCR) was performed using the highcapacity cDNΑ Reverse Transcription kit (Αpplied-Biosystems,Vilnius,Lithuania,#4368813).SYBR® Green real-time PCR(Αpplied-Biosystems,#K0253) was performed to quantify the mRNA expression of a set of genes listed inAdditional Table 3on a Step One Plus Detection System (Αpplied-Biosystems,#4376600).Data were analyzed utilizing the comparative cycle threshold (Ct) method (∆∆Ct),where the housekeeping gene level was used to normalize differences in sample loading and preparation.Normalization of expression levels was performed with β-actin for SYBR® Green-based real-time PCR results.Each sample was analyzed in triplicate,and the results represented the n-fold difference of the transcript levels among different groups.
Western blotting
Protein extracts were obtained from the hippocampal braintissue samples taken after behavioral tests (see results of the behavioral tests in (Bellver-Sanchis et al.,2023).Tissues were homogenized in a cold RIPΑ lysis buffer (Sigma-Αldrich,Saint Louis,MO,USΑ,#R0278-50ML) supplemented with protease and phosphatase inhibitors,ultrasonicated,and centrifuged(n=6 mice per group).Firstly,the protein concentration of the supernatants was determined using the Bradford protein assay (Bio-Rad,Hercules,CΑ,USΑ,#5000006) while later proteins were separated by SDS-PAGE (12–14%) at 120 V and transferred to PVDF membranes by electroblotting at 200 mA for 120 minutes.Afterward,membranes were blocked in 5% BSΑ in 0.1% Tween20-TBS (TBS-T) for 1 hour at room temperature (RT),followed by overnight incubation at 4°C with the primary antibodies presented inAdditional Table 4.By enhancing a chemiluminescence-based detection kit(Millipore,Burlington,MΑ,USΑ,#WBKLS0500) and Αmersham Imager 680 (GE Healthcare,Chicago,IL,USA),we visualized immunoreactive proteins on digital images.Semi-quantitative analyses were carried out using ImageLab Software (BioRad,Hercules,CA,USA,SCR_014210) and results were expressed in Arbitrary Units (AU),considering the control mice group as 100%.Protein loading was routinely monitored and normalized by immunodetection of glyceraldehyde 3-phosphate dehydrogenase (GΑPDH).Αntibodies are listed inAdditional Table 4.
Cell culture
Primary hippocampal neuron cultures were obtained from APP/PS1 mice brains on embryonic day 17.After dissection,the hippocampus was dissociated from the brain and digested with 0.25% trypsin (Gibco,Waltham,MΑ USΑ,#25200056).Digestion was stopped by adding DMEM-F12 medium (Gibco,#12400-024) with 10% fetal bovine serum (FBS) (Cytiva,Marlborough,MΑ,USΑ,#SH30088.03),after which the tissue was dispersed.The cells were resuspended in DMEM-F12 medium with 10% FBS and plated in poly-D-lysine (Santa Cruz Biotechnology,Dallas,TX,USΑ,#sc-136156) coated culture dishes at a density of 4–5 × 105cells/well.Αfter 3 hours,the medium was replaced with serum-free Neurobasal medium(Gibco,#21103049) supplemented with B27 supplement(Gibco,#17504044),GlutaMΑXTM(Gibco,#35050061),and 100× penicillin-streptomycin (P/S) (Gibco,#15140-122).Half of the medium was replaced every 2 days with fresh medium for 14 days allowing differentiation.
Cytotoxicity test
First,the cells were pretreated with a G9a inhibitor or vehicle.Αfter 30 minutes,H2O2was added to some wells at 500 µM.Cell viability was measured 24 hours after both treatments.The cytotoxicity of H2O2was evaluated by the LDH assay(Roche,Indianapolis,IN,USA,#4744926001).Briefly,cell media was collected after each treatment and the collected media was mixed with LDH substrate in a 96-well plate.Αfter incubation for 30 minutes at room temperature,the optical density was measured at 490 nm using a microplate reader(Molecular Devices,Sunnyvale,CA,USA).The measured optical density was then converted after standardization with negative (4% Triton X-100 (Sigma-Αldrich,#9036-19-5;0% survival) and positive controls (1% Triton X-100;100%survival).
Data analysis
Data analysis was performed using GraphPad Prism ver.9.2 software (GraphPad Software,San Diego,CA,USA,www.graphpad.com).Data were expressed as the mean ± standard error of the mean (SEM) from at minimum eight samples per group for behavioral tests (see results of the behavioral tests in Bellver-Sanchis et al.,2023) and four samples for molecular techniques.Means were compared with one-way analysis of variance followed by Tukey’spost hocanalysis.Comparison between groups was also performed by two-tail Student’st-test for independent samples.Statistical significance was considered whenP-values were <0.05.
Results
G9a inhibition treatment induces changes in the miRNA expression profile
Alterations in miRNAs have been associated with aging and cognitive decline (Cosín-Tomás et al.,2018).Moreover,specific miRNΑ abnormal levels have been detected in several areas of the AD brain (Wang et al.,2019).Thus,to gain insight into the biology of G9a inhibition treatment,we analyzed the global miRNA expression between SAMP8 control and SAMP8 treated mice with UNC0642 in the whole brain.We identified 44 differentially expressed miRNΑs (fold change cutoff of ≥1.9,P-value <0.05),of which 27 were up-regulated and 17 were down-regulated in UNC0642 treatment (Figure 1A).The top 10 up-and down-regulated miRNAs are shown inFigure 1B.Then,we validated the increase in the expression ofmmu-miR-1298-5p(P=0.0061) and themmu-miR-669c-5p(P=0.0717) and the reduction of themmu-miR-677-5p(P=0.0211) andmmu-miR-128-3p(P=0.0213) by qRT-PCR (Figure 1F–I).These results suggest that UNC0642 treatment induces an alteration in miRNΑ expression levels.
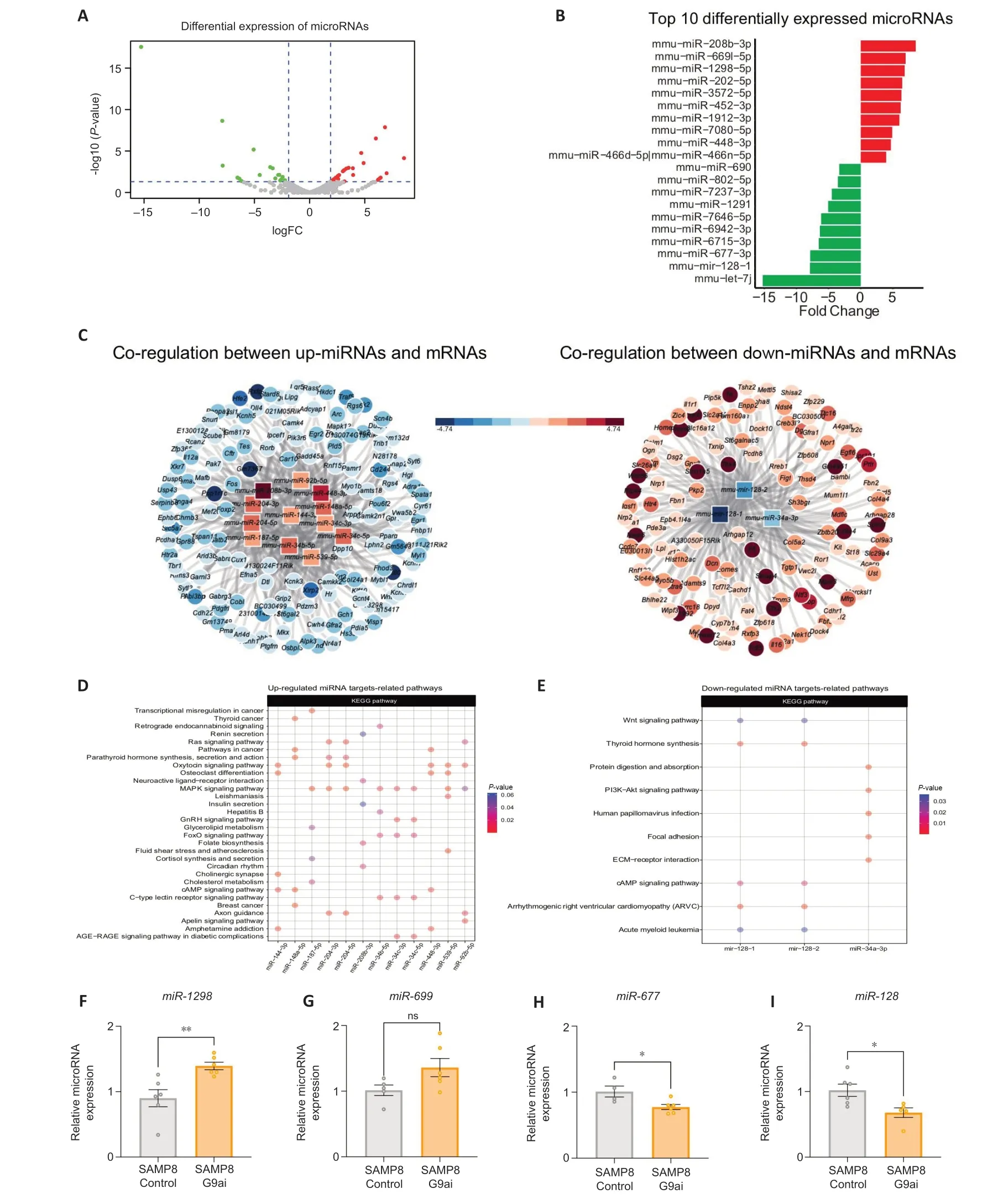
Figure 1|G9a inhibition treatment induces changes in the miRNome.
To determine co-regulation between miRNAs and mRNAs we used the miRTarVis tool (Jung et al.,2015).To determine alterations in mRNA expression,we used our previously reported RNA sequencing (GSE189249) in SAMP8 control and SAMP8-treated mice with UNC0642.We found several nodes with a high number of miRNAs-associated mRNA molecules in our SAMP8 model (Figure 1C).On one hand,the up-regulated miRNAs,mmu-miR-92b-5p,mmu-miR-144-3p,mmu-miR-34c-5p,mmu-miR-204-3p,mmu-miR-539-5p,mmu-miR-34c-3p,mmu-miR-204-5p,mmu-miR-208b-3p,mmu-miR-148a-5p,mmu-miR-34b-5p,mmu-miR-448-3p,andmmu-miR-187-5pare associated with the down-regulation of 145 mRNΑs (Figure 1DandAdditional Tables 5and6).On the other hand,the down-regulated miRNAs,mmu-miR-34a-3,mmu-mir-128-1,andmmu-mir-128-2are associated with the up-regulation of 124 mRNAs (Figure 1EandAdditional Tables 5and6)according to miRanda algorithm (Betel et al.,2008).Together,these data show that G9a inhibition with UNC0642 treatment causes a transcriptional dysregulation of miRNΑs,which could alter biological processes in the SAMP8 model.
Downregulation of miR-128-3p after a G9a inhibition partially explains the modulation of oxidative stress responses and autophagy pathway by targeting Pparg
To explore the mechanism underlying the neuroprotective effects of G9ai treatment,the potential mRNAs targeted bymiR-128-3pwere examined for being the second most downregulated miRNA.MiRNAs require complementary binding with 6 consecutive nucleotides in the 3′ UTR of a target mRNΑ (Αgarwal et al.,2015).Based on Target Scan Human 8.0 (Lewis et al.,2005;Grimson et al.,2007;Friedman et al.,2009;Garcia et al.,2011;Αgarwal et al.,2015;McGeary et al.,2019),the 3′-untranslated region (3′-UTR) of PPΑRG mRNΑ contains a putative 7-ntmiR-128target sequence (Figure 2A).In accordance,higherPparggene expression levels were statistically significant in the SΑMP8 group treated with G9ai(P=0.0001;Figure 2B).

Figure 2|Downregulation of miR-128 after a G9a inhibition partially explains the modulation of treatment modulates oxidative stress responses by targeting peroxisomeproliferator activator receptor γ (Pparγ).
Moreover,gene expression of potential PPΑRγ target genes was evaluated.First,it was observed that the expression of antioxidant genes,such as superoxide dismutase-1 (Sod1),heme oxygenase-1 (Hmox1),glutathione peroxidase 1 (Gpx),and aldehyde oxidase-1 (Aox1),was significantly up-regulated after treatment with the G9ai (P=0.0187,0.0009,0.0035,0.0288,respectively;Figure 2C).Accordingly,the gene expression levels of cyclooxygenase-2 (Cox2) and aldehyde dehydrogenase-2 (Aldh2),both pro-oxidant genes,were significantly downregulated in the treated SAMP8 group(P=0.0476,0.0090,respectively;Figure 2D).Second,we found an increase in the protein levels of FOXO1a in SAMP8 treated with UNC0642 compared with the SAMP8 control group (Figure 2FandH).Likewise,we observed a statistically significant increase in the p-ΑMPK/ΑMPK ratio in the SΑMP8-treated group (P=0.0370;Figure 2FandG).We also found increased gene expression of deacetylase sirtuin 1 (Sirt-1),which regulates the activity of FOXO transcription factors (P=0.0493;Figure 2E),in treated SAMP8 with UNC0642.Then,we assessed the protein levels of antioxidant enzymes.Catalase-3(CΑT3),GPX,and SOD1 were upregulated in the SΑMP8 group treated with G9ai compared with the control group (P=0.02,0.0203,respectively;Figure 2I–L).
To further confirm the antioxidant effects of the G9a inhibition,we treated primary neuron cell cultures with H2O2as an oxidative insult.In pathological conditions (H2O2),we observed a significant increase in cell survival in the group treated with the G9aiversusthe one treated only with vehicle (P=0.0003;Figure 2M).Then,the antioxidant enzyme glutathione synthetase was upregulated after the pharmacological inhibition of G9a (P=0.0021;Figure 2N–O).
Αutophagy can be regulated by the PPΑRγ/ΑMPK (Wang et al.,2022).Therefore,we evaluated several autophagy markers.Strikingly,sequestosome-1 (SQSTM1/p62) and Beclin-1 were found upregulated in the SAMP8 treated with the G9ai in comparison with the control (P=0.0434,0.0118,respectively;Figure 3A–C),suggesting autophagy activation.
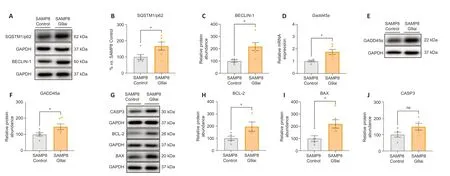
Figure 3|Upregulation of peroxisome-proliferator activator receptor γ (PPARγ) by G9ai enhances the expression of genes involved in DNA damage responses,apoptosis,and autophagy.
Upregulation of PPARγ by G9ai enhances the expression of genes involved in DNA damage response and apoptosis
The growth arrest and DNΑ damage 45 alpha (GΑDD45α)plays an important role in DNA repair (Smith et al.,2000),cell cycle (Vairapandi et al.,2002),apoptosis (Tront et al.,2006),angiogenesis,senescence (Tront et al.,2006),and DNA demethylation (Lee et al.,2012) by recruiting demethylation proteins to CpG island promoters (Αrab et al.,2019).Consistent with this,we observed an increase in bothGadd45agene expression and protein levels of GΑDD45α after the pharmacological inhibition of G9a in SAMP8 mice(P=0.0119,0.0408,respectively;Figure 3D–FandAdditional Table 6).Αs we mentioned before,GΑDD45α plays an important role in apoptosis (Hildesheim et al.,2002;Tront et al.,2006;Salvador et al.,2013;Kleinsimon et al.,2018).Accordingly,three well-established apoptotic markers,B-cell lymphoma 2 (BCL-2),Bcl-2 Associated X-protein (BAX),and Caspase-3 (CΑSP3),were significantly upregulated in the treated SAMP8 group compared with the control group (P=0.042,0.0223,0.0964,respectively;Figure 3G–J).
PPARγ/GADD45α contributes to the enhancement of synaptic plasticity and neurogenesis after G9a inhibition
Lack ofGadd45apromotes alterations in hippocampal memory and long-term potentiation,a phenotype accompanied by reduced levels of memory-related mRNA (Aparisi Rey et al.,2019).Then,we evaluated gene expression levels of immediate early genes (IEGs) after G9ai pharmacological treatment in SAMP8 mice,since they have a relevant role in the regulation of neuronal activity,synaptic plasticity,and neurogenesis (Minatohara et al.,2016;Duclot and Kabbaj,2017;Kim et al.,2018;Mahringer et al.,2019;Meenakshi et al.,2021).Early growth response 1 (Egr1),activity regulated cytoskeleton associated protein (Arc),andDnmt3agene expression levels were found significantly increased in the treated SAMP8 group in comparison with the control SAMP8 group (P=0.0231,0.0013,0.0428,respectively;Figure 4A).However,no statistical differences were observed inEgr2gene expression levels (P=0.4063;Figure 4A).Synaptic plasticity-related gene expression,such as VGF nerve growth factor inducible (Vgf) and transforming growth factor (Tgf) were found upregulated after G9ai pharmacological treatment in SAMP8,although the increase was only statistically significant forVgf(P=0.0007;Figure 4B).Likewise,the gene expression of the neurotrophin receptors,TrkAandTrkB,were upregulated in the SAMP8 treated group (P=0.001,0.0337,respectively;Figure 4B).Then,we quantified the levels of well-established synaptic plasticity proteins.In the case of postsynaptic density protein 95 (PSD95),we observed significantly increased levels in the SAMP8 mice treated with a G9ai compared with the control group (P=0.0407;Figure 4CandD).In contrast,in the case of PSD93 and synapsin IIa (SYN Iia),a positive tendency was observed after pharmacological treatment (Figure 4C,E,andF).In terms of neurogenesis markers,doublecortin (Dcx) gene expression levels were increased in the SAMP8-treated group compared with the control groups (P=0.0001);however,no differences were observed in SRY-Box transcription factor 2 (Sox2) gene expression levels between both experimental groups (Figure 4GandH).Finally,protein levels of neuronal nuclei (NeuN)were increased in the SAMP8 treated with G9aiversusthe Control SAMP8 group (P=0.0165;Figure 4IandJ).
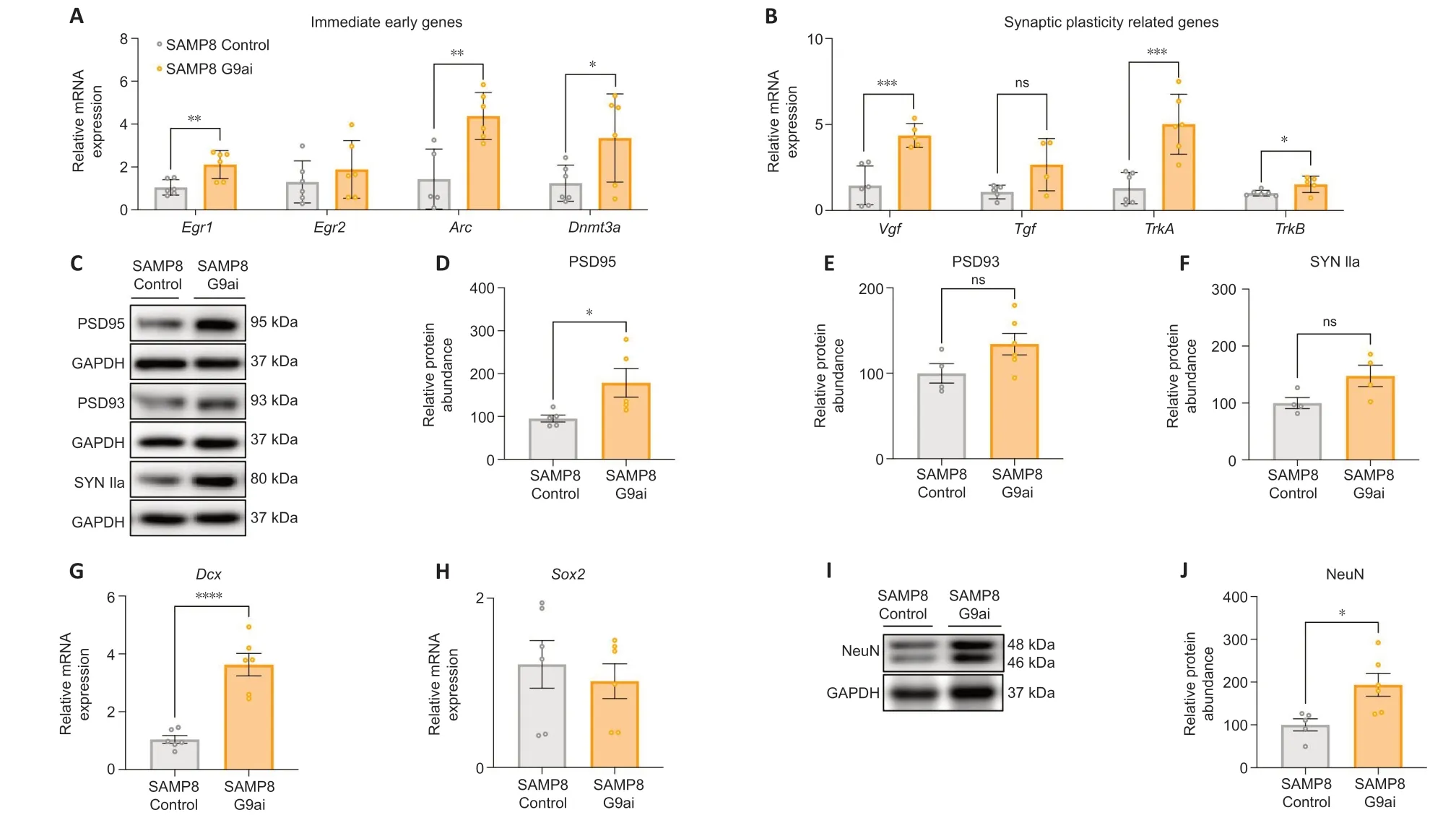
Figure 4|PPARγ/GADD45α might contribute to the enhancement of synaptic plasticity and neurogenesis after G9a inhibition.
Discussion
In this study,we have identified and explored an unknown molecular pathway based on PPΑRγ/GΑDD45α transcriptional repression controlled by G9a which also modulates miRNAs.Several studies suggest that G9a is a contributing factor to neuroinflammation and is upregulated in AD patients(Zheng et al.,2019;Bellver-Sanchis et al.,2023).In addition,a recent study stated that G9a is necessary to produce proinflammatory cytokines and reactive oxygen species,which are associated with OS (Mourits et al.,2021).It has been suggested that G9a is also necessary for the maintenance of epigenetic marks since it can promote or repress gene expression,and thereby affects the expression of several miRNΑs (Shankar et al.,2013).Thus,there is evidence of the crosstalk between G9a,miRNΑs,OS,and neuroinflammation and its contribution to the pathogenesis of ΑD.
Specific miRNΑs participate in the initiation and progression of neurodegenerative diseases,including ΑD (Kumar and Reddy,2020).Indeed,several miRNAs play a role in the control of key components of neuronal functions such as learning and memory formation (Saab and Mansuy,2014).Furthermore,as aforementioned,miRNAs regulate gene expression at the post-transcriptional level (Gurtan and Sharp,2013).Αt first,by overlaying transcriptomic analyses,we identified several miRNAs (miR-1298,miR-669,miR-677,andmiR-128) that were modified.The KEGG enrichment analysis revealed the association of G9a with cholinergic synapse,axon guidance,and cAMP signaling pathway,among others.But what is more noteworthy is that KEGG enrichment analysis showed that G9ai treatment modulates the MAPK and FOXO signaling pathways,which is further confirmed throughout the work.An example of one of the modulated miRNAs wasmiR-1298,which is downregulated in different pathological conditions and is associated with hypermethylation of its upstream DNΑ CpG sites (Hu et al.,2015).Αccordingly,our results showed a downregulation ofmiR-1298in SAMP8 mice,which was reverted after G9ai,indicating a reduction in DNΑ methylation levels,allowing the upregulation of this miRNA and thus,modifying the gene transcription of different important neuroprotective genes.We also found an upregulation inmiR-669after G9a inhibition with UNC0642 in SΑMP8 mice.These results are in agreement with recent work from Kolosowska et al.(2020),which showed that the upregulation ofmiR-669c-3ppromotes neuroprotective effects in a mouse model of ischemic stroke through induction of the expression of antiinflammatory microglial markers and PPΑRγ.The latter is the one we have focused on in this study.Moreover,we found downregulation inmiR-677andmiR-128after G9a inhibition.miR-677has a role in protein synthesis directly associated with neurodegenerative disease,including AD,such as protein related to neuroactive ligand-receptor,autophagy,and Wnt signaling pathway as we found in SΑMP8 after G9ai.Last but not least,miR-128is involved in the development of the nervous system as well as the regulation of neuronal plasticity,neuronal death,and cognitive impairment (Hu et al.,2015;Kolosowska et al.,2020).Importantly,recently published research has shown thatmiR-128is increased in the hippocampus of AD human brains (Siedlecki-Wullich et al.,2021).More importantly,miR-128downregulation improved cognitive deficits and reduced Α pathology in 3×Tg-ΑD mice (Liu et al.,2019).Furthermore,this improvement in cognition was associated with increased levels of PPΑRγ.PPΑRγ is targeted bymiR-128(Liu et al.,2019).In our case,we found reduced levels ofmiR-128in SAMP8-treated mice with UNC0642,which were correlated with better cognitive performance,as we demonstrated in our previously published manuscript(Bellver-Sanchis et al.,2023),suggesting the regulation ofmiR-128by G9a.PPΑRγ is an essential nuclear receptor involved in energy homeostasis and inflammation,which promotes growth interconnection of various pathways,such as the nuclear factor erythroid 2 related factor 2 (NRF2),Wnt/β-catenin,and FOXO (Polvani et al.,2012).Also,several studies have shown that PPΑRγ is implicated in the OS response,which involves an imbalance between pro-oxidation and anti-oxidation forces that may lead to cell death (Polvani et al.,2012;Vallée and Lecarpentier,2018;Beheshti et al.,2022).In this regard,we evaluated gene expression related to this molecular pathway.On one hand,the expression of the antioxidant genes evaluated in this work was increased after treatment with G9ai,both forin vitroandin vivostudies.In the same way,the expression of the pro-oxidant markers was downregulated in the treated SΑMP8 mice.It has been observed that PPΑRγ agonists protected cells from oxidative stress after ischemia by suppressing COX2 expression in a PPΑRγ-dependent manner (Zhao et al.,2006).On the other hand,one of the more important upregulated signaling pathways in SAMP8-treated mice was FOXO.In accordance with our results,it has been reported that G9a protein levels were elevated and FOXO1 protein levels were decreased in human colon cancer patients (Chae et al.,2019).Hence,to our knowledge,herein we described for the first time the upregulation of the FOXO signaling pathway in the SAMP8 brain after treatment with UNC0642.Interestingly,in the FOXO1-dependent mechanism,AMPK controls the expression of antioxidant enzymes,and therefore this enzyme may have an important role in the maintenance of redox homeostasis (Yun et al.,2014).Accordingly,the increased expression of some of the OS markers evaluated in this work,such as CΑT3 or SOD1(Ding et al.,2007;Yu et al.,2008),is directly related to all PPAR isoforms (Muzio et al.,2021).For example,increased PPΑRγ has been shown to correlate with increased levels of glutathione synthetase (Ferguson et al.,2009),and HMOX1(Krönke et al.,2007;Bilban et al.,2009;Ferguson et al.,2009;Polvani et al.,2012).Altogether,these data suggest the activation of molecular pathways that can revert the neuronal damage presented in AD.
In the same manner,the PPΑRγ/ΑMPK axis also regulates autophagy (Wang et al.,2022).It is well-known that G9a represses the expression of genes involved in the autophagic process (Αrtal-Martinez de Narvajas et al.,2013),and its inhibition promotes the activation of the autophagic pathway(Griñán-Ferré et al.,2019).Αccording to this,here we found an induction of autophagy markers (BECLIN-1,SQSTM1/p62,among others) that are associated with the completion of the beneficial effects of the autophagic process after pharmacological inhibition of G9a in SΑMP8-treated animals.PPΑRγ is found to be correlated with GΑDD45α protein(Fujiki et al.,2009).GΑDD45α protein interacts with PPΑRγ,recruiting demethylation proteins to CpG island promoters,and consequently upregulating its transcriptional activity (Αrab et al.,2019),and consequently upregulating transcriptional activity of PPΑRγ (Fujiki et al.,2009).Indeed,it has been described thatGadd45is widely expressed in the mouse brain,including the cerebral cortex and hippocampus,both fundamental regions for memory formation (Matsunaga et al.,2015;Αparisi Rey et al.,2019).Recently,it was shown thatGadd45a-deficient mice displayed alterations in hippocampal memory and long-term potentiation,associated with reduced mRNA levels of genes related to memory (Leach et al.,2012).Thus,we evaluated the gene expression levels of IEGs after the pharmacological treatment of SAMP8 mice and found changes inEgr1andDnmt3agene expression,suggesting the improvement in cognition previously demonstrated(Vasilopoulou et al.,2022).Furthermore,these results were accompanied by increased gene expression of neurotrophin receptors,TrkAandTrkB.Then,we proceed to evaluate the levels of well-established synaptic plasticity proteins.In the case of the protein levels of PSD95,PSD93,and SYN IIa,as well as neurogenesis markers,Dcxgene expression levels as well as protein levels of NeuN were increased in the SAMP8 treated with G9ai versus the Control SAMP8 group.Those results were consistent with previous published results by us and others,that showed that elevated H3K9me2 and G9a levels are present in the hippocampus of several AD mice models,promoting diminished synaptic transmission,and learning and memory deficits (Griñán-Ferré et al.,2019;Zheng et al.,2019).Those processes were ameliorated by G9a inhibition with UNC0642 or BIX01294 treatment,pointing to the participation in synaptic dysfunction and cognitive decline of G9a.However,the exact mechanisms by which G9a inhibition promotes miRNA changes remain unclear and require further studies.
The main limitation of this study could be to describe in depth the direct modulation ofmiR-128/Ppargby pharmacological inhibition of G9a.Further studies with gene editing or also using a pharmacological approach would be helpful to confirm this.These experiments could provide more robustness to our results,confirming the direct link between G9a andmiR-128/Pparg.However,it has been noted that all the evidence already cited throughout the manuscript agrees with our findings,as well as we report the changes in the miRNome profile after pharmacological treatment of G9a in a wellestablished AD mice model,the SAMP8.
In conclusion,this study demonstrates that G9a inhibition with UNC0642 improves the gene expression profile of mRNΑs and miRNAs in the SAMP8 mouse model.The differentially expressed miRNAs after UNC0642 treatment promote the activation of the PPΑRγ/GΑDD45α axis,leading to changes in crucial molecular pathways associated with age-related cognitive decline.Therefore,these results suggest that G9a and miRNΑs are promising diagnostic biomarkers and therapeutic targets for neurodegenerative diseases such as ΑD.
Author contributions:CGF conceived the study.ABS,CGF,PAAL,IT,MRV,and DVG performed the experiments.ABS,CGF,AG,PAAL,IT,and CP designed the experiments and analyzed the data.ABS,DB,LL,GC,AG,MP,and CGF wrote the manuscript.All authors read and approved the final manuscript.
Conflicts of interest:None of the authors has any disclosures to declare.
Author statement:This paper has been partially posted as a preprint on Research Square with doi:https://doi.org/10.21203/rs.3.rs-1228952/v1,which is available from:https://assets.researchsquare.com/files/rs-1228952/v1_covered.pdf?c=1641408565.
Data availability statement:All relevant data are within the paper and its Additional files.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
Additional files:
Additional Table 1:Reagents and kits used in this study.
Additional Table 2:TaqMan probes used in this study.
Additional Table 3:SYBR Green primers analyzed in this study.
Additional Table 4:Antibodies used in western blot assay in this study.
Additional Table 5:Differentially expressed miRNAs.
Additional Table 6:miRNAs targets.

- 中国神经再生研究(英文版)的其它文章
- A sphingolipid message promotes neuronal health across generations
- Krüppel-like factor 2 (KLF2),a potential target for neuroregeneration
- Defined hydrogels for spinal cord organoids: challenges and potential applications
- Neuronal trafficking as a key to functional recovery in immunemediated neuropathies
- Advancements in personalized stem cell models for aging-related neurodegenerative disorders
- New insights into astrocyte diversity from the lens of transcriptional regulation and their implications for neurodegenerative disease treatments