
lncRNA KCNQ1OT1靶向调控miR-132-5p对帕金森病细胞损伤的保护机制
2024-04-07 02:18付琨燕刘慧斌苏男李斯琴
河北医药 2024年5期
付琨燕 刘慧斌 苏男 李斯琴
帕金森病是一种黑质多巴胺随年龄增长而呈退行性变的疾病,故发病人群以>60岁为主,目前尚未有根治手段,研究该病的发生、发展作用机制,对其治疗具有重要意义[1-3]。长链非编码RNA(lncRNA)是可在转录后调节基因表达的非编码RNA分子,可作为帕金森治疗的潜在靶点,但lncRNA KCNQ1OT1可在转录后调节基因表达而参与肿瘤、糖尿病肾病及心肌病的发生、发展,但在帕金森中相关作用机制目前尚不明确[4]。miRs在神经细胞凋亡、氧化应激及炎性反应过程中均发挥重要作用,miR-132在帕金森病中呈高表达,可参与应激反应调节[5]。1-甲基-4-苯基吡啶阳离子(MMP+)是一种在帕金森病细胞实验模型建立中应用较为广泛的神经毒素,可造成多巴胺能神经元氧化应激、细胞凋亡[6]。笔者发现lncRNA KCNQ1OT1、miR-132-5p在帕金森病模型细胞中的作用和关系尚不明确,基于此,本研究通过MMP+诱导建立帕金森病模型细胞,以下调lncRNA KCNQ1OT1、miR-132-5p表达的方法明确其对帕金森病模型细胞损伤的影响。
1 材料与方法
1.1 研究材料 (1)研究细胞:人神经母细胞瘤株SH-SY5Y细胞来自中国科学院细胞库,本研究经巴彦淖尔布医院伦理委员会审核批准。(2)主要试剂:RPMI-1640培养基(武汉益普生物科技有限公司),活性氧(ROS)、超氧化物歧化酶(SOD)、丙二醛(MDA)、IL-1β、IL-6、肿瘤坏死因子-α(TNF-α)检测试剂盒(北京索宝科技有限公司),lncRNA KCNQ1OT1抑制剂(anti-lncRNA KCNQ1OT1)、模拟物(lncRNA KCNQ1OT1)、miR-132-5p抑制剂(anti-miR-132-5p)、模拟物(miR-132-5p)、阴性对照(KCNQ1OT1-NC、miR-NC、si-lncRNA KCNQ1OT1、pcDNA)、miR-132-5p的野生及突变型双荧光素酶载体均购自南通百奥迈科生物公司,流式细胞仪(美国BD公司),BCA蛋白定量试剂盒(上海碧云天生物技术公司),Bcl-2蛋白、Bax蛋白抗体(SCBT公司)。
1.2 方法
1.2.1 细胞培养:将人神经母细胞瘤株SH-SY5Y细胞放入RPMI-1640培养液(10%胎牛血清、100 U/mL青霉素、100 μg/ml酶联素)中,之后将其放入培养箱(温度37℃、5%CO2)中进行培养,每3天更换1次培养液,密度达80%~90%时,将培养基除去,进行传代培养,取3~5代细胞进行后续实验。
1.2.2 模型建立:取对数生长期SH-SY5Y细胞分为Con组(正常培养),MMP+组(将浓度为100 μmol/L的MMP+加入培养液中,诱导24 h进行转染)。
1.2.3 细胞转染:取对数MMP+培养SH-SY5Y细胞,稀释至1×106个细胞/mL,后接种于6孔板上,200 μL/孔(1×106个/孔),待细胞融合度达80%,以Lipofectamine 2000进行转染,分组:lncRNA KCNQ1OT1抑制组(转染anti-KCNQ1OT1-NC和anti-lncRNA KCNQ1OT1),miR-132-5p抑制组(转染anti-miR-132-5p-NC和anti-miR-132-5p),lncRNA KCNQ1OT1过表达组(转染KCNQ1OT1-NC+lncRNA KCNQ1OT1),lncRNA KCNQ1OT1低表达和miR-132-5p过表达(转染anti-KCNQ1OT1+pcDAN和anti-KCNQ1OT1+pcDAN+miR-132-5p),野生型和突变型双荧光素酶报告系统(转染KCNQ1OT1-NC+WT-miR-132-5p、lncRNA KCNQ1OT1+WT-miR-132-5p、KCNQ1OT1-NC+MUT-miR-132-5p、lncRNA KCNQ1OT1+MUT-miR-132-5p),转染48 h后进行后续实验。
1.2.4 lncRNA KCNQ1OT1、miR-132-5p表达量检测:SH-SY5Y细胞lncRNA KCNQ1OT1、miR-132-5p表达量采用实时荧光定量PCR(RT-PCR)法检测,采用TRIzol试剂盒提取和纯化RNA,使用Takara逆转录试剂盒进行逆转录处理,获得总cDNA,采用Primer 5.0软件设计引物序列,启动PCR仪,反应条件:95℃、94℃、59℃、72℃分别为5 min、30 s、45 s、45 s,45个循环,采用2-ΔΔCt方法计算出lncRNA NEAT1表达量。
1.2.5 ROS、SOD、MDA、IL-1β、IL-6、TNF-α水平检测:取对数生长SH-SY5Y细胞,接种于6孔板,孵育过后通过酶标仪对ROS、SOD、MDA、IL-1β、IL-6、TNF-α水平进行检测,孔中加入10 μL待测样品、40 μL待测样品稀释液,摇匀,加入酶标试剂100 μL,封板并孵育60 min(37℃),将孔中液体弃去,Wash Sohltion重复清洗后加入50 μL显色剂,摇匀,显色处理15 min(37℃),后加入终止液50 μL,OD值在450 nm波长下检测ROS、SOD、MDA、IL-1β、IL-6、TNF-α水平。
1.2.6 细胞凋亡率检测:取对数生长SH-SY5Y细胞,接种于6孔板上,2×104个/孔,培养48 h后将培养液弃去,PBS进行洗涤,共2次,后将结合缓冲液重悬细胞(200 μL)、AnnexinV-异硫氰酸荧光素(5 μL)、PI(5 μL)加入,结合缓冲液在孵育15 min后加入,共300 μL,后通过流式细胞仪对细胞凋亡率进行检测。
1.2.7 Bcl-2、Bax蛋白表达检测:细胞中蛋白依照细胞蛋白提取试剂盒说明进行萃取,Bcl-2、Bax蛋白表达量通过Western blot法进行检测,SDS-PDGE电泳于每孔加入80 μg蛋白后进行,电泳结束后将蛋白电转至PVDF膜上,5%脱脂奶粉对PVDF膜进行封闭,Bcl-2、Bax蛋白一抗(1∶1 000)于摇床上室温孵育,120 min,完成后采用TBST洗膜3次,5 min/次,后将Bcl-2、Bax蛋白K二抗(1∶1 000)于摇床上室温孵育,60 min,完成后采用TBST洗膜3次,10 min/次,化学发光显影采用ECL,以GAPDH为内参,相对条带灰度光密度值采用Image J软件分析。
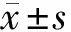
2 结果
2.1 lncRNA KCNQ1OT1和miR-132-5p在MMP+诱导SK-N-SH细胞损伤中的表达 与Con组相比,MMP+组lncRNA KCNQ1OT1、miR-132-5p表达量较高,差异均有统计学意义(P<0.05)。见表1。
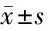
表1 lncRNA KCNQ1OT1和miR-132-5p在MMP+诱导SK-N-SH细胞损伤中的表达
2.2 lncRNA KCNQ1OT1低表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响 与Con组比较,MMP+组lncRNA KCNQ1OT1、ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均升高,SOD水平、Bcl-2蛋白表达均降低,差异均有统计学意义(P<0.05);与MMP++KCNQ1OT1-NC组比较,MMP++si-lncRNA KCNQ1OT1组lncRNA KCNQ1OT1、ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均降低,SOD水平、Bcl-2蛋白表达均升高,差异均有统计学意义(P<0.05)。见表2,图1。

图1 Western blot检测凋亡相关蛋白表达
表2 lncRNA KCNQ1OT1低表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响
2.3 干扰miR-132-5p表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响 与Con组比较,MMP+组miR-132-5p表达量、ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均升高,SOD水平、Bcl-2蛋白表达均降低,差异均有统计学意义(P<0.05);与MMP++miR-NC组相比,MMP++si-miR-132-5p组miR-132-5p表达量、ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均下降,SOD水平、Bcl-2蛋白表达均上升,差异均有统计学意义(P<0.05)。见图2,表3。

图2 Western blot检测凋亡相关蛋白表达
表3 干扰miR-132-5p表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响
2.4 双荧光素酶报告实验 双荧光素酶报告系统结果显示,与KCNQ1OT-NC组比较,过表达lncRNA KCNQ1OT组野生型WT-miR-132-5p的荧光从荧光素酶相对活性较高,差异有统计学意义(t=6.708,P<0.05),而突变型MUT-miR-132-5p无显著变化,差异无统计学意义(P>0.05)。见表4。
表4 双荧光素酶报告实验
2.5 miR-132-5p过表达逆转了lncRNA KCNQ1OT1低表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响 与MMP++KCNQ1OT1-NC组比较,MMP++si-lncRNA KCNQ1OT1组miR-132-5p表达量、
ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均降低,SOD水平、Bcl-2蛋白表达均增加,差异有统计学意义(P<0.05);与MMP++si-lncRNA KCNQ1OT1+pcDNA组比较,MMP++si-lncRNA KCNQ1OT1+pcDNA+miR-132-5p组miR-132-5p表达量、ROS、MDA、IL-1β、IL-6、TNF-α水平、凋亡率、Bax蛋白表达均上升,SOD水平、Bcl-2蛋白表达均下降,差异有统计学意义(P<0.05)。见表5,图3。

图3 Western blot检测凋亡相关蛋白表达
表5 miR-132-5p过表达逆转了lncRNA KCNQ1OT1低表达对MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡的影响
3 讨论
帕金森病是发病隐秘、病程进展缓慢的老年疾病,临床症状多表现为肢体震颤、活动期间姿态笨拙,且部分患者可伴随步态障碍或运动缓慢等症状,进而对患者正常机体活动造成了不利影响[7-9]。有学者发现神经元氧化应激及炎性损伤、细胞凋亡为帕金森病发病的主要原因,因此寻找可有效抑制神经元氧化应激及炎性损伤、细胞凋亡的分子靶点对临床诊治具有关键意义[10-12]。
研究有明,帕金森病病理机制中lncRNA具有重要的调控作用,如lncRNA NEAT1可通过调控RAB3IP、miR-212-5p表达而减轻MMP+引发的细胞毒性及凋亡,lncRNA UCA1可通过抑制P13K/AKT激活而减轻氧化应激炎性反应,lncRNA H19可通过激活Wnt/β-catenin、调控HPRT1表达而减轻神经元细胞损伤,lncRNA KCNQ1OT1作为新型lncRNA可参与内脂应激、神经元损伤及凋亡调节,在脑缺血再灌注损伤、缺血性脑卒中、神经胶质瘤等脑部相关疾病中均发挥重要作用[13-15]。近年来研究表明,lncRNA可通过调控miRNA表达而对下游效应分子功能进行调节,而miRNA可通过靶向调控蛋白表达、抑制或靶向降解其靶mRNA而参与人类疾病的发生、发展,miR-132在帕金森病中呈高表达,且与患者疾病严重程度、疾病分期密切相关,而抑制miR-132表达可减少神经细胞凋亡,增加细胞存活量[16]。本研究显示,miR-132-5p在MMP+诱导SK-N-SH细胞中呈高表达,双荧光素酶实验结果显示,miR-132-5p为lncRNA KCNQ1OT1的靶基因,lncRNA KCNQ1OT1可正向调控miR-132-5p表达,表明lncRNA KCNQ1OT1可靶向调控miR-132-5p表达。
氧化应激反应易对大脑造成影响,故临床多认为神经系统疾病的发病与氧化应激反应存在密切联系[17]。ROS为线粒体产物,其低水平表达与正常细胞功能维持存在密切联系,当抗氧化防御系统与ROS平衡遭到破坏时,可引发氧化应激反应,继而推动了帕金森病的发生、发展[18-19]。SOD作为抗氧化酶,具有清除自由基的作用,其水平过低可造成ROS在机体内累及,进而促进了酮基、MDA等降解产物的产生,因此MDA可对细胞损伤程度进行间接反映,同时过表达的MDA可对线粒体膜造成损伤,进而对其流动性、通透性造成影响,使得线粒体内Cyt C释放进入细胞质,最终造成细胞凋亡[20-21]。本研究表明,低表达lncRNA KCNQ1OT1、干扰miR-132-5p表达均可减轻降低ROS、MDA水平,提升SOD水平,而lncRNA KCNQ1OT1低表达和miR-132-5p过表达可提升ROS、MDA水平,降低SOD表达,表明低表达lncRNA KCNQ1OT1、干扰miR-132-5p表达均可抑制细胞氧化应激损伤,而miR-132-5p过表达可逆转lncRNA KCNQ1OT1低表达产生的细胞氧化应激损伤保护。
帕金森病的主要病理生理标志之一为慢性神经炎症,慢性神经炎症可通过激活小胶质细胞核星形胶质细胞而加重多巴胺能神经元变性[22]。IL-1β、IL-6、TNF-α均为具有促炎作用的小胶质细胞活化M1表型,可通过激活细胞凋亡信号而造成器官损坏甚至衰竭,且可促进线粒体产生ROS并释放细胞色素,最终造成细胞凋亡[23]。细胞凋亡为涉及凋亡相关基因表达及产物相互影响、作用、一个基因级联反应的细胞自我消亡过程,Bcl-2、Bax蛋白均为细胞凋亡相关蛋白,分别具有抑制细胞凋亡、促进细胞凋亡的作用,两者表达量与神经细胞凋亡存在密切联系[24-25]。本研究表明,低表达lncRNA KCNQ1OT1、干扰miR-132-5p表达均可减轻降低IL-1β、IL-6、TNF-α水平、Bax表达,提升Bcl-2表达,而lncRNA KCNQ1OT1低表达和miR-132-5p过表达可提升IL-1β、IL-6、TNF-α水平、Bax表达,降低Bcl-2表达,表明低表达lncRNA KCNQ1OT1、干扰miR-132-5p表达均可抑制细胞炎性损伤及凋亡,而miR-132-5p过表达可逆转lncRNA KCNQ1OT1低表达产生的细胞保护。
本文通过转染lncRNA KCNQ1OT1抑制剂成功下调lncRNA KCNQ1OT1表达后发现MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡受抑,同时转染miR-132-5p干扰序列后上调miR-132-5p表达后发现lncRNA KCNQ1OT1低表达造成的细胞氧化应激、炎症损伤及凋亡抑制作用减弱,提示lncRNA KCNQ1OT1低表达可靶向miR-132-5p而抑制MMP+诱导SK-N-SH细胞氧化应激、炎症损伤及凋亡,发挥保护帕金森病细胞损伤的机制。说明lncRNA KCNQ1OT1、miR-132-5p是帕金森病的潜在分子靶点,可为帕金森病临床诊治提供指导。
猜你喜欢
保健医苑(2022年6期)2022-07-08
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
世界科学技术-中医药现代化(2020年2期)2020-07-25
山东医药(2020年9期)2020-05-20
活力(2019年22期)2019-03-16
西南军医(2016年6期)2016-01-23
医学研究杂志(2015年5期)2015-06-10
中国医药导报(2015年27期)2015-02-28
西南军医(2015年2期)2015-01-22
