
肉苁蓉总苷对HepG2肝癌荷瘤小鼠的影响
2024-04-08 02:41蒋勇军周士琦李静圆闫文杰
食品科学 2024年6期
冯 朵,王 靖,蒋勇军,周士琦,段 昊,李静圆,闫文杰,*
(1.北京联合大学生物化学工程学院,生物活性物质与功能食品北京市重点实验室,北京 100023;2.农业农村部食物与营养发展研究所,北京 100081;3.内蒙古三口生物科技有限公司,内蒙古 鄂尔多斯 017000)
肉苁蓉具有多种保健功能,具有明显的药用价值和经济价值[1]。肉苁蓉作为食药同源物质,在肝癌治疗中可以在早期预防、中晚期辅助治疗方面起到预防和治疗兼顾的作用。已有多项研究证明,肉苁蓉具有增强免疫力、抗肿瘤、抗氧化、延缓衰老等多种营养保健作用[2-6]。
研究表明肉苁蓉总苷(total glycosides,TG)具有保肝护肝、神经保护、抗氧化和改善记忆的作用。研究发现荒漠肉苁蓉TG可激活核因子E2相关因子2(nuclear factor NF-E2-related factor 2,Nrf2)/Kelch样ECH相关蛋白1(Kelch-like ECH-associated protein 1,Keap-1)通路,保护急性酒精性肝损伤[7]。此外,肉苁蓉TG对神经细胞缺血再灌注损伤具有一定的保护作用[8],表观改善大鼠认知功能[9-10]。还可与枸杞多糖配伍提高小鼠学习记忆能力,抑制脑内氧化应激水平[11]。
人体的生物屏障、免疫系统与庞大的肠道菌群之间形成一种微妙的和谐共生关系[12],始终处于动态、复杂的人体生态系统中[13]。肠道微生物群组成及其代谢物与肝癌的发生密切相关[14],之间存在复杂的相互作用[15]。肠-肝轴双向连接调节肠道微生物群的组成、肠上皮屏障和血管屏障的完整性,研究发现可以利用微生物治疗肝细胞癌[16]。其中,微生物组成和功能是了解肝癌进展的重要方面[17]。
调节肠道微生物是提高肝癌免疫治疗疗效、减少不良反应的很有前途的策略[14]。此外,调节肠道微生物还可重塑炎症微环境,肠道微生物群会影响肝脏炎症微环境,可为癌症预防和治疗开辟新思路[18]。Chen Wen等[15]在肠道菌群失衡的小鼠中可观察到胆固醇调节元件结合蛋白2(sterol-regulatory element binding protein 2,SREBP2)升高,SREBP2的药理学抑制可减弱肠道菌群失衡下小鼠肝癌的起始作用。Schneider等[18]研究发现肠道微生物群直接影响小鼠和人的肝脏炎症微环境,从而影响肝细胞癌的发展。随着对肝脏相关疾病的深入研究,专家学者们发现肠道菌群及其代谢物是肝癌发展中的重要参与者,因此从新视角深入了解肠道微生物在肝癌中的作用机制尤为重要[19]。
目前,肉苁蓉TG对肝癌小鼠的研究少有报道,因此本实验将HepG2肝癌细胞皮下注射到小鼠右后腿处,待细胞不断增殖成瘤,期间给小鼠灌胃不同剂量TG,观察药物对裸鼠体内肿瘤的抑制作用,从氧化应激、调节免疫、改善代谢和平衡肠道微生物层面深入探讨TG在裸鼠体内的抗肿瘤效果,以期为TG深度开发为抗肿瘤药物提供理论依据。
1 材料与方法
1.1 材料与试剂
Balb/c-nu裸鼠(雄性,体质号量18~22 g)(生产许可证号:SCXK(京)2019-0008)购自北京华阜康生物科技有限公司,于北京联合大学应用文理学院保健食品功能检测中心SPF级动物房(使用许可证号:SYXK(京)2023-0013)单笼饲养,保持12 h的明/暗循环,温度(26±1)℃,相对湿度(60±5)%,适应性喂养1 周后进行实验。
4%多聚甲醛、二甲苯、无水乙醇 国药集团化学试剂有限公司;过氧化氢酶(catalase,CAT)、谷草转氨酶(aspartate aminotransferase,GOT)、谷丙转氨酶(glutamic-pyruvic transaminase,GPT)和碱性磷酸酶(alkaline phosphatase,AKP)试剂盒 南京建成科技有限公司;AxyPrepDNA凝胶回收试剂盒 美国Axygen公司。
肉苁蓉TG(质量分数≥75%,以松果菊苷和毛蕊花糖苷合计)由内蒙古三口生物科技有限公司提供,用水溶解至相应质量浓度[20-21]。
1.2 仪器与设备
AB Triple TOF 6600质谱仪 美国AB SCIEX公司;290 Infinity LC超高效液相色谱系统、Bioanalyzer 2100生物分析仪 美国安捷伦科技公司;QuantiFluor™-ST蓝色荧光定量系统 美国Promega生物技术有限公司;ASP200S全自动脱水机、RM2235石蜡切片机、HI1220烤片台、DM3000正置荧光显微镜 德国Leica有限公司;INFINITE M NANO多功能酶标仪 瑞士Tecan公司;UV-1600紫外分光光度计 上海美谱达仪器有限公司。
1.3 方法
1.3.1 实验分组及皮下成瘤模型建立
HepG2肝癌细胞生长至培养瓶90%时进行传代,传代3 次后,使用胰酶消化,完全培养液终止消化细胞,收集细胞并计数,用磷酸盐缓冲液重悬至2×107个/mL,然后取0.1 mL细胞悬液用一次性注射器接种于裸鼠右后腿处皮下。接种HepG2肝癌细胞10 d后,将造模成功的动物随机分为4 组,分别为A组:模型组(生理盐水,n=9);B组:4 mg/kgmbTG(n=12);C组:16 mg/kgmbTG(n=11);D组:64 mg/kgmbTG(n=12)。连续灌胃18 d,期间每4~5 d使用游标卡尺测量肿瘤长度和宽度,按式(1)计算小鼠皮下实体瘤体积:
末次给药后,称小鼠体质量,眼球取血后脱颈处死小鼠,解剖剥离小鼠肝、脾和肿瘤组织。将剖离的肿瘤组织称质量后按式(2)计算抑瘤率,对各个组织称质量并按(3)计算脏器指数:
1.3.2 苏木精-伊红(hematoxylin-eosin,HE)染色观察组织病理变化
首先将肝脏和肿瘤组织使用多聚甲醛固定后进行石蜡包埋,切成4 μm厚的薄片。组织充分水化后用HE染色,脱水并风干切片,中性树胶封片后,于光学显微镜下观察各组织的病理变化并拍照。
1.3.3 肝癌小鼠脾脏淋巴细胞转化能力
以刀豆蛋白A(concanavalin A,con A)诱导动物脾脏淋巴细胞转化,利用四甲基偶氮唑盐法对淋巴细胞悬液进行测定,待细胞培养到72 h时,在570 nm波长处测定其光密度值(OD570nm)。通过式(4)计算淋巴细胞的增殖能力:
1.3.4 肝脏组织抗氧化指标检测
准确称取肝脏组织质量,按照质量体积比1∶9(g/mL)加入生理盐水,剪碎组织,冰水浴超声匀浆,离心后取上清液,放入4 ℃待测。待测样本准备好后,使用BCA法测定各样本的蛋白质量浓度,按照试剂盒说明书内容检测肝脏CAT、GOT、GPT、AKP活力。
1.3.5 肠道内容物16S rDNA扩增子测序与非靶代谢组学分析
收集肝癌小鼠肠道内容物于冻存管中,委托上海中科新生命生物科技有限公司完成。
1.3.5.1 16S rDNA 扩增子测序
采用十六烷基三甲基溴化铵或十二烷基硫酸钠方法对样本的基因组DNA进行提取,并对DNA的纯度和浓度进行检测。根据测序区域的选择,使用带Barcode的特异引物和高保真DNA聚合酶对选定的V3~V4可变区进行聚合酶链式反应(polymerase chain reaction,PCR)扩增。PCR产物用2%琼脂糖凝胶电泳进行检测,并对目标片段进行切胶回收,胶回收采用AxyPrepDNA凝胶回收试剂盒。参照电泳初步定量结果,对PCR扩增回收产物用QuantiFluorTM-ST蓝色荧光定量系统进行检测定量,按照每个样本的测序量要求,进行相应比例的混合。使用NEB Next®UltraTMDNA Library Prep Kit构建库试剂盒构建文库。构建好的文库通过Agilent Bioanalyzer 2100和Qubit进行质检,文库质检合格后进行上机测序。
获得原始数据之后,需要进行拼接、过滤得到有效数据。然后对有效数据进行相关的注释和分析,筛选组间差异显著的菌群(线型判别分析(linear discriminant analysis,LDA)值>2以及P<0.05)。
1.3.5.2 非靶代谢组学分析
提取样本,采用预冷的甲醇-乙腈-水溶液(2∶2∶1,V/V),涡旋充分混合,4 ℃、14 000×g离心20 min,取上清液备用。
采用超高效液相色谱系统HILIC色谱柱进行分离。柱温25 ℃;流速0.5 mL/min;进样量2 μL;流动相A为水+25 mmol/L乙酸铵+25 mmol/L氨水,B为乙腈;梯度洗脱程序如下:0~0.5 min,5% A、95% B;0.5~7 min,5%~35% A、95%~65% B;7~8 min,35%~60% A、65%~40% B;8~9 min,60% A、40% B;9~9.1 min,60%~5% A、40%~95% B;9.1~12 min,5% A、95% B。
样本通过色谱系统分离后进行质谱分析。电喷雾离子源:正、负离子模式进行检测:雾化气辅助加热气1(Gas1和Gas2)压力为60 psi。一级质荷比检测范围:60~1 000 Da,二级子离子质荷比检测范围:25~1 000 Da,一级质谱扫描累计时间:0.20 s/spectra,二级质谱扫描累计时间0.05 s/spectra。
获得原始数据之后,进一步对数据进行整理和定量分析,筛选组间差异显著的代谢物(变量权重值(variable importance for the projection,VIP)>1以及T检验(P<0.05),然后对筛选到的显著差异代谢物进行定性分析。
1.3.5.3 关联分析
基于Illumina双端测序平台,选择变异区域,利用保守区设计通用引物进行PCR扩增,对高变区进行操作分类单元(operational taxonomic units,OTU)聚类分析和菌种鉴定。利用β多样性进行组间分析,找出在不同分类水平上的分组间丰度变化差异显著的物种。
基于液相色谱-质谱联用技术,对生物样本中的小分子代谢物进行定性和相对定量分析。利用多维统计分析正交偏最小二乘判别分析(orthogonal partial least-squares discriminant analysis,OPLS-DA)的VIP值和单变量统计分析的P值,找出分组间丰度变化差异显著的代谢物。
利用Spearman统计方法分析实验样本中筛选到的显著差异菌群与显著差异代谢物之间的相关性系数,并结合R语言和Cytoscape软件,多角度挖掘菌群-代谢物之间的相互作用关系。
1.4 数据处理与分析
采用SPSS 25.0统计软件进行ANOVA单因素显著性分析,结果以表示;使用Graphpad Prism 8.0.2软件作图。P<0.05表示差异具有统计学意义,P<0.01表示具有极显著差异。
2 结果与分析
2.1 TG对HepG2肝癌小鼠体质量的影响
为了确定药物干预过程是否会影响裸鼠的正常生长,在饲喂期间对裸鼠的生长、饮食、活动能力进行跟踪观察,发现裸鼠在整个饲喂过程中,体质量正常生长、饮食正常,无毒副作用,这与Gao Yuqiu等[22]的研究结果一致。说明TG在抑制裸鼠实体瘤的同时,不会影响裸鼠的正常生长。
随着肿瘤的不断长大,在干预后期,模型组小鼠精神状态欠佳、行动缓慢、食欲下降、体型变瘦、活力下降、皮肤泛白,逐渐表现出恶病质症现象;其他处理组的裸鼠精神状态良好、皮肤红润、活动能力正常;在干预前期,TG各剂量组体质量生长速率较其他组更高(图1),由此推断TG可以给裸鼠提供一定的营养补充,但TG各组之间体质量没有差异。

图1 各组裸鼠平均体质量趋势变化Fig.1 Trends in average body mass of nude mice in each group
整体来看,裸鼠干预前期体质量增长快速,可能是由于肿瘤的快速增长促进了小鼠的摄食,导致体质量快速增长;中间增长趋势缓慢,可能是肿瘤的增长进入了平台期;而后期体质量的增长可能是由于肿瘤增长占据了主导地位。
2.2 TG对HepG2肝癌小鼠肿瘤体积的影响
为了观察药物是否可以抑制肿瘤的增长,每5 d测量一次裸鼠皮下实体瘤体积,通过表1发现,在干预前期,与模型组对比,处理组的增长趋势较为缓慢,当癌细胞快速繁殖,瘤体积不断扩大时,TG具有一定的抑制体内肿瘤体积的作用,且高质量浓度抑制效果更好((716.59±514.46)mm3)。

表1 TG对HepG2肝癌小鼠肿瘤体积的影响Table 1 Effect of TG on tumor volume of HepG2-bearing mice
2.3 TG对HepG2肝癌小鼠脏器指数、抑瘤率的影响
如表2所示,与模型组对比,经TG处理后的小鼠肝脏指数、脾脏指数均相应地降低,且具剂量-效应关系,但各组别之间均不存在显著差异(P>0.05)。各处理组的肿瘤质量均小于模型组,说明TG对HepG2荷瘤小鼠体内肿瘤均有一定程度的抑制作用,抑瘤率分别为6.133 2%、18.270 6%、26.855 7%,具有剂量依赖性。由于小鼠存在个体差异,处理组抑瘤率虽有所增长,但是各组之间并不存在显著差异(P>0.05)。

表2 TG对HepG2肝癌小鼠脏器指数、抑瘤率的影响Table 2 Effect of TG on visceral organ indices and tumor suppression rate of HepG2-bearng mice
2.4 TG对HepG2肝癌小鼠组织的影响
如图2a所示,模型组显示不规则的结构和颗粒细胞质,大多数肝细胞体积肿胀,细胞质疏松,细胞间隙扩大,部分肝细胞显示周围微小性症细胞浸润现象[23]。各处理组小鼠肝脏肝细胞结构较规则,坏死情况减少,炎性细胞浸润现象减轻,肝细胞多角或类圆形,呈清淡的嗜酸性着色,细胞核呈圆形或多角形,位于中央,染色质较稀疏,核膜清楚。

图2 TG作用于小鼠组织HE染色形态学变化Fig.2 Morphological changes of TG-treated HepG2-bearing mice by HE staining
肿瘤组织HE染色结果显示,模型组小鼠肿瘤细胞正常,生长旺盛,细胞核大,肿瘤细胞排列不规则,分布紧密,细胞结构完整(图2b)。各处理组肿瘤细胞分布变得越来越稀疏,细胞核出现固缩,空泡变样程度增加,细胞破裂乃至溶解坏死现象,结构不清,其中,高剂量TG组肿瘤细胞坏死效果最明显。
2.5 TG对HepG2肝癌小鼠脾脏淋巴细胞增殖能力的影响
从图3可知,与模型组对比发现随着TG质量浓度的提高,脾脏淋巴细胞增殖能力逐渐上升,分别增长了5.96%、29.57%、54.88%,呈剂量依赖性,但组间不具有显著差异(P>0.05)。该结果表明,TG可以通过改善小鼠脾脏淋巴细胞增殖能力从而达到抑制肝癌小鼠体内肿瘤增长的效果。
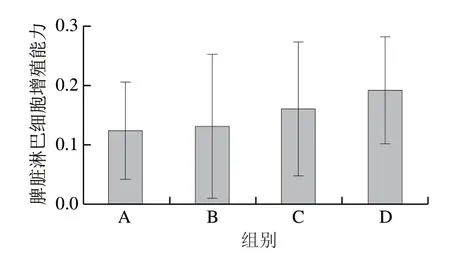
图3 TG作用于肝癌小鼠各组脾脏淋巴细胞增殖能力Fig.3 Effect of TG on proliferative capacity of splenic lymphocytes in HepG2-bearing mice
2.6 TG对HepG2肝癌小鼠肝脏组织抗氧化指标的影响
在癌变和免疫系统受损发生时,机体的抗氧化能力也会下降。由图4可知,与模型组比较,各处理组的肝脏组织CAT活力都有所上升,低、中、高剂量TG组分别提高了7.62%、18.51%、39.31%,提高幅度较大,各组与模型组之间均存在极显著差异(P<0.01)。与模型组对比,TG各剂量组(低、中、高)AKP活力分别下降了0.52%、3.76%、25.46%,其中,TG高剂量组与模型组存在极显著差异(P<0.01)。与模型组对比发现,随着TG质量浓度的提高,各TG剂量组GPT活力分别提高了35.70%、53.59%、62.46%。低、中、高TG剂量组小鼠肝组织GOT活力较模型组分别下降了2.21%、11.00%、11.58%。综上,推测模型组的肝脏发生了非常严重的病变,处理组较模型组病变程度低,并且随着TG浓度的升高,其病变程度随之降低。结果表明TG可以提高小鼠抗氧化能力、缓解小鼠肝脏氧化损伤程度、改善肝脏抗氧化功能,从而起到抑制小鼠体内肿瘤生长的作用,提示后续可采用肉苁蓉辅助治疗肝癌患者以保护肝脏器官免受损伤。

图4 TG对HepG2肝癌小鼠肝脏组织抗氧化指标的影响Fig.4 Effect of TG on antioxidant indexes of liver tissues in HepG2-bearing mice
2.7 肠道菌群代谢组学分析
由图5a可知,本实验正负离子模式合并后共鉴定得到1 889 种代谢物,其中正离子鉴定到1 163 种,负离子鉴定到726 种。对鉴定到的所有代谢物进行分类统计,按照化学分类归属信息主要包括有机酸及其衍生物、脂质和类脂分子、有机杂环化合物、苯环型化合物、有机氧化合物、苯丙烷类和聚酮类、有机氮化合物、核苷酸及其类似物等。

图5 肉苁蓉TG作用于小鼠肠道内容物代谢组学结果Fig.5 Metabolomics results of intestinal contents in TG-treated mice
OPLS-DA是一种对偏最小二乘判别分析(partial least squares-discrimination analysis,PLS-DA)进行修正的分析方法,可以滤除与分类信息无关的噪音,提高模型的解析能力和有效性;在OPLS-DA得分图中,有预测主成分和正交主成分,OPLS-DA将组间差异最大化的反映在t[1]上,可直接区分组间变异,而在正交主成分to[1]上则反映了组内的变异,椭圆代表95%置信区间,点的分布状态反映组间、组内的差异度。由图5b可知,OPLS-DA模型可以区分A组和D组。
显著差异代谢物(VIP>1,P<0.05)的层次聚类分析结果如图5c所示,聚在同一簇内的代谢物具有相似的表达模式,可能具有相似的功能或者共同参与同一代谢过程或细胞通路。图5c中每行代表一个差异代谢物,每列代表一组样品,红色代表显著上调,蓝色代表显著下调,颜色深浅表示上下调的程度,表达模式接近的代谢物聚在左侧同一聚类下。结果表明A组和D组代谢物明显不同,D组中茴香霉素、伊马替尼、拉贝洛尔、巨大戟醇、DL-正缬氨酸和原海葱苷A的含量较高,水杨酸含量较低,表明TG可显著调节肝癌小鼠肠道代谢物水平。
2.8 肠道菌群转录组学分析
群落结构组分图(图6a)可以展示每个样本或分组在不同分类水平上的群落结构,根据物种注释结果,选取A组和D组在属水平上最大丰度排名前10的物种,生成物种相对丰度柱形累加图,以便直观查看在属水平上相对丰度较高的物种及其比例。由图6a可知,D组中拟杆菌属(Bacteroides)较A组升高,罗氏菌属(Roseburia)、丹毒丝梭菌属(Erysipelatoclostridium)较A组降低。


图6 肉苁蓉TG作用于小鼠肠道内容物16S rDNA检测结果Fig.6 16S rDNA sequencing results of intestinal contents in TG-treated mice
微生物覆盖率为测序深度指数,均在0.99以上,品种间文库覆盖率指数通过Tukey和Wilcox秩和检验,如图6b所示,A组和D组存在显著差异(P=0.045<0.05),D组菌群多样性高于A组,表明测序深度已基本覆盖样品所有的物种,可进行下一步数据分析。
β多样性指数组间差异分析中,箱形图可以直观地反映组内物种多样性的中位数、离散程度、最大值、最小值、异常值。通过Wilcox秩和检验发现A和D组间物种多样性差异极显著(P=0.000 659 4<0.01),组间差异大于组内差异(图6c),表明分组具有意义。
图6d、e分别是不同处理组小鼠样本LDA值分布柱状图及具有统计学差异的Biomarker柱状图,代表在A组和D组起重要作用的微生物类群。重点关注两组中差异菌属的相对丰度,A组中卟啉单胞菌属(Porphyromonas)、鲁杰氏菌属(Ruegeria)和Tyzzerella_3菌属的丰度较高;D组中丁酸球菌属(Butyricicoccus)丰度较高。
2.9 显著差异菌群和显著差异代谢物联合分析
关联分析是数据深入挖掘的一种方式,主要用来发现隐藏在数据集中的关联性或相关性。16S rDNA与代谢的关联分析主要是基于统计学的算法发现显著差异菌群与显著差异代谢物之间的关联。为直观地反映显著差异菌群和显著差异代谢物表达模式的异同,对差异显著的菌群和代谢物进行Spearman相关性层次聚类分析。
层次聚类热图中每一行表示一个差异显著的菌属,每一列表示一种差异显著的代谢物。左侧树杈表示对差异菌属进行聚类的结果,上侧树杈表示对差异代谢物进行聚类分析的结果。聚类出现在同一个聚类的显著差异代谢物或者差异的菌属具有相似的相关性模式。层次聚类热图中每一个格子包含两种信息(相关系数r和P值)。r以颜色表示,r>0表示正相关(红色);r<0表示负相关(蓝色),颜色越深表示相关性越强。图7展示了185 对显著性相关的差异菌属和代谢物,其中58 对的P<0.01,相关性极显著。

图7 显著差异菌群与显著差异代谢物的Spearman相关性分析层次聚类热图Fig.7 Hierarchical clustering heatmap of Spearman correlation analysis of significantly differential intestinal microflora and metabolites
A组和D组样本在代谢组学层面上以及菌群多样性上具有明显的区分,共筛选到12 个显著差异菌群与60 个显著差异代谢物。通过Spearman统计学分析方法,对两者之间的相关性进行计算,共找到72 对具有显著性正相关的菌群-代谢物,113 对具有显著性负相关的菌群-代谢物。
3 讨论与结论
前期研究TG在体外水平上,可以显著抑制HepG2细胞增殖,诱导细胞凋亡、坏死[21]。因此需利用动物模型进行体内检测,从而对抗肿瘤药物进行全面分析解释,为肉苁蓉临床应用提供更准确的理论依据。选用HepG2细胞接种在裸鼠右后腿处,建立裸鼠皮下实体瘤模型,造模成功后,灌胃不同质量浓度TG,在整个饲喂过程中,测量小鼠体质量变化并分析小鼠的肿瘤体积增长趋势,监督观察其精神状态、饮食和活动能力,结果发现模型组小鼠精神状态较差,而处理组精神状态良好,且皮肤红润,此外,TG不会影响小鼠的正常生长,可以抑制小鼠皮下肿瘤体积的增长。
在HE染色中,苏木精染液呈碱性,可使细胞核着蓝紫色;伊红染液呈酸性,可使细胞质和细胞外基质着红色。该实验采用HE染色小鼠肝脏和肿瘤组织,发现TG可以减轻肝脏病理变化,缓解炎性细胞浸润现象。此外,TG在保护肝脏的同时[23],破坏肿瘤组织中肿瘤细胞结构,导致细胞坏死。HE染色结果直观地反映了肉苁蓉在肝癌小鼠中的肝保护和抗癌作用。
CAT是过氧化物酶体的标志酶,促使过氧化氢分解为水和氧分子,使细胞和组织器官免受H2O2的毒害[24]。GPT是一种参与人体蛋白质新陈代谢的酶,是肝细胞损伤最敏感和特异的指标。GOT是反映肝细胞坏死程度的标准,主要存在于肝细胞线粒体中,可以通过检测肝脏组织中GOT的含量确定肝脏组织病变程度。AKP是一种膜结合酶,广泛存在于人体各脏器中,其中以肝脏含量最高。临床研究表明,当肝系统受到恶性肿瘤细胞刺激时,AKP水平增加[25]。通过检测肝组织抗氧化指标,发现TG可以缓解小鼠肝脏氧化损伤程度,改善肝脏抗氧化功能,减轻肝癌小鼠肝脏病变程度,从而起到抑制小鼠体内肿瘤生长的作用。阻止肿瘤细胞侵害肝脏,为肉苁蓉作为辅助治疗或中医治疗药物在肝癌临床应用上提供一定的实验依据。
有大量临床数据和实验研究表明,许多癌症患者均有免疫功能低下的症状,首先是细胞免疫功能较低,在肿瘤生长过程中,体液免疫也在慢慢降低。有研究报道,肉苁蓉及其活性成分可以缓解化疗药物对免疫细胞造成的损伤,改善机体免疫功能[26-27]。此外,肉苁蓉多糖能够促进小鼠淋巴细胞的增殖并释放Ca2+,细胞内Ca2+的升高能够促进免疫因子的表达,循环调节淋巴细胞的增殖[28]。本实验表明肉苁蓉可以成为化学药物治疗中解决免疫能力低下的方法之一。
研究报道肝细胞癌患者在门静脉和粪便样本中的亚油酸和苯酚水平低于健康对照组,亚油酸和苯酚可显著抑制肝癌细胞增殖,表明代谢物在肝细胞癌的发病机制中起重要作用[29]。研究发现茴香霉素激活p38丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通过介导H3S10磷酸化促进HCC细胞的铁死亡[30]。伊马替尼可能通过miR-124和STAT3/HLF/IL-6通路发挥其抗肝纤维化活性[31]。巨大戟醇可减轻肝和胃肠道损伤,减少腹水体积,加剧H22细胞凋亡程度,改善异常的促炎细胞因子[32]。Nair等[33]发现拉贝洛尔与细胞分裂周期因子20表现出显著的相互作用,可作为一种潜在的抗肝癌靶向药物。原海葱苷A对多种HCC细胞系具有很强的抗癌作用,可显著抑制HCC细胞增殖、迁移和侵袭,与细胞凋亡、线粒体损伤、自噬密切相关,且对MHCC97H异种移植裸鼠中HCC细胞也有明显的抑制作用[34]。本研究结果显示,肉苁蓉可通过调节代谢物水平抑制肝癌的发展。此外,负离子模式下,D组中正缬氨酸较A组升高,水杨酸较A组降低,这与肉苁蓉多糖缓解肠道微生物失调相关患者的学习和记忆障碍结果[35]一致。
研究发现,卟啉单胞菌属会产生纤维化介质(例如转化生长因子-β1),加剧非酒精性脂肪型肝炎的纤维化[36]。D组中较A组卟啉单胞菌属相对丰度低,表明TG可以缓解肝纤维化,从而抑制肝癌的发生。此外,一项研究中提到Tyzzerella_3可作为预测胃癌的潜在标志物[37],由此推测模型组可能已经发生了肿瘤转移情况,后续需要进一步研究探讨。TG可以改善丁酸球菌属相对丰度,这与Mu Hongna等[38]研究柚皮苷减轻高脂肪饮食诱导的非酒精性脂肪肝病结果一致。表明TG可通过改善肠道微生物组成,减轻肝细胞持续损伤,抑制肝炎的发生,并在肝损伤后及时进行肝修复,在肝癌未发生时起到预防干预作用。
综上,肉苁蓉作为食药同源物质,富含营养价值和功能成分,可改善机体免疫能力,抑制肿瘤生长代谢,阻止肿瘤侵袭转移,调节机体代谢水平,改善机体肠道微生物组成等,表现出一定的抗肝癌作用。本实验关于TG抗HepG2肝癌效应的相关研究可以为肉苁蓉的后期开发利用和推广提供一定的理论依据和技术支撑。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
中老年保健(2022年2期)2022-08-24
基层中医药(2021年11期)2021-06-05
科学(2020年4期)2020-11-26
甘肃林业(2016年2期)2016-11-07
分析测试学报(2015年7期)2016-01-13
动物营养学报(2015年10期)2015-12-01
质谱学报(2015年5期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
新疆农垦科技(2014年10期)2014-02-28
