
壳聚糖/姜黄素光动力复合涂膜对金黄色葡萄球菌的抑菌效果及机理
2024-04-08 02:41张鹏敏王文秀孙剑锋陈志周马倩云
食品科学 2024年6期
张鹏敏,王文秀,孙剑锋,陈志周,马倩云,王 颉
(河北农业大学食品科技学院,河北 保定 071000)
金黄色葡萄球菌(Staphylococcus aureus)通过产生耐高温和耐酸的肠毒素而导致食物中毒,是最常见的食源性致病菌之一。由于S.aureus对高温、高盐等条件具有一定耐受能力,因此常规灭菌手段难以清除。每年因S.aureus所引发的食源性疾病在全球多个国家和地区均有暴发。据欧洲食品安全局报道,2021年欧洲境内由S.aureus细菌毒素引起的病例占总体食源性病例的1.5%,位居细菌毒素感染疾病第2位[1]。美国国家监测局报告指出,2009—2018年间美国各州食源性疾病暴发227 321 次,其中由包括S.aureus在内的细菌毒素引起的事件占7.5%[2]。在我国,2020年由S.aureus引起的食物中毒事件占全国食品中毒事件的1.1%[3]。国家疾病预防控制中心也将S.aureus列入引起我国细菌性食物中毒事件的主要致病因素之一[4]。因此,如何高效清除S.aureus成为研究热点。
最近,越来越多的数据表明,光动力灭活(photodynamic inactivation,PDI)是一种有前景的冷杀菌技术,可以抑制细菌、霉菌、病毒和寄生虫等多种微生物生长[5]。PDI处理的抑菌原理是在特定波长光照下,光敏剂吸收具有能量的光子并由基态进入激发态,迫使氧分子发生电子转移,产生活性氧(reactive oxygen species,ROS),ROS与细菌细胞内多种分子相互作用,进而抑制细菌生长[6]。PDI处理仅需要光敏剂、氧气和与光敏剂匹配的光源。相比于其他冷杀菌技术如超声、射线等具有耗能低、绿色环保等优势。目前常见的光敏剂包括5-盐酸氨酮戊酸[7]、酞菁锌[8]、亚甲基蓝[9]、玫瑰红[10]、竹红菌素[11]、金丝桃素[12]、叶绿素[13]以及姜黄素[14]等。
姜黄素是一种从姜黄根茎中提取的多酚化合物,常作为食品着色剂,具有抗增殖、抗氧化和抗菌等多种生物活性。因此,姜黄素在食品中的研究最为广泛。姜黄素分子本身具有抗菌活性,对S.aureus、铜绿假单胞菌[15]和单核细胞增生李斯特菌[16]等均有抑制作用,但是该抗菌活性效率较低、耗时长。研究表明,在400~470 nm激发波长下,姜黄素可快速产生具有细胞毒性的ROS从而清除细菌。但是,由于ROS半衰期短,光敏剂需尽可能接近目标细胞,从而对细胞产生破坏性损伤。然而在混合液中有约90%的姜黄素不能与细菌结合处于游离状态,导致姜黄素产生的ROS利用度极低[17]。
为改善上述问题,本研究拟利用壳聚糖作为连接细菌和姜黄素的纽带,增强姜黄素和细菌的结合能力。主要基于壳聚糖可作为分子载体[18-20],吸附游离姜黄素;同时壳聚糖特有的氨基结构可与菌体细胞壁表面带负电荷的蛋白质、磷脂等产生静电吸引[21-23],增加姜黄素特异性附着点数量,提高ROS的利用率。壳聚糖作为一种生态友好型生物材料,还具有较强的抗菌活性。此外,本研究制备的壳聚糖/姜黄素抗菌膜具有黑暗/光照双重抗菌性能。在实际应用中,亟需快速高效灭菌时,可采用光照处理加速灭菌过程。为了达到上述目的,本研究首先重点考察了壳聚糖/姜黄素双重抗菌涂膜对S.aureus的抑制效果,并从细胞结构和分子层面揭示其抑菌机理,旨在为新型冷杀菌技术在食品安全领域中的应用奠定理论基础。
1 材料与方法
1.1 材料与试剂
姜黄素、壳聚糖、琼脂粉、2,7-二氯二氢代荧光黄二醋酸(2,7-dichlorodihydrofluorescein diacetate,DCFHDA)、丙二醛(malondialdehyde,MDA)含量检测试剂盒和过氧化物酶(peroxidase,POD)活性检测试剂盒北京索莱宝科技有限公司;S.aureus(ATCC6538)上海士锋生物科技有限公司;碱性磷酸酶(alkaline phosphatase,AKP)试剂盒、超微量Na+/K+-ATP酶(ATP酶)试剂盒和总超氧化物歧化酶(superoxide dismutase,SOD)测定试剂盒(WST-1法)南京建成生物工程研究所;碘化丙啶(propidium iodide,PI)、羧基荧光素二乙酸酯(carboxyfluorescein diacetate,cFDA)上海麦克林生化科技股份有限公司;安全核酸染料Safe Red 莫纳(苏州)生物科技有限公司;细菌基因组DNA提取试剂盒 天根生化科技(北京)有限公司;LB肉汤 浙江海波生物技术有限公司;氯化钠、乙酸 国药集团化学试剂有限公司。
1.2 仪器与设备
Guava EasyCyte流式细胞仪 美国路名克斯贸易有限公司;FV1200激光共聚焦显微镜 日本奥林巴斯工业有限公司;420 nm光源 江苏省吴江区松陵镇辉恒灯具商行;TU-180紫外分光光度计 北京普析通用仪器有限责任公司;Nicolet iS20傅里叶变换红外光谱仪美国赛默飞世尔科技公司;SCIENTZ-1500F超声波分散仪 宁波新芝生物科技股份有限公司;MIRA LMS扫描电子显微镜(scanning electron microscope,SEM)泰思肯(中国)有限公司。
1.3 方法
1.3.1 壳聚糖/姜黄素涂膜溶液(chitosan/curcumin solution,CCs15)制备和表征
壳聚糖溶液(chitosan solution,Cs)的制备参考Rhim等[24]的方法,稍作改动。取1.0 g壳聚糖粉末溶解于100 mL 1%(体积分数)乙酸水溶液中,并在450 r/min转速条件下,90 ℃加热10 min,制得Cs。取10 mg姜黄素粉末溶解于4 mL无水乙醇中,制备姜黄素储备液。然后,取60 μL姜黄素储备液于10 mL Cs中混匀,得到CCs15,于4 ℃环境中保存。
将CCs15、姜黄素水溶液和姜黄素乙酸溶液(curcumin acetic acid solution,Curs)置于光程为1 cm 的比色皿中进行紫外-可见光扫描,扫描范围300~800 nm[25]。将15 mL膜溶液倒置在直径为90 mm的塑料平板中,置于40 ℃真空干燥箱10 h,待冷却至室温后,将膜放置在相对湿度50%、(24±1)℃条件下24 h,供后续傅里叶变换红外光谱测试。采用KBr压片法制备测试样品,利用反射光谱模式对涂膜的官能团进行测量分析,分辨率4 cm-1,扫描次数为32,检测范围为400~4 000 cm-1[26]。
1.3.2S.aureus的培养
LB 液体培养基:称取LB肉汤粉末25.0 g溶解于1 000 mL蒸馏水中混匀,进行高压蒸汽灭菌(121 ℃,15 min)处理,备用。
LB固体培养基:称取LB肉汤粉末25.0 g和琼脂粉15.0 g溶解于1 000 mL蒸馏水中混匀,进行高压蒸汽灭菌(121 ℃,15 min)处理,备用。
S.aureus活化与培养:将冻存于-80 ℃冰箱中的S.aureus于室温条件下解冻,然后接种到LB液体培养基中,在恒温振荡器(30 ℃、120 r/min)中培养。使用紫外分光光度计测定600 nm波长处吸光度以确定菌液浓度。
1.3.3 光动力涂膜对S.aureus抑制效果
1.3.3.1 光动力涂膜对S.aureus生长曲线的影响
参考Zhou Ting等[27]的方法,稍作改动。菌液(OD600nm=0.5)、涂膜溶液(生理盐水、Cs和CCs15)和LB液体培养基以1∶2∶2(V/V)的比例混合;在37 ℃条件下培养10 min。然后将菌悬液在波长420 nm、光强20 mW/cm2光源处照射5 min。之后在37 ℃条件下培养30 min。孵育结束后,将200 µL不同处理的菌液分别转移至装有15 mL新鲜LB液体培养基的锥形瓶中继续培养(30 ℃、120 r/min),并在不同时间点取样,使用酶标仪测定菌液在600 nm波长处吸光度。分组如下:菌液与生理盐水混合培养后,接受光照,记为KB;菌液与Cs混合培养后,接受光照,记为Cs;菌液与CCs15混合培养后,不接受光照,记为CCs/dark;菌液与CCs15混合培养后,接受光照,记为CCs/light。
1.3.3.2 不同涂膜溶液对S.aureus抑制效果
实验方案参照1.3.3.1节,涂膜溶液选用生理盐水、Cs、Curs(姜黄素质量浓度为15 mg/L)和CCs15,设置光照组和无光照组。光照组记为KB/light、Cs/light、Curs/light和CCs/light;无光照组记为KB/dark、Cs/dark、Curs/dark和CCs/dark。光照后,在37 ℃培养20 h。孵育结束后,取100 µL菌悬液稀释涂布在LB固体培养基平板上,将平板放入恒温培养箱中37 ℃培养24 h。最后,统计琼脂平板上的菌落数(lg(CFU/mL))。
1.3.3.3 姜黄素质量浓度对S.aureus抑制效果的影响
实验方案参照1.3.3.2节。姜黄素质量浓度梯度为0、5、10、15、20、25 mg/L。不设置无光照组。
1.3.3.4 光照时间对S.aureus抑制效果的影响
实验方案参照1.3.3.2节,涂膜溶液选用CCs15,光照时间梯度为0、1、3、5、7、10 min。
1.3.4 光动力处理对S.aureus细胞死/活状态的影响
采用流式细胞仪分析不同光照时间对细胞状态的影响,参考Meng Lingyun等[28]的方法,稍作改动。实验方案参照1.3.3.1节。涂膜溶液选用生理盐水和Curs。生理盐水处理的菌悬液光照为5 min,记为KB。Curs处理的菌悬液分别光照0、0.5、3、5、10 min,依次记为Curs0、Curs0.5、Curs3、Curs5、Curs10。孵育结束后,每取1 mL待测样品加入10 μL质量浓度为1 mg/mL PI染料,4 ℃孵育10 min,再加入50 μL质量浓度为0.46 mg/mL的cFDA染料,37 ℃孵育15 min,多余染料离心去除。
利用流式细胞仪对样品上机检测,选择488 nm波长处激光激发,cFDA染料在525 nm波长处发射绿色荧光,PI染料620 nm波长处发射红色荧光。低速采集10 000 个细胞,测定样品中每个细胞前向散射(forward scatter,FSC)、侧向散射(side scatter,SSC)以及绿色荧光通道和红色荧光通道信号,并转化成数字信号,通过FlowJo v10软件对所获得数据进行分析。
1.3.5 光动力处理对S.aureus细胞内ROS含量的影响
采用激光共聚焦显微镜测定不同光照时间对细胞内ROS含量的影响,参考Misba等[29]的方法,稍作改动。在100 mL LB液体培养基中加入0.1 mL DCFH-DA储备液,制备含药培养基。S.aureus菌液离心取菌泥,用含药培养基重悬,使菌液OD600nm=1.0。孵育30 min后,用生理盐水洗涤细胞3 次。用LB培养基制备OD600nm=0.5的菌悬液,供后续实验。孵育结束后,激光共聚焦显微镜对样品上机检测,选择488 nm波长激光激发,发射波长为525 nm。分组情况和处理条件参照1.3.4节。
1.3.6 光动力涂膜对S.aureus细胞膜通透性的影响
1.3.6.1 核酸泄漏
孵育结束后,将菌液离心(3 000 r/min,10 min),小心取出上清液,稀释一定倍数后用紫外分光光度计测定260 nm波长处吸光度检测核酸[30]。分组情况和处理条件参照1.3.3.1节。
1.3.6.2 蛋白质泄漏
孵育结束后,将菌液离心(3 000 r/min,10 min),小心取出上清液,用考马斯亮蓝法测定上清液蛋白质[31]。取考马斯亮蓝G-250溶液5 mL,向其中加入上清液1 mL,静置3 min后,于紫外分光光度计595 nm波长处测定其吸光度。分组情况和处理条件参照1.3.3.1节。
1.3.6.3 MDA含量
采用硫代巴比妥酸法检测MDA含量。详细操作步骤参照MDA含量试剂盒说明书。分组情况和处理条件参照1.3.3.1节。
1.3.7 光动力涂膜对S.aureus形态的影响
采用已报道的方法[32]利用SEM对S.aureus形态进行分析。加盖、喷金后在SEM下放大15 000、20 000 倍观察。设置KB、Cs、Curs/light、CCs/dark、CCs/light处理组,处理条件参照1.3.3.1节。
1.3.8 光动力涂膜对S.aureus内源酶活性的影响
从内源酶活性(AKP、ATP酶、POD和SOD)变化的角度出发,深层探究光动力涂膜对S.aureus的抑制机理。孵育结束后,详细操作步骤参照AKP试剂盒、超微量Na+/K+-ATP酶试剂盒、POD活性检测试剂盒和SOD活性测定试剂盒说明书。分组情况和处理条件参照1.3.3.1节。
1.3.9 光动力涂膜对S.aureus细胞内DNA和蛋白质的影响
根据细菌基因组D N A 提取试剂盒说明书提取S.aureus基因组DNA。将基因组DNA加载到1 g/100 mL琼脂糖凝胶和5 V/cm电压的电泳设备上,开始DNA电泳分离。电泳30 min后,用Safe Red浸泡染色30 min。在凝胶成像仪下观察,激发波长为305 nm。
采用文献已报道的方法[32]对S.aureus的蛋白质质量进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)测试。电泳结束后,使用考马斯亮蓝R-250进行染色,后用醋酸甲醇的混合溶液脱色,摇床脱色过夜至胶条背景蓝色消失后即可看见明显的蛋白条带。分组情况和处理条件参照1.3.3.1节。
1.4 数据处理分析
每组实验进行3~5 个平行,实验数据采用Excel软件进行计算,结果以表示;以SPSS 26.0软件进行方差分析(最小显著性差异检验法),P<0.05表示差异显著,采用GraphPad Prism 9.0软件作图。
2 结果与分析
2.1 光动力涂膜对S.aureus抑制效果分析
通过绘制细菌生长曲线研究光动力涂膜对S.aureus的抑制作用。如图1A所示,CCs/dark、Cs组与KB组相比延滞期延长16 h,可能是壳聚糖分子水解肽聚糖、破坏细菌细胞壁结构、抑制细菌生长引起的。而CCs/light延滞期长达28 h,说明光动力涂膜与无光照涂膜相比抑菌效果更明显。该对比结果表明,PDI是一种高效的抗菌技术,未来在控制S.aureus食源性污染方向有巨大应用潜力。

图1 光动力涂膜对S.aureus的抑制作用Fig.1 Effect of PDI treatment on the inactivation of S.aureus
进一步对比了在无光照和光照条件下,不同涂膜处理后S.aureus的菌落总数,用于分析蓝光和壳聚糖对光动力涂膜抑菌效果的影响。图1B是不同溶液处理后的S.aureus菌落总数。数据显示,KB/light组与KB/dark组相比,菌落总数下降0.31(lg(CFU/mL))。这说明仅蓝光处理可以影响S.aureus的生长,但是不能有效抑制细菌繁殖。这主要与蓝光照射能分解S.aureus表面的肠毒素,降低细菌生长繁殖能力相关[33]。Curs/light与Curs/dark相比,菌落总数下降了1.5(lg(CFU/mL));而Cs/light与Cs/dark相比,菌落总数下降了1.13(lg(CFU/mL))。因为PDI处理产生的ROS有抑菌作用,但是由于姜黄素和细菌结合松散和ROS寿命短等缺点,不能有效发挥ROS的抑菌作用。而Curs/light、CCs/light与KB/dark相比,菌落总数分别下降了4.65、6.89(lg(CFU/mL)),说明壳聚糖促进了姜黄素和细菌的结合,增加了ROS利用率。由此可得,壳聚糖在PDI处理过程中发挥重要作用。同时根据图1结果可知,壳聚糖/姜黄素光动力涂膜抑菌过程中起抑菌作用的因素按抑制效果大小排列依次是:ROS>壳聚糖>姜黄素>蓝光。
图1C为姜黄素质量浓度对S.aureus抑制效果的影响,与KB组相比,S.aureus经PDI处理后菌落总数明显减少(P<0.05),并且姜黄素质量浓度每增加5 mg/mL,S.aureus菌落总数会下降1~1.5(lg(CFU/mL))。这与Wang Ziyuan等[34]的研究结果一致。因此,姜黄素质量浓度是影响PDI抑菌效果的重要因素。当姜黄素质量浓度达到25 mg/L时,没有观察到S.aureus菌落。而Górski等[15]的研究中无光照条件下清除3(lg(CFU/mL))的S.aureus需要姜黄素质量浓度达到39 mg/L。本研究与之相比,姜黄素质量浓度降低了14 mg/L。这一结果说明PDI处理抑菌效果更好,在姜黄素低质量浓度下就可以达到明显的抑菌效果。
在上述研究基础上,进一步探究了光照时间对S.aureus抑制效果的影响。如图1D所示,光照1 min的菌悬液与无光照组相比,S.aureus菌落总数下降了1.81(lg(CFU/mL))。随着光照时间的延长,S.aureus菌落总数显著下降(P<0.05)。这说明延长光照时间可以增强PDI处理对S.aureus的抑制作用。另外,对光照时间和姜黄素质量浓度作方差分析,发现光照时间对S.aureus的抑制作用影响大于姜黄素质量浓度。随着光照时间继续延长(7~10 min),菌落总数变化不明显。这一结果说明过长的光照时间对抑菌效果无促进作用。王洋等[35]研究发现光照20 min后,菌落总数随光照时间延长缓慢上升。这一结果可能与溶液温度有关,还可能与较长时间光照导致姜黄素降解生成阿魏酸和香草醛导致抑菌效果下降有关[36]。因此,PDI处理时要避免长时间光照。
2.2 光动力处理对S.aureus细胞的影响
2.2.1 对S.aureus细胞死/活状态的影响
由图2可知,不同时间光动力处理后,仅姜黄素孵育的S.aureus有89.8%是活细胞。这说明仅姜黄素处理能影响细菌生长,但是不能完全杀死细胞。PDI处理后,活细胞数量明显减少;同时随着处理时间延长,处于活的非可培养(viable but non-culturable,VBNC)状态的细胞数量逐渐增加。当光照时间为3 min时,处于VBNC状态的细胞达到36.2%。随着处理时间继续延长,VBNC状态的细胞数量减少,死亡细胞逐渐增多。这说明PDI处理短时间内会促进细菌进入VBNC状态,而适当延长PDI处理时间可以有效杀灭细菌。

图2 光动力处理后S.aureus双染色结果Fig.2 Results of double staining of S.aureus after PDI treatment
与PDI处理相似,脉冲电场、紫外辐照等多种抗菌方法均会诱导细胞进入VBNC状态,而且S.aureus进入VBNC状态与氧化胁迫和ATP含量下降相关[28,37]。尽管VBNC状态的细菌处于休眠状态,但它们仍然生成毒素。因此,必须在杀菌过程中清除VBNC状态的细菌。以上结果表明,PDI有清除细菌VBNC状态的能力。
2.2.2 对S.aureus细胞内ROS含量的影响
由图3可知,仅光照处理,细菌细胞内仅有少量ROS产生;无光照处理组(Curs0),发现微量绿色荧光,原因可能是S.aureus为抵抗姜黄素影响,细胞产生内源性ROS,随着光照时间延长,细胞内ROS含量呈先增多后减少的趋势,ROS含量在光照3 min时最高。外源性ROS进入细胞内部,而兼性厌氧菌体内含有SOD和POD等抗氧化酶,因此细胞本身具有ROS耐受性,少量ROS不会严重影响细胞活性。随着光照时间延长,ROS含量超出细菌耐受极限,防御系统崩溃,致使细胞死亡,荧光强度下降。

图3 光动力处理对S.aureus细胞内ROS的影响Fig.3 Effect of PDI treatment on intracellular ROS in S.aureus
结合图2结果可得:随光照时间延长,ROS产量逐渐增多,攻击细胞;同时细胞内ROS含量升高,大量细胞进入VBNC状态;当ROS产生量超出细胞耐受极限时,ROS杀死处于VBNC状态的细胞,细菌生命活性下降,表现为荧光强度降低。因此,PDI是一个损伤逐渐积累的过程,只有ROS达到一定程度才能起到杀死细菌的作用。
2.2.3 对S.aureus细胞形态的影响
图4为流式细胞仪对不同光照时间下S.aureus细胞形态的检测结果。理论上FSC与细胞大小有关,SSC与细胞膜、细胞质、细胞核膜有关。随着光照时间延长,中心区域不再集中,向左下和右上扩散,表现为细胞形态发生变化,大小不均,内容物含量差异增加。这说明PDI处理改变细胞形态,破坏细胞膜结构,导致细胞物质运输受阻和细胞内容物泄漏。这可能会导致部分细胞不能向外运输物质,细胞增大;也可能导致细胞内外渗透压失去平衡,发生细胞皱缩现象[38]。细胞SSC和FSC信号变小,说明ROS致使部分细菌细胞膜破裂形成细小碎片,并伴有内容物泄漏[39]。少量细胞SSC和FSC信号增强,细胞膜破裂,可能发生膜黏连现象。因此,PDI处理致使细胞死亡的原因可能是ROS作用于细胞膜,致使细胞膜严重受损或者物质运输受阻,最终细胞因为不能进行正常生命活动而死亡。

图4 光动力处理对S.aureus细胞形态的影响Fig.4 Effect of PDI treatment on cell morphology of S.aureus
2.3 光动力涂膜对S.aureus的抑菌机理分析
2.3.1 光动力涂膜对细胞膜通透性的影响
首先,研究不同溶液处理对S.aureus细胞膜通透性的影响。DNA作为遗传物质,存在于S.aureus细胞内部,最大吸光度在260 nm波长处;当细胞膜和细胞壁受到破坏时,其内部核酸物质泄漏,泄漏程度与细胞膜通透性相关,因此用上清液中核酸含量评价细胞膜通透性。如图5a所示,Cs、CCs/dark、和CCs/light组OD260nm与空白组相比分别上升了14.18%、19.15%和25.33%。可能是因为S.aureus细胞膜被壳聚糖、姜黄素和ROS破坏,核酸物质从细胞内泄漏到培养基中。因此,壳聚糖、姜黄素和ROS皆可引起S.aureus细胞膜通透性增加。而且CCs/light组中产有外源性ROS,对细胞膜通透性影响最显著(P<0.01)。S.aureus死亡和细胞膜通透性增加直接相关。进一步通过蛋白质泄漏实验评估PDI处理对细胞膜通透性的影响。如图5b所示,CCs/light组上清液中蛋白质量浓度极显著提高(P<0.01),说明ROS与壳聚糖和姜黄素相比有更强的破坏作用,这与路振康等[40]的结果一致。蛋白质泄漏情况与核酸泄漏情况相似,进一步说明PDI处理能够增加细菌细胞膜通透性。
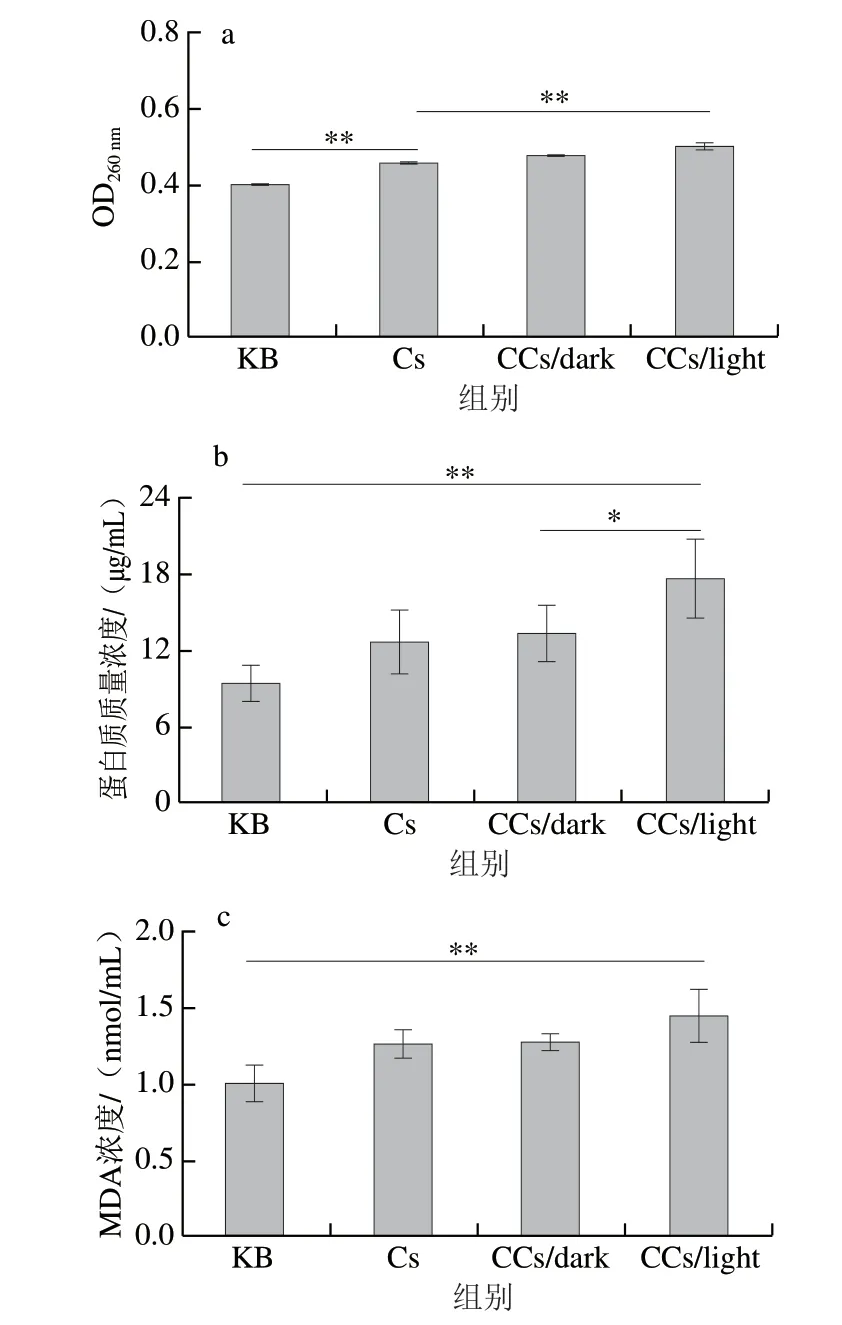
图5 光动力涂膜对S.aureus细胞膜的影响Fig.5 Effect of PDI treatment on cell membrane of S.aureus
如图5c所示,与空白相比,经CCs/light处理的细菌MDA浓度极显著增加(P<0.01)。MDA是细菌脂质膜过氧化的最典型产物之一。MDA浓度升高是细胞膜上磷脂分子氧化所导致的,磷脂分子的过度氧化会改变细胞膜通透性,这说明ROS通过氧化细胞膜磷脂分子进而影响细胞膜通透性。
2.3.2 光动力涂膜对S.aureus微观形态的影响
尽管通过流式细胞仪测定已经证明PDI处理会导致部分细胞形态变化,但具体结果尚不清晰。为进一步明确结构微观变化,研究采用SEM揭示PDI处理后S.aureus细胞微观结构层面的变化。如图6a所示,KB组S.aureus表现出正常的细胞形态,呈球形或椭圆形,大小均一、形态完整、表面光滑。在Cs组中,经壳聚糖涂膜溶液处理后细菌表面附着大量壳聚糖颗粒(图6b),表明壳聚糖可以吸附于细菌细胞壁表面。此外,细胞表面粗糙、出现褶皱和裂痕,少量细胞表面出现孔洞,但大部分细胞仍维持原有形态。这是由于壳聚糖与S.aureus表面蛋白质、磷脂等发生静电吸引作用,改变了细胞壁结构[22]。经CCs/dark处理的S.aureus表面也吸附了大量颗粒(图6d),而与Cs、Curs/light处理相比(图6b、c),CCs组表面颗粒大小明显高于两组,说明壳聚糖的存在增加了姜黄素在细胞表面的吸附量。为了进一步验证该结果,采用紫外-可见全光谱扫描测定了壳聚糖对姜黄素的吸附效果,结果如图6f所示。由姜黄素的最大吸光度可知,姜黄素在蒸馏水和乙酸溶液中溶解度较差,而在Cs中溶解度明显高于二者,这是由于壳聚糖分子载体作用[18-20]。为了揭示二者的相互作用,测定二者的傅里叶变换红外光谱。结果表明,壳聚糖和姜黄素之间未产生新的化学键,但是羟基(3 283.96 cm-1)的强度和波数发生了变化(图6g),这是由非共价相互作用和氢键共同作用的结果,该结果与Roy等[18]研究结果相同。上述结果表明,壳聚糖发挥了纽带作用,增强姜黄素和细菌的结合能力。结合之后的CCs,经PDI处理后,S.aureus细胞呈现严重损伤,细胞膜遭到破坏,细胞内容物大面积泄漏,失去细胞正常形态,形成很多碎片和空壳(图6e),该结果与Yuan Yuan等[41]的报道相似。与Curs/light结果对比说明,壳聚糖增强了姜黄素对细菌的PDI效果,同时也说明对细胞产生不可逆损伤,最终造成细胞破裂是壳聚糖、姜黄素和ROS共同作用于细胞膜和细胞壁的结果,其中ROS影响最大。图6结果在验证上述核酸、蛋白质和MDA测定结果的同时,说明壳聚糖在连接姜黄素和细菌过程中发挥重要作用,同时也论证了PDI处理破坏细菌细胞结构完整性的结论。细胞膜作为细胞屏障,其完整性是细胞生长、繁殖和其他生理活动的基础,细胞膜破损程度,直接影响细胞生命活性[42-43]。PDI处理通过作用于细胞膜,破坏细胞膜和细胞壁结构,促使细胞内核酸、蛋白质、多糖等营养物质泄漏。随着破坏程度增强,细胞内营养物质含量降低,将无法维持细胞正常生命活动。

图6 SEM观察光动力涂膜处理后S.aureus微观形态和涂膜溶液表征Fig.6 Microscopic morphology of S.aureus after PDI treatment observed by SEM and characterization of CCs
2.3.3 光动力涂膜对S.aureus细胞内源酶活性的影响
AKP在微生物体内可直接参与磷酸基团的转移和代谢,存在于细胞壁和细胞膜之间。细胞壁受损时,AKP由胞内渗出,细菌磷酸基团代谢途径受到影响。如图7a所示,经Cs处理后,S.aureusAKP活性极显著高于KB组(P<0.01)。这是因为壳聚糖分子增加了S.aureus细胞壁通透性,使AKP从细胞膜中泄漏出来。而CCs/light处理后S.aureus较Cs组AKP活性下降。这是因为ROS非靶向攻击蛋白质的一级结构,影响二级结构形成,破坏AKP完整性,促使AKP活性位点消失,进而使S.aureusAKP活性下降。AKP活性改变,说明ROS能破坏细胞壁结构,同时攻击细胞内源酶,破坏细胞正常代谢途径。

图7 光动力涂膜对S.aureus酶活性的影响Fig.7 Effect of PDI treatment on enzyme activities in S.aureus
ATP存在于细菌细胞膜中,通过糖酵解过程产生。细菌细胞膜上有多种酶可以促进ATP生成和代谢。ATP酶是这种机制中最重要的酶之一,其主要是利用ATP水解产生的能量对Na+、K+、Ca2+、Mg2+等进行由低浓度至高浓度运输,保持膜内外离子浓度的平衡,维持正常生命活动,因此能够通过ATP酶活性变化分析PDI处理对菌体细胞的影响。如图7b所示,Cs、CCs/dark、与KB组相比ATP酶活性下降,CCs/light组极显著低于KB组(P<0.01)。这是因为壳聚糖和姜黄素破坏细胞壁结构,诱导质子动力势(proton-motive force,PMF)崩溃。PMF是由Δψ和ΔpH组成的电化学梯度,PMF驱动膜运输系统促进ATP合成和离子及其他代谢物的积累[44]。细胞膜内外PMF崩溃导致ATP酶合成相应减少。而CCs/light组ATP酶活性最低,壳聚糖的存在使ROS更容易进入细胞内并作用于ATP酶,导致ATP酶活性进一步下降。ATP酶失活使细胞无法通过分解ATP实现能量转移以维持生命活动,最终导致S.aureus死亡。
图7c、d分别是不同处理条件下S.aureus细胞内SOD、POD活性。CCs/light处理组与KB组相比S.aureus细胞内SOD和POD活性极显著增加(P<0.01)。这是因为PDI处理产生的ROS进入细胞内部,细菌防御机制启动,以SOD、POD为代表的抗氧化酶大量表达以维持内部ROS平衡并保护细胞免受损伤。SOD将ROS催化为H2O2,随着H2O2含量升高,细胞快速转录翻译,产生POD将H2O2降解为H2O。因此,SOD和POD活性升高是细菌为了应对高ROS环境做出的反应。该结果与杨璟[45]的研究结果相似。
总之,PDI处理产生的ROS会引发细菌防御机制启动,SOD、POD活性升高。随着细胞壁被破坏,大量ROS进入细胞内部。最后,因为ROS和H2O2含量过高,细胞内SOD、POD和其他抗氧化酶不能将其恢复到正常水平,进而影响AKP和ATP酶活性。结果导致S.aureus生物合成、磷酸转移、能量运输途径受阻,从而抑制S.aureus的生长。
2.3.4 光动力涂膜对S.aureus细胞内DNA和蛋白质的影响
通过琼脂糖凝胶电泳测定光动力涂膜对S.aureus基因组DNA的影响。如图8a所示,空白对照组DNA电泳条带亮度高,图像清晰;而CCs/light组的基因组DNA条带亮度降低,清晰度降低。ROS氧化作用是导致细胞DNA损伤的主要因素之一[46]。外源性ROS渗透和内部抗氧化防御系统崩溃导致细胞内ROS含量急剧上升,进而损伤DNA[47]。另外,外源性ROS可直接与核酸物质反应,DNA发生氧化损伤,如单链或双链切割、交联和碱基序列变化。DNA损伤导致转录过程中断或破坏,进而导致细胞死亡。同时,ROS、壳聚糖和姜黄素可作用于参与DNA修复的酶(如DNA聚合酶),导致酶失活或功能改变,造成DNA损伤无法修复[48]。细胞中产生的脂质过氧化产物,如MDA,也可以导致DNA和蛋白质交联,引起DNA损伤[49]。
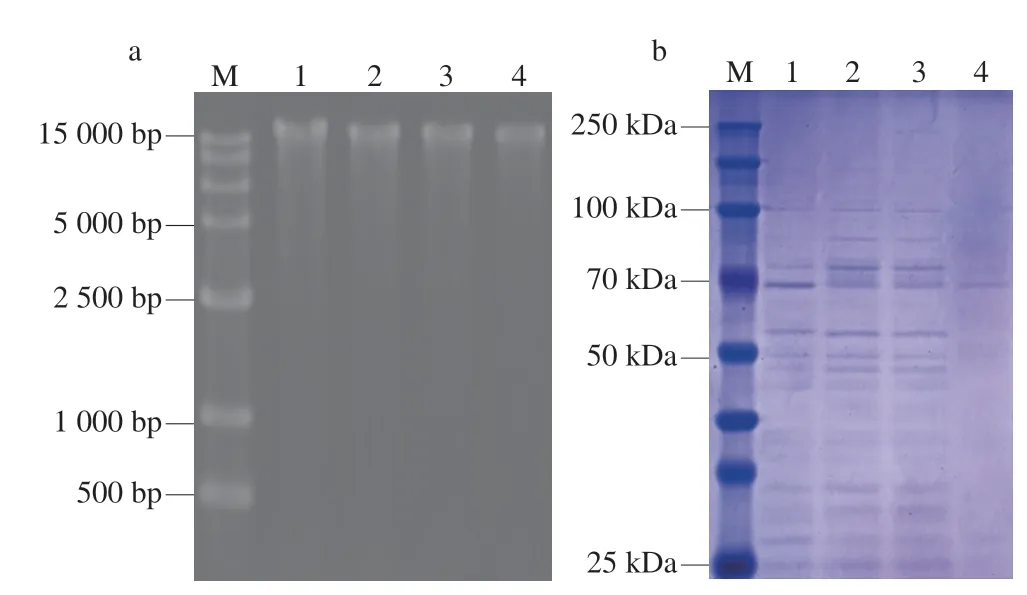
图8 光动力涂膜对S.aureus核酸和全细胞蛋白质的影响Fig.8 Effect of nucleic acid and whole cell proteins in S.aureus after PDI treatment
通过SDS-PAGE测定光动力涂膜对S.aureus蛋白质的影响。观察电泳图像发现,与KB组相比,Cs和CCs/dark组在70 kDa左右的条带亮度下降,80 kDa左右的条带亮度提高(图8b),可能和细菌的应激反应相关。而CCs/light组的蛋白质条带严重降解,说明ROS在细胞内部非定向氧化,使得细胞内蛋白质分子氧化变性,丧失原本的结构。Dosselli等[50]的研究同样证明了ROS攻击蛋白质使得一级结构破坏,丧失生物活性;并且芳香族氨基酸容易受到ROS的攻击[51]。部分胞内酶由蛋白质构成,蛋白质结构损伤也是影响酶活性的重要原因。另外,基因组DNA损伤影响RNA转录翻译,细菌蛋白质合成能力下降是蛋白质条带降解的重要原因。
综上所述,ROS通过破坏细胞膜致使细胞内ROS含量急速增加,最后因细胞内ROS含量过高导致细胞防御系统崩溃,损伤细菌基因组DNA和蛋白质等生物大分子。因此,PDI清除S.aureus是多靶点致死效应导致的结果,而ROS破坏细菌体内DNA和蛋白质导致细菌产生不可逆损伤,是造成S.aureus死亡的重要因素。
3 结论
为促进姜黄素与细菌的结合,改善PDI处理时姜黄素利用率低的问题,本研究利用壳聚糖的细菌黏附性特点制备了壳聚糖/姜黄素光动力涂膜以增强PDI抑菌效果。在此基础上,探究了影响涂膜抑菌效率的关键因素及PDI抑菌机理。结果显示,PDI处理对S.aureus有优异的抑制作用,姜黄素质量浓度为25 mg/L的壳聚糖涂膜溶液在波长为420 nm蓝光处光照仅5 min,对S.aureus可达到99.9%以上的灭活效果。壳聚糖的加入增强了姜黄素和细菌的结合,有利于PDI处理产生的ROS直接作用于细胞膜,破坏细菌第一层屏障,进入细胞内,致使细菌细胞形态改变,蛋白质和DNA泄漏。同时,ROS作用于AKP、ATP酶、POD和SOD等内源酶,进而破坏细胞内生物合成、能量转化和细胞防御系统,并且影响细胞内DNA和蛋白质等生物大分子,影响RNA转录、DNA自我修复、酶的合成和细胞膜修复,最终导致S.aureus死亡。综上,PDI是一种损伤逐渐积累的多靶点杀菌技术。本研究结果可为PDI在食品安全领域中的应用提供理论基础。
猜你喜欢
塑料包装(2021年3期)2021-01-25
Medical Data Mining(2019年2期)2019-07-16
中成药(2018年2期)2018-05-09
中成药(2018年3期)2018-05-07
中国医疗美容(2015年4期)2015-04-27
科学中国人(2015年13期)2015-02-28
西南军医(2015年1期)2015-01-22
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年21期)2014-03-11
食品科学(2013年15期)2013-03-11
