
观赏蕨类姬蕨同配子体自交的孢子体萌发和发育特性
2024-04-11 07:15杨春萍
园艺与种苗 2024年1期
李 莉,杨春萍,商 辉*
(1.上海辰山植物园,华东野生濒危资源植物保育中心,上海 201602;2.唐山汉沽管理区农业技术服务中心,河北唐山 050701)
观赏蕨类属于观叶植物,具有美化、彩化和净化等功能。别具特色的观赏特性、耐阴喜湿的生态习性使蕨类植物在园艺市场上的应用越来越广泛,需求量也逐渐增大。蕨类植物虽没有艳丽的花朵,但在色彩、形态、习性等方面独树一帜。如:叶型奇特,从单叶至复叶;形体相差悬殊,小到匍匐地被,大到数米生长,高大挺拔,生机盎然;孢子囊群造型独特,圆形、肾形、线形等,其着生部位既有叶脉顶端、亦有叶中部位,变化多端,是种子植物不具备的观赏价值。此外,蕨类植物的根状茎、叶片、孢子囊也常常具备独特的观赏价值。
蕨类植物在园林绿化中的应用形式多样。在室内以盆栽及微景观形式应用于大厅、主题会场、家庭及办公室等;在城市建筑如商场、办公楼、宾馆的庭院绿化、阴生专类园、水景园、家庭庭院布置及城市园林绿化带可使用蕨类植物作为地被及中下层植物[1-4]。
中国现有蕨类植物38 科,177 属,2 215 种,228 个种下分类群,其中有1 200 余种属于中国特有[5-6]。目前国内观赏蕨类在园艺市场上依旧呈现出种类少及应用单一现象。观赏蕨类植物相比种子植物占比极小,而野生蕨类植物的开发利用则是补充种类多样性的有效途径。
姬蕨(Hypolepis punctata)隶属于碗蕨科(Dennstaedtiaceae)姬蕨属(Hypolepis),生长在热带亚热带区域,国内主要分布于长江中下游及以南地区,喜温暖多湿半阴环境,在弃耕地、山谷溪流边和林缘、林窗中经常可以发现[7],是园林观赏和家庭种植的优良物种。早在1994 年姬蕨就是英国最流行的观赏蕨类植物之一[8],但在国内的应用几乎没有。姬蕨植株姿态飘逸优美、适应性强、易繁殖、资源量大,可以作为基础种植材料应用于园林绿化中,也可应用于阴暗角落或高大乔木下较阴暗处,特别是在园林中较难处理的墙边、篱边、路边的配置。飘逸优美的羽状复叶还可以作为切花配叶。总之,姬蕨具有较好的园艺市场应用前景。
目前姬蕨的人工扩繁以孢子萌发培养为主。由于配子体产生精子器和颈卵器的时期不同,甚至有些配子体可能一生只产生精子器,这使得在孢子批量混播的情况下,精卵结合绝大多数情况下发生在不同配子体之间,即配子体间。然而,园艺上进行观赏蕨类的培养,混播的方式很难保证性状的稳定性。为保持优良蕨类品种的性状稳定性,许多蕨类开展了组培快繁技术,如波斯顿蕨[9]。可以通过对单个孢子进行培养,或在配子体产生精子器和颈卵器前移栽进行单独培养,人为创造条件进行配子体内自交,从而保障品种性状的稳定性。但由于蕨类生活史的特性,配子体内自交可产生基因组上所有等位基因完全的纯合子,可能面临着较严重的近交衰退[10],因此该蕨类植物繁殖方案需要繁殖生态学的评估。
蕨类的扩散,一般认为是以风力为主导,特别是一些大洋岛屿的蕨类植物分布证据,使得风的长距离传播能力得以印证[11]。风媒传播下,单个孢子的传播将不可避免,配子体上颈卵器成熟时将无法得到其他配子体的精子,而不得不进行配子体内自交。所以借助风力进行种群扩张的同型孢子蕨类建立的新种群,配子体内自交占主导,遗传多样性低下。许多试验证明,自交对于同型孢子的蕨类植物繁殖不利。人们普遍认为蕨类植物孢子萌发后同配自交是有害的,异交才是蕨类配子体主要的繁殖模式[12-13]。
绝大多数蕨类植物产生同型孢子,即其孢子萌发产生的配子体既可以长出精子器产生精子,也可以长出颈卵器产生卵细胞,因此蕨类植物的交配模式可分为3 类:异配异交,即来源不同母本的配子体间的精卵结合;异配自交,即来源同一母本的配子体间的精卵结合;同配自交,即同一配子体间的精卵结合[10]。蕨类植物的孢子散播之后,大部分都散落在母株附近,只有小部分孢子长距离散播到其他地方。近距离散播孢子发育成配子体后,周边以来源于相同母株的配子体为主,少量同其他植株来源的配子体,所以交配模式以异配自交为主、异配异交为辅。长距离散布的孢子,很难落在同种其他孢子旁边,因此同配自交是其唯一可选择的繁殖方式。由于遗传负荷(致死基因或有害基因)的存在,推测同配自交产生的后代适应性可能较弱,甚至直接死亡。但是如果同配自交发生概率高,可能会产生多代的过滤作用。即有害基因在长期的迭代过程中已经被淘汰,同配自交的适合率可能并不一定显著低下。而在常规孢子培养中,蕨类的交配系统对产生和质量影响不够重视,单独孢子培养不常见,而通过混播孢子进行蕨类繁殖,其后代可能来自于异配自交或异配异交,因为无法确定野外收集的孢子是否来自于不同植株,该文统称它们为异配混交。
该研究以姬蕨为材料,通过配子体单独培养的方法,检测同配自交是否产生严重的近交衰退,从而探讨该方法在观赏蕨类品种优良性状保持上是否具有可行性。
1 材料与方法
设计配子体培养试验,比较单独配子体和多配子体的孢子体萌发,即同配自交与异配混交的萌发状态(萌发率、存活率),孢子体成长情况及生长速率。播种生长情况及步骤如图1。

图1 两种交配系统的处理及播种生长情况
1.1 孢子采收与保存
姬蕨孢子来自于湖南桑植县野外种群。采集孢子时,取其成熟且尚未开裂的孢子叶,去除叶面杂质,置于干净的信封或硫酸纸袋中,使用变色硅胶于封口袋中干燥5~10 d。收集孢子时,拍打信封或硫酸纸袋致使孢子得以充分脱落,并移至干净的硫酸纸上,去除多余的叶片及孢子囊群盖,即可用于培养,或将其收集于离心管中,置于-20℃冰箱保存[14]。
1.2 播种方法及培养过程
播种时所需基质选用经过清水浸泡、剪碎的水苔,121℃灭菌锅20 min,将灭菌后的水苔均匀铺于组培瓶的底部,压实,使水苔平整,厚度为2~3 cm,无菌水浸润,备用。取适量孢子置于2 mL 离心管中,加入5% NaClO 水溶液1 mL,静置消毒5 min,8 000 r/min 离心5 min,待孢子沉淀后用无菌水冲洗3~5 遍,放入容器中制成悬浊液,均匀喷洒在组培瓶中的水苔上,加上透明盖。保持培养微环境温度为20℃~25℃,白炽灯光1 500 lx,光照12 h/d,湿度保持90%以上。一般5~10 d 后萌发长出原丝体,后者不断发育,形成多细胞原叶体。
1.3 配子体隔离培养试验设计
待配子体萌出后,性器官精子器未长出之前进行配子体的移植。移栽基质选用腐殖土,用网孔直径小于2 mm的细筛筛出,121℃灭菌锅灭菌20 min[10]。后将基质置于培养皿中,厚度为1 cm。将配子体移植到腐殖土上,使基质湿润,加盖。成熟配子体上精子器在播种后50 d 左右成熟,颈卵器55 d 左右成熟,卵细胞在播种后2~3 个月完成受精,受精卵发育成胚胎,进而长成幼孢子体。
其中移植单独的配子体24 株于单独的直径9 cm 培养皿中,作为同配自交处理。另将24 个配子体按5×5 的网格(中间留空)排列移植于15 cm 的培养皿中,作为异配混交处理,如图1。每天统计2 种交配系统萌发情况,包括萌发的正常幼孢子体数量及萌发时间记录,连续统计36 d。计算2 种交配系统的幼孢子体的萌发率及萌发时间。同时每周记录幼孢子体的存活情况,连续记录7 周,计算幼孢子体的存活率。
1.4 孢子体移栽与测量
孢子播种120 d 后幼孢子体长到3~5 cm 时,具有2~3 片真叶即可进行分盆,移栽盆口径5 cm 左右,基质选用体积比5∶1 草炭土和蛭石。保持土壤湿润,环境温度在15℃~25℃,自然光照10~12 h/d,喷水每天1 次,遮蔽保湿,湿度大于70%。做好盆土水分管理、病虫害管理,以确保植物良好生长。每天测量统计两种交配系统孢子体生长情况,包括叶片的数量、宽度及长度,23 d 内测量6次(第1、6、11、15、19 和23 天),并计算生长速率和近交衰退系数(自交混交的差值除以混交的值)。自交和混交个体生长速率的差异用SPSS 软件进行t 检验。待孢子体继续长到10 cm 以上,具有10 片以上真叶可换盆至口径10 cm的盆器中培养,移栽时切记勿伤根,无须去除旧土。
2 结果与分析
2.1 不同交配系统下孢子体萌发及存活率情况比较
姬蕨孢子隔离培养试验表明,在相同且适宜的人工培养条件下,姬蕨配子体单独培养后,孢子体萌发率仍然较高,达到了87.5%;混合培养的孢子在萌发时间上早于单独培养4 d,且最终孢子体萌发率为100%(图2A)。在萌发出幼孢子体后,随着时间的推移,自交的个体存活率在不断下降,在最终的7 周统计时间内,自交个体的存活率已经下降到71.4%,最终存活率为62.5%(以配子体总数为基数),混交的存活率则依旧是100%(图2B)。
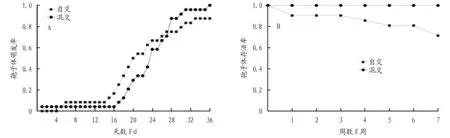
图2 两种交配系统孢子萌发率(A)及存活率(B)
2.2 不同交配系统下萌发个体的生长情况
姬蕨的孢子体幼苗经过移栽后的生长检测表明,包括植物的叶片数量、长度和宽度上一直弱于混交的配子体(图3)。同时,自交的生长速率也弱于混交个体(图4),其中叶片长度的生长速率自交个体为(0.13±0.12)cm/d,混交个体为(0.17±0.07)cm/d(P=0.206);叶片宽度的生长速率自交个体为(0.06±0.04)cm/d,混交个体为(0.09±0.05)cm/d(P=0.036);叶片数目的生长速率自交个体为(0.1±0.08)片/d,混交个体为(0.19±0.11)片/d(P=0.003)。近交衰退系数分别为24.8%、35.1%和46%。
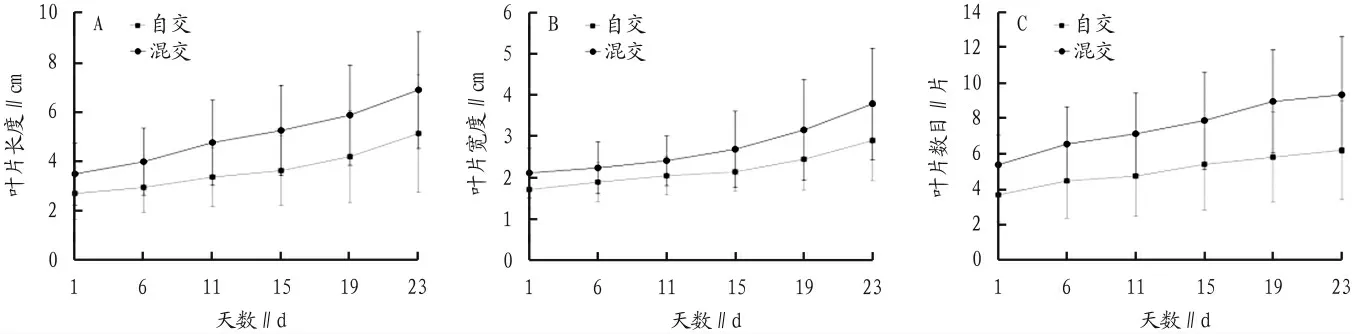
图3 两种交配系统幼孢子体生长速率

图4 两种交配系统幼孢子体生长速度
3 讨论
3.1 同配自交存在近交衰退
单独培养(同配自交)配子体上开始萌出孢子体的时间晚于混合培养(混交)配子体约4 d,这是因为颈卵器约晚于精子器出现4~5 d,只有等到颈卵器出现后,才会发生配子体内自交。单独培养和混合培养的孢子体萌出速度大致相当,最终混合培养的配子体全部萌出孢子体(100%),而单独培养的孢子体萌出率为87.5%,即24 个配子体中有3 个配子体受精失败或胚胎死亡,而萌出后又有6 个自交孢子体死亡。这些结果说明相比于配子体间异交,配子体内自交确实存在一定程度的致死率,总致死率为37.5%,配子体可能存在致死基因,在受精为孢子体后,由于形成纯合子,在胚胎发育以及幼苗生长阶段发挥作用,导致胚胎和幼苗死亡。幼苗移栽后的生长状况,同配自交的孢子体弱于异配异交的孢子体,特别是叶片宽度和叶片数目具有显著差异,这说明同配自交具有近交衰退。
3.2 同配自交的近交衰退率对种群延续的影响
不同于种子植物有大孢子和小孢子的区别,蕨类具有同形孢子,也就是说单个孢子产生的配子体既可以产生精子,也可以产生卵细胞。蕨类这种特殊的生活史,使得同配自交成为可能,其产生的孢子体基因组上所有的等位基因都会纯合。对于完全纯合的孢子体来说,只要有一个位点存在有害基因就会发挥作用,特别地如果是致死基因,会直接导致死亡。而该结果表明,至少高达62.5%的孢子没有致死基因(并不是所有的死亡都能归结到致死基因,但没有死亡可以说明没有致死基因),这说明在野外条件下,单个孢子同配自交的情况也经常发生,在强烈的自然选择作用下,致死基因得到了有效清除。
而在幼苗生长过程中,虽然同配自交的个体长势要弱一些。但以异配异交的个体为标准,同配自交的近交衰退率仅为24.8%~46%,这说明在自然条件下,有害(非致死)基因也得到一定程度的清除,使得同配自交的后代仍然保持一定生活力,也使得单个孢子也能在很大概率上延续种群。笔者将在后续的培养过程中逐代检测其生活力的变化趋势。
3.3 配子体单独培养在观赏蕨类繁殖上的应用前景
种子植物育种过程中,往往需要多代自交才可以得到稳定的自交系,而对于蕨类植物来说,一次同配自交就可以得到完全纯合的个体,这是蕨类植物育种的优势。该试验所用的孢子皆来自于野外采集,其致死基因和有害基因已经得到很大程度的过滤,在人工培养的情况下,可以继续进行人工选择,尽最大程度地清除致死基因和有害基因,同时保留优良基因,从而获得有价值且生命力强壮的园艺品系。
该研究证明了姬蕨同配自交产生的后代仍然具有较高的生活力,通过单独培养的方式可以在短时间内得到完全纯合的自交系植株,同时可以得到具有较高观赏特性和遗传稳定的品系,可作为进一步杂交育种的优良亲本,在园艺育种方向具有很好的应用价值,为观赏蕨类的育种提供理论支持。
致谢:感谢严岳鸿博士、王莹、齐新萍、邵文在本研究过程中特别是在孢子繁殖方面给予的帮助;感谢潘向艳对论文修改的建议。
猜你喜欢
渔业研究(2022年4期)2022-08-27
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
渔业研究(2019年1期)2019-03-04
西北植物学报(2018年6期)2018-07-19
飞碟探索(2016年9期)2016-09-21
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年3期)2016-05-09
中国民族医药杂志(2016年3期)2016-05-09
