
鸡neurexophilin 1基因在毕赤酵母中的表达
2015-10-26 05:49陈美玲聂彬姜勋平刘桂琼
生物技术通报 2015年3期
陈美玲 聂彬 姜勋平 刘桂琼
(华中农业大学动物科技学院,武汉 430070)
鸡neurexophilin 1基因在毕赤酵母中的表达
陈美玲 聂彬 姜勋平 刘桂琼
(华中农业大学动物科技学院,武汉 430070)
Neurexophilin 1是母鸡子宫阴道结合部位的差异表达基因。根据鸡nxph1的序列,结合毕赤酵母密码子的偏好性,合成nxph1基因,设计引物以合成基因为模板,扩增其去除信号肽的DNA序列△nxph1,将合成的nxph1基因及去除信号肽的nxph1基因片段△nxph1分别插入pPICZαA真核表达载体,构建重组表达质粒pPICZαA/nxph1和去除信号肽的重组表达质粒pPICZαA/△nxph1,电转入毕赤酵母GS115,阳性菌用终浓度为1%的甲醇诱导表达,SDS-PAGE电泳和Western blot检测表达产物,结果表明重组菌成功分泌鸡NXPH1蛋白和去除信号的△NXPH1蛋白,大小约29 kD,为探索NXPH1在母鸡生殖道内的功能奠定基础。
nxph 1;毕赤酵母;基因表达
Neurexophilin(NXPH)是一种从大鼠和牛的脑部分离纯化的神经糖蛋白,其成熟肽29 kD,因其与神经元表面蛋白Iα(neurexin Iα)紧密结合而命名[1]。哺乳动物至少含4个与NXPH相关的基因,分别是nxph1、nxph2、nxph3和nxph4,这4种基因在不同动物不同组织中的表达不一致,如人类可表达4种基因,而鼠类则只表达nxph1、nxph3和nxph4基因,且只有NXPH1和NXPH3可以与neurexin Iα结合[2]。朱金金等[3]发现nxph1是鸡子宫阴道结合部的差异表达基因,且该基因在高受精率组母鸡子宫阴道交接部的表达高于低受精率组,推测其可能与精子贮存相关。刘桂琼等[4]也发现在人工授精后,母鸡贮精腺表达神经系统内特异表达的Neurexophilin1基因,而高受精力母鸡贮精腺Neurexophilin1 表达量高于低受精力母鸡,因此神经调控可能参与贮精腺内的精子贮存。本研究根据酵母基因表达的密码子偏好改造鸡nxph1基因,在毕赤酵母GS115中表达该基因,以期为后续研究NXPH 1蛋白的生物学功能奠定基础。
1 材料与方法
1.1 材料
真核表达载体pPICZαA、毕赤酵母GS115菌株为华中农业大学国家微生物重点实验室郭爱珍教授课题组惠赠;克隆载体PMD-18T购自大连宝生物公司(TaKaRa)公司;大肠杆菌DH5α菌株购自北京全式金生物技术有限公司;
ZeocinTM购自Invitrogen公司;质粒提取试剂盒购自Omega公司;T4 DNA连接酶、TEMED、限制性内切酶EcoRⅠ、NotⅠ、SacⅠ均购于TOYOBO公司;neurexophilin 1抗体购自美国R&D公司。
1.2 方法
1.2.1 Nxph1基因合成和引物设计 根据GenBank公布的鸡nxph1基因(XM-418683)mRNA编码序列,结合毕赤酵母密码子的偏好性优化序列,在基因的两端添加EcoRⅠ和NotⅠ限制性内切酶位点,序列长为833 bp,序列依次为保护性碱基、EcoRⅠ酶切位点、氨基酸的编码序列813 bp、NotⅠ酶切位点和保护性碱基。依据优化后的碱基序列设计一对可扩增去除NXPH1蛋白信号肽的特异性引物P1和P2,并在引物序列的两端分别添加EcoRⅠ和NotⅠ两个限制性内切酶位点,其扩增产物为770 bp,其中氨基酸的编码序列长750 bp,将该序列命名为△nxph1。优化基因序列和引物由北京金唯智公司合成。
1.2.2 Nxph1基因的序列鉴定 以合成的nxph1基因为模板,分别以P1/P2为上下游引物进行PCR扩增。PCR反应条件:94℃ 预变性4 min,94℃变性30 s,55℃退火1 min,72℃延伸 30 s,40个循环,72℃延伸10 min。2%的琼脂糖凝胶检测PCR产物。DNA凝胶回收试剂盒(北京庄萌生物公司)回收PCR产物,与pMD18-T载体连接,转化DH5α,提取质粒后用EcoRⅠ和NotⅠ双酶切鉴定,阳性克隆菌液送上海生工测序。
1.2.3 酵母表达载体的构建 将△nxph1质粒、合成的基因nxph1和酵母表达载体pPICZαA用 EcoRⅠ和NotⅠ在37℃条件下双酶切3 h,2%的琼脂糖凝胶电泳,回收酶切产物,T4 DNA连接酶16℃连接过夜,构建成重组表达质粒pPICZαA/ nxph1和pPICZαA/△nxph1。连接产物转化DH5α感受态细胞,挑选菌落培养提质粒,用通用引物5' AOX和3' AOX进行PCR 鉴定,用 EcoRⅠ和NotⅠ进行双酶切鉴定,阳性克隆菌液送上海生工测序。
1.2.4 构建重组nxph1的酵母菌株 用 SacⅠ线性化重组质粒pPICZαA/ nxph1 和pPICZαA/△nxph1,取制备好的GS115感受态50 μL与10 μL线性化的pPICZαA/nxph1、pPICZαA/△nxph1质粒充分混匀后转移到预冷的0.2 cm电击杯中,放入Bio-Rad电转仪中1 500 V、50 μF和500 μs电击,立即加入1 mL冰冻的山梨醇液,冰浴静止10 min,菌液30℃孵育1.5 h,离心后弃上清液,取100 μL混合沉淀涂布在含有Zeocin(100 μg/mL)的YPD平板上,30℃培养至出现白色单菌落。挑选白色单菌落培养,采用“煮-冻-煮”的方法提取酵母基因组,用通用引物扩增,1 % 的琼脂糖凝胶电泳检测扩增产物。
1.2.5 NXPH1在酵母中的表达 阳性酵母重组质粒菌液接种BMGY培养基,30℃、220 r/min培养20-24 h。室温6 000 r/min离心5 min,弃上清,沉淀用100 mL的BMMY培养基悬浮,30℃、220 r/min振荡培养,甲醇诱导表达,每隔24 h添加100 %甲醇使终浓度为1.0 %,持续诱导96 h,每次添加甲醇时取2 mL的菌液,12 000 r/min离心5 min,取上清至新的离心管中 -20℃保存。按照BCA蛋白浓度定量试剂盒(北京索莱宝生物)测定表达蛋白的浓度,选择诱导72 h的上清液进行真空冷冻干燥,冻干后的蛋白于-20℃长期保存备用。
1.2.6 表达产物的SDS-PAGE和Western blot检测 将100 μL的三氯乙酸加入到1 mL的表达上清,4℃过夜。12 000 r/min离心10 min,弃上清液,加入1 mL预冷的丙酮,-20℃静止30 min,离心弃上清,室温晾干获得浓缩蛋白。向离心管中加入100 μL的2×SDS-PAGE Loading Buffer,将离心管在沸水中煮10 min,取10 μL做Tricine-SDS-PAGE,考马斯亮蓝染色,同时转印到硝酸纤维素膜上进行Western blot分析。
2 结果
2.1 △nxph1的PCR扩增
PCR扩增的产物经2 %的琼脂糖凝胶进行电泳,可以看出在750 bp附近有一特异性的DNA条带,与预期目的产物(770 bp)相符(图1)。将PCR产物连接T载体,阳性克隆再用EcoRⅠ和NotⅠ双酶切鉴定,在2 000 bp和750 bp有特异性条带(图2),分别是T载体片段和△nxph1目的基因片段,将此阳性克隆的菌液进行测序检测,确定为目的基因片段。
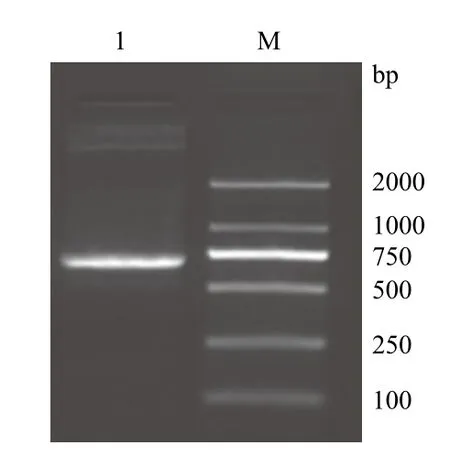
图1 △nxph1 PCR产物
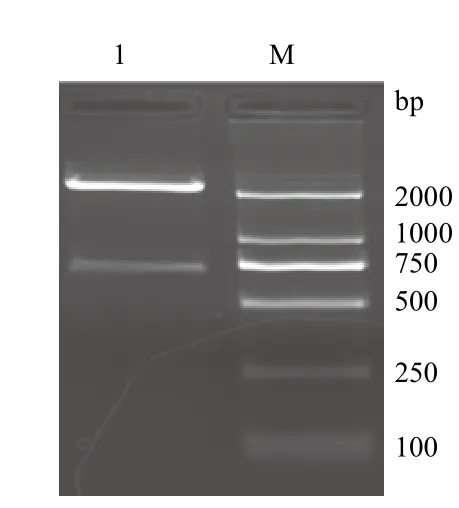
图2 △nxph1阳性克隆的双酶切
2.2 重组表达质粒的鉴定
用 EcoRⅠ和NotⅠ双酶切重组质粒pPICZαA/ nxph1 和pPICZαA/△nxph1,酶切的重组质粒pPICZαA/ nxph1应在约3 500 bp处和813 bp处有特异性的条带,酶切的重组质粒pPICZαA/△nxph1应在约3 500 bp处和750 bp处有特异性的条带,图3显示在3 500 bp处和700-1 000 bp处分别有两条带,与预期结果相符。
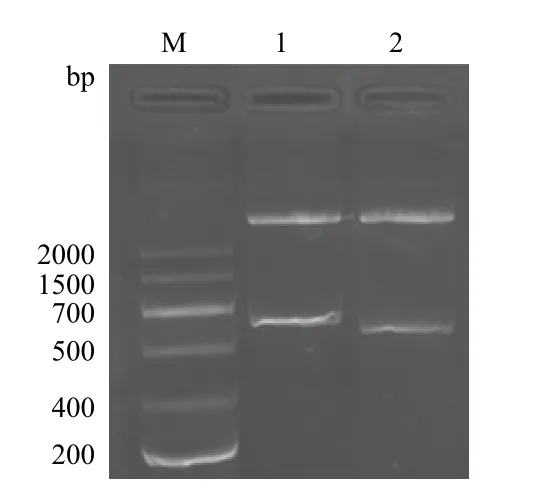
图3 重组表达质粒pPICZαA/ nxph1 和pPICZαA/nxph1的酶切图
以5'AOX和3'AOX为引物,以重组表达质粒pPICZαA/nxph1和pPICZαA/△nxph1为模板进行PCR反应,检测到扩增出的目的片段大小在1 000-1 500 bp之间(图4)。目的基因序列大小为分别为813 bp和750 bp,载体自身序列为588 bp,故扩增产物的片段大小约1 300 bp,扩增产物片段与预测片段一致,可以进一步进行测序检测。
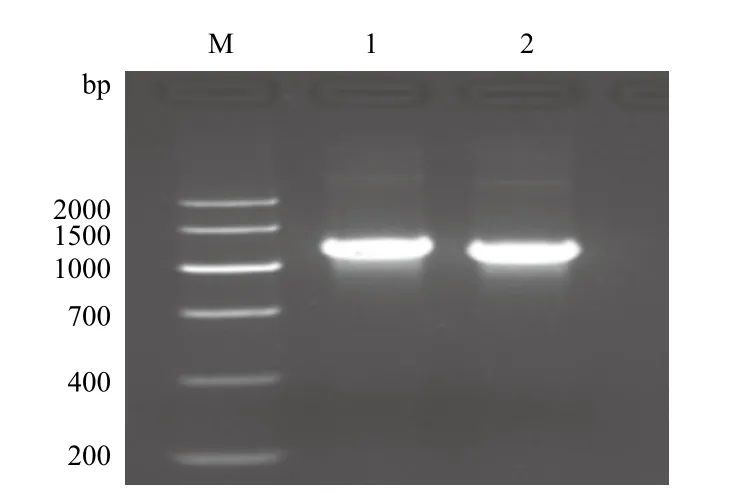
图4 重组质粒pPICZαA/ nxph1 和pPICZαA/△nxph1的PCR检测
将PCR鉴定和双酶切鉴定正确的重组质粒pPICZαA/ nxph1和pPICZαA/△nxph1送上海生工进行序列测定,测序结果证明目的基因片段已正确插入到载体pPICZαA中。
2.3 重组酵母菌的鉴定
重组表达质粒pPICZαA/ nxph1和 pPICZαA/△nxph1转入到毕赤酵母GS115后,以5'AOX和3'AOX为引物进行PCR检测,在1 000-1 500 bp处有一条特异性条带(图5),扩增产物的片段大小约1 300 bp,扩增产物片段与预测片段一致,说明目的基因片段已整合入酵母基因组。
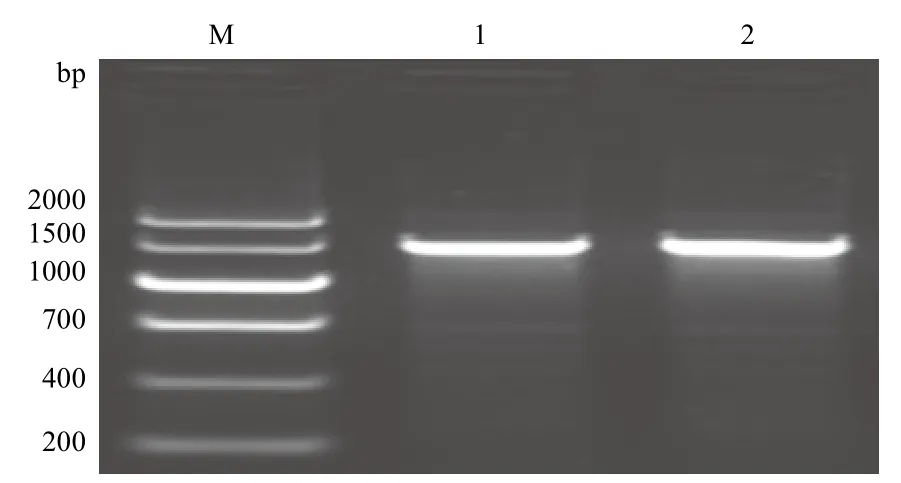
图5 pPICZαA/ nxph1 和pPICZαA/△nxph1在GS115中的PCR鉴定
2.4 Tricine-SDS-PAGE和Western blot检测结果
阳性重组菌用甲醇诱导表达,将表达产物浓缩变性后进行Tricine-SDS-PAGE电泳,用pPICZαA空酵母载体作对照,电泳完毕后考马斯亮蓝R-250进行染色,发现在约29 kD处有一条清晰的细条带,对照组没有条带。选择诱导72 h的蛋白做Western blot,结果(图6)显示在相应位置有特异的免疫性条带。
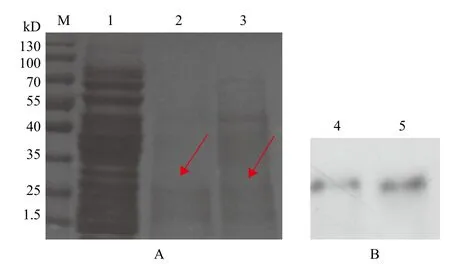
图6 重组蛋白的Tricine-SDS-PAGE和Western blot检测
2.5 蛋白浓度检测结果
蛋白标准品绘制完成蛋白标准曲线,R2=0.992,说明标准曲线较为理想,可以用于测定蛋白样品浓度。表1是分别对诱导48 h、72 h、96 h的重组蛋白pPICZαA/ nxph1和 pPICZαA/ △nxph1的浓度检测结果,从表中可以看出去除信号肽的NXPH1表达量相对更高一些,且在72 h蛋白的浓度达到最高。
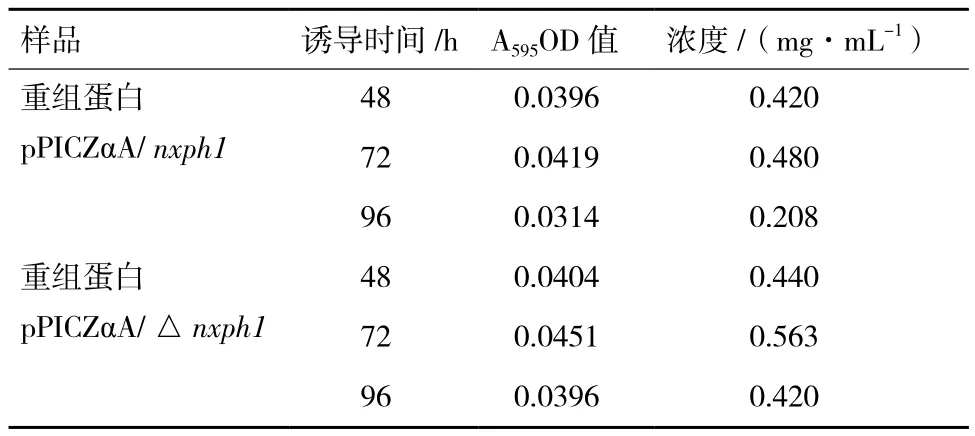
表1 蛋白浓度测定结果
3 讨论
巴斯毕赤酵母作为真核机体对产生蛋白的翻译可以进行修饰,另外有利于表达蛋白的分离纯化[5]。pPICZαA载体是含有强启动子AOX1和α-交配因子信号肽序列,能够引导外源蛋白的分泌,可以用ZeocinTM抗性基因对重组子直接进行筛选,同时载体上His标签可以进行亲和纯化[6-8]。本试验选用pPICZαA作为表达载体,携带外源基因nxph 1进入酵母表达系统并整合入酵母GS115的染色体,利用pPICZαA表达载体上的信号肽序列引导NXPH1蛋白的分泌,使NXPH1高效表达,也证实外源基因在毕赤酵母中能得到高表达。
1987年,Hoekema等报道了酵母所偏爱的25个密码子,所以在合成外源基因时可以根据毕赤酵母的密码子偏爱性,在保持翻译时氨基酸不变的情况下对密码子进行突变以获得高效表达的蛋白。2002年,Sinclair和Choy在毕赤酵母中表达人源葡糖脑苷酯酶时,通过对C+G含量的调整和密码子的优化证明其蛋白产量比天然基因提高很多倍[9]。张朝春和梁伟锋[10]根据人神经营养素(hNT-3)的基因序列,根据毕赤酵母密码子的偏爱性把编码氨基酸的密码子进行了替换,发现蛋白表达量提高11倍。Delroisse等[11]研究发现没有改造的赤拟盗羧酯酶基因在巴斯德毕赤酵母表达系统中并不表达,但是当按照酵母密码子的偏好性改造之后,可以检测到改基因的表达。Shumiao等[12]研究也发现通过优化密码子之后,可以大幅度提高瑞氏木霉纤维素内切酶基因的表达量。本研究根据GenBank上公布的鸡nxph1基因mRNA编码序列,结合毕赤酵母密码子的偏好性优化序列,将优化后的鸡nxph1全长基因序列和去除信号肽的鸡△nxph1基因序列分别定向插入酵母载体,电转化入GS115,转化后阳性菌用抗性进行筛选,甲醇诱导表达,表达的产物为NXPH1,表明我们已经通过毕赤酵母表达系统成功表达鸡NXPH1蛋白。Thill等[13]研究发现,如果在外源基因的自身信号肽引导下表达蛋白,则酵母细胞自身信号肽的表达量不高,而且分泌效率也低,因此在构建质粒时通常将外源基因的信号肽去除。龚婷等[14]对鸡白介素-2基因进行去除信号肽和全基因的诱导表达研究发现,在相同条件下去除信号肽的重组质粒的表达量高。马亮等[15]也在弓形虫表面抗原 SAG3基因的原核表达发现,不含信号肽的SAG3 基因,所构建的原核表达质粒在大肠杆菌中具有高效表达。另有研究表明,利用外源基因的信号肽引导表达的蛋白较易降解,利用酵母细胞对外源蛋白的信号肽识别后对蛋白进行传送,则表达的外源蛋白更稳定[16]。所以本试验中同时选取了去除信号肽的△nxph1基因和nxph1全基因,转化到酵母中,蛋白的表达量分别为0.563 mg/mL和0.480 mg/mL,结果也证实去除信号肽的NXPH1表达量相对更高一些。
4 结论
本研究根据GenBank上公布的鸡nxph1的序列,结合毕赤酵母码子的偏好性,成功构建了重组表达质粒pPICZαA/ nxph1和去除信号肽的重组表达质粒pPICZαA/△nxph1,并成功获得大小约29 kD鸡NXPH1蛋白和去除信号的△NXPH1蛋白,且去除信号肽的NXPH1表达量相对更高。
[1]Petrenko AG, Kovalenko VA, Shamotienko OG, et al. Isolation and properties of the alpha-latrotoxin receptor[J]. EMBO J, 1990, 9(6):2023-2027.
[2]Yang P, Wang J, Gong G, et al. Cattle mammary bioreactor generated by a novel procedure of transgenic cloning for large-scale production of functional human lactoferrin[J]. PLoS One, 2008, 3(10):e3453.
[3] 朱金金, 刘桂琼, 王志跃, 等. 鸡贮精腺差异表达基因的研究[J]. 畜牧兽医学报, 2007(11):1176-1179.
[4]Liu G, Jiang X, He C, et al. Neurexophilin 1 gene polymorphism in chickens and its variation among species[J]. Biochemical Genetics, 2013, 51(7-8):618-625.
[5]Ciarkowska A, Jakubowska A. Pichia pastoris as an expression system for recombinant protein production[J]. Postepy Biochem,2013, 59(3):315-321.
[6]董燕, 王捷, 杨连生, 等. 人骨唾液酸蛋白在巴斯德毕赤酵母中的高效分泌表达[J]. 华南理工大学学报:自然科学版, 2003(1):26-29.
[7]黄海, 谢蓓, 于瑞嵩, 等. 猪IFNα基因在毕赤酵母中的高效分泌表达[J]. 遗传, 2005, 27(2):215-220.
[8] 朱骞, 张汇东, 等. 犬白细胞介素2在毕赤酵母中的诱导表达及生物活性的测定[J]. 农业生物技术学报, 2007(4):574-578.
[9]Sinclair G, Choy FY. Synonymous codon usage bias and the expression of human glucocerebrosidase in the methylotrophic yeast,Pichia pastoris[J]. Protein Expr Purif, 2002, 26(1):96-105.
[10]张朝春, 梁伟锋, 杨希才. 神经营养素-3基因的人工合成及其在毕赤酵母中的分泌性表达[J]. 微生物学报, 2004(2):210-214.
[11] Delroisse JM, Dannau M, Gilsoul JJ, et al. Expression of a synthetic gene encoding a Tribolium castaneum carboxylesterase in Pichia pastoris[J]. Protein Expr Purif, 2005, 42(2):286-294.
[12]Shumiao Z, Huang J, Zhang C, et al. High-level expression of an Aspergillus niger endo-beta-1, 4-glucanase in Pichia pastoris through gene codon optimization and synthesis[J]. J Microbiol Biotechnol, 2010, 20(3):467-473.
[13]Thill GP, Davis GR, Stillman C, et al. Positive and negative effects of multi-copy integrated expression vectors on protein expression in Pichia pastoris[C]// In Proceedings of the 6th International Symposium on Genetics of Microorganisms. Paris:Societe Franscaise de Microbiologie, 1990, 2:477-490.
[14]龚婷, 杨孝朴, 李银聚, 等. 鸡IL-2全基因和去信号肽基因的原核表达[J]. 甘肃农业大学学报, 2009(01):11-14.
[15]马亮, 刁玉梅, 任保彦, 等. 弓形虫RH株表面抗原SAG3去信号肽基因的蛋白原核表达及鉴定[J]. 吉林农业大学学报,2012, 34(1):104-108.
[16]Ferrarese L, Trainotti L, Gattolin S, et al. Secretion, purification and activity of two recombinant pepper endo-beta-1, 4-glucanases expressed in the yeast Pichia pastoris[J]. FEBS Lett, 1998, 422(1):23-26.
(责任编辑 李楠)
Expression of Chicken Neurexophilin 1 Gene in Pichia pastoris
Chen Meiling Nie Bin Jiang Xunping Liu Guiqiong
(College of Animal Science and Technology, Huazhong Agricultural University, Wuhan 430070)
Neurexophilin 1 is the differentially expressed gene at the chicken utero-vaginal junction. According to the mRNA sequence of chicken nxph1 and codon bias of Pichia pastoris, the modified DNA sequence for chicken nxph1 which encodes the same amino acid sequence as that of chicken nxph1were synthesized. Specific primers were designed to amplify part of the synthesized DNA sequence whose expression product has no signal peptide. The two sequences were inserted into pPICZαA vector to construct the recombinant pPICZαA/ nxph1 and pPICZαA/△ nxph1(without signal peptide), and then the recombinant plasmids were transfected into Pichia pastoris GS115 by electroporation. The positive recombinant strains were inducted to express protein by addition of 1 % methanol. The expression protein was displayed on the Tricine-SDS- PAGE electrophoresis and Western blot. The result showed that chicken NXPH1 protein and △NXPH1 protein were successfully expressed, and the molecular weight of the protein was approximately 29 kD, which provided foundations for analyzing functions of NXPH1 in chicken reproductive tract.
nxph 1;Pichia pastoris;gene expression
10.13560/j.cnki.biotech.bull.1985.2015.04.030
2014-07-07
国家自然科学基金项目(31071088)
陈美玲,女,硕士研究生,研究方向:动物遗传育种与繁殖;E-mail:chenmeiling064@126.com
刘桂琼,副教授,E-mail:liuguiqiong@mail.hzau.edu.cn
猜你喜欢
生物学通报(2020年11期)2020-10-22
中成药(2018年7期)2018-08-04
食品与发酵工业(2018年3期)2018-04-12
浙江农业学报(2017年3期)2017-04-08
广东饲料(2016年1期)2016-12-01
中国烟草学报(2016年1期)2016-11-16
工业微生物(2016年5期)2016-11-11
化学与生物工程(2016年10期)2016-11-10
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
茶叶通讯(2014年4期)2014-02-27
