
青藏高原青稞耐寒种质资源基于SSR标记的遗传多样性及群体结构分析
2017-03-01 07:59巴桑玉珍刘新春付国勇李东梅王丹丹强小林冯宗云
麦类作物学报 2017年1期
巴桑玉珍,刘新春,付国勇,李东梅,王丹丹,强小林,冯宗云
(1.四川农业大学农学院作物遗传育种学系大麦青稞研究中心,四川成都 611130;2.西藏自治区农牧科学院农业研究所,西藏拉萨 850032)
青藏高原青稞耐寒种质资源基于SSR标记的遗传多样性及群体结构分析
巴桑玉珍1,2,刘新春1,付国勇1,李东梅1,王丹丹1,强小林2,冯宗云1
(1.四川农业大学农学院作物遗传育种学系大麦青稞研究中心,四川成都 611130;2.西藏自治区农牧科学院农业研究所,西藏拉萨 850032)
为了解青藏高原青稞耐寒种质资源的遗传基础,利用SSR标记分析了来自青藏高原地区的71份青稞耐寒育种资源的遗传多样性和群体遗传结构。结果表明,利用从分布于大麦7个连锁群上的200对SSR引物中筛选的48对多态性引物,共检测到230个等位条带,变化范围为1~10个,平均每对SSR引物可检测到4.79个条带。多态性信息含量(PIC)变化范围为0.054 7~0.856 9,平均值为0.489 8。遗传相似系数(GS)的变化范围为0.469~0.924,平均值为0.745。经聚类分析,在GS约0.740处可将71份材料分为五大类,第51、43和14号品种分别聚为A、B和C类,第22、27、39和50号品种聚为D类,其余64个品种聚为E类,其中,第51号品种的穗长(3.8 cm)、穗粒数(44.2粒)最低,第43号的单穗小穗数(90个)最高,但千粒重(35.6 g)最低,第14号品种的生育期(86 d)最短,但穗粒数(69.6粒)最多。此外,群体遗传结构分析表明,71份青稞资源材料可划分为2个亚群。
青稞;遗传多样性;SSR;群体结构;聚类分析
青稞(Hordeumvulgarevar.nudumHooker f.)即裸大麦(hulless barley),在我国境内主要生长在素有“世界屋脊”之称的青藏高原及辐射边缘区域,是这些区域主要的粮食作物[1-3]。青藏高原的青稞种质资源极其丰富,耐寒性各异。在青藏高原地区,低温伤害是影响生产情况的重要限定因素[4],青稞耐寒性种质资源筛选评价及耐寒青稞品种选育日益受到青稞育种者的高度关注。研究青藏高原耐寒育种资源的遗传多样性及群体遗传结构,对于了解青藏高原的这些种质资源的遗传基础及进一步指导青稞耐寒育种工作具有重要的意义。
SSR即简单序列重复(simple sequence repeats),又称为微卫星(microsatellites)标记。自1994年Saghai Maroof等[5]利用4对SSR引物研究了世界大麦主产区的栽培大麦及以色列野生二棱大麦的SSR多态性以来,该标记因其具有使用方便、费用低及重复性好等优点使其在大麦研究中得到较广泛地应用[6]。在我国,冯宗云等[7-9]最早利用SSR标记分析了西藏野生大麦和青藏高原青稞地方品种的遗传多样性。随后,一些研究者也利用该标记对我国不同来源的青稞地方品种、选育品种等开展了遗传多样性研究,但并不多见[10-15]。在群体遗传结构分析上,利用SSR标记分析大麦地方品种、育成品种、野生大麦及国外引进材料等的群体遗传结构的研究还不少[14,16-20],但利用该标记分析青稞群体遗传结构的研究却少见[14-15]。而对于耐寒青稞种质而言,至今尚未见利用SSR标记揭示其遗传多样性及遗传结构的报道。
鉴于此,本研究以青藏高原耐寒性各异的青稞农家品种及育成品种为材料,利用48对SSR分子标记对其遗传多样性及群体遗传结构进行研究,以期了解这些材料的遗传基础,为青稞耐寒基因资源的挖掘及耐寒育种资源的高效利用提供依据。
1 材料与方法
1.1 材 料
71份青藏高原青稞种质材料中,62份来自西藏、4份来自青海、3份来自四川、2份来自甘肃,除编号为1、3~6、10~14、24~25、70~71的材料为育成品种外,其余57份材料均为农家品种,这些材料的耐寒性差异较大(表1)。所有材料现均保存于四川农业大学农学院大麦青稞研究中心。
1.2 基因组DNA的提取
在PGX型多段可编程光照培养箱(宁波东南仪器设备有限公司)中,于温度为25 ℃、日光照时间为8 h条件下,培养青稞材料至三叶一心期时,每份青稞材料取新鲜健康的幼叶约3~5 g,参照CTAB法[21]提取基因组DNA,用1%琼脂糖凝胶电泳检测DNA质量,用紫外分光光度计测定DNA浓度,于-20 ℃保存备用。
1.3 SSR分析
1.3.1 SSR引物的选择
选用覆盖大麦7个连锁群的200对SSR标记[22](由上海英俊生物有限公司合成),对不耐寒的青稞选育品种“藏青320”和高度抗寒的青稞地方品种“理塘勾芒”进行SSR标记筛选,根据扩增效果、扩增稳定性及在染色体上的分布原则在1H、2H、3H、4H、5H、6H和7H染色体上分别选择了3、8、3、9、10、6、9对共48对SSR标记(表2),对71份青稞材料进行遗传多样性及遗传结构分析。
1.3.2 PCR扩增
PCR在BIO-RAD扩增仪上进行。PCR反应试剂均购自TaKaRa公司。PCR总反应体系为20 μL,DNA模板(50 ng·μL-1)2.0 μL,dNTPs(2.5 mmol·L-1)1.6 μL,10×Buffer 2.0 μL,MgCl2(25 mmol·L-1)1.2 μL,rTaq聚合酶(2.5 U·μL-1)0.4 μL,正向和反向引物(10 μmol·L-1)各0.5 μL,补超纯水至总体积20 μL。反应程序: 95 ℃预变性5 min;95 ℃变性45 s,55 ℃退火45 s,72 ℃延伸1 min,35个循环;72 ℃延伸10 min,4 ℃保存。
1.3.3 扩增产物的检测
扩增产物加3×Loading Buffer 10 μL,95 ℃变性5 min后,用预热30 min的6%变性聚丙烯酰胺凝胶恒功率(75 W)电泳,至指示剂离胶板底部三分之一处(约75 min),取下胶板。采用硝酸银染法检测[22],用数码相机拍照,统计带型。
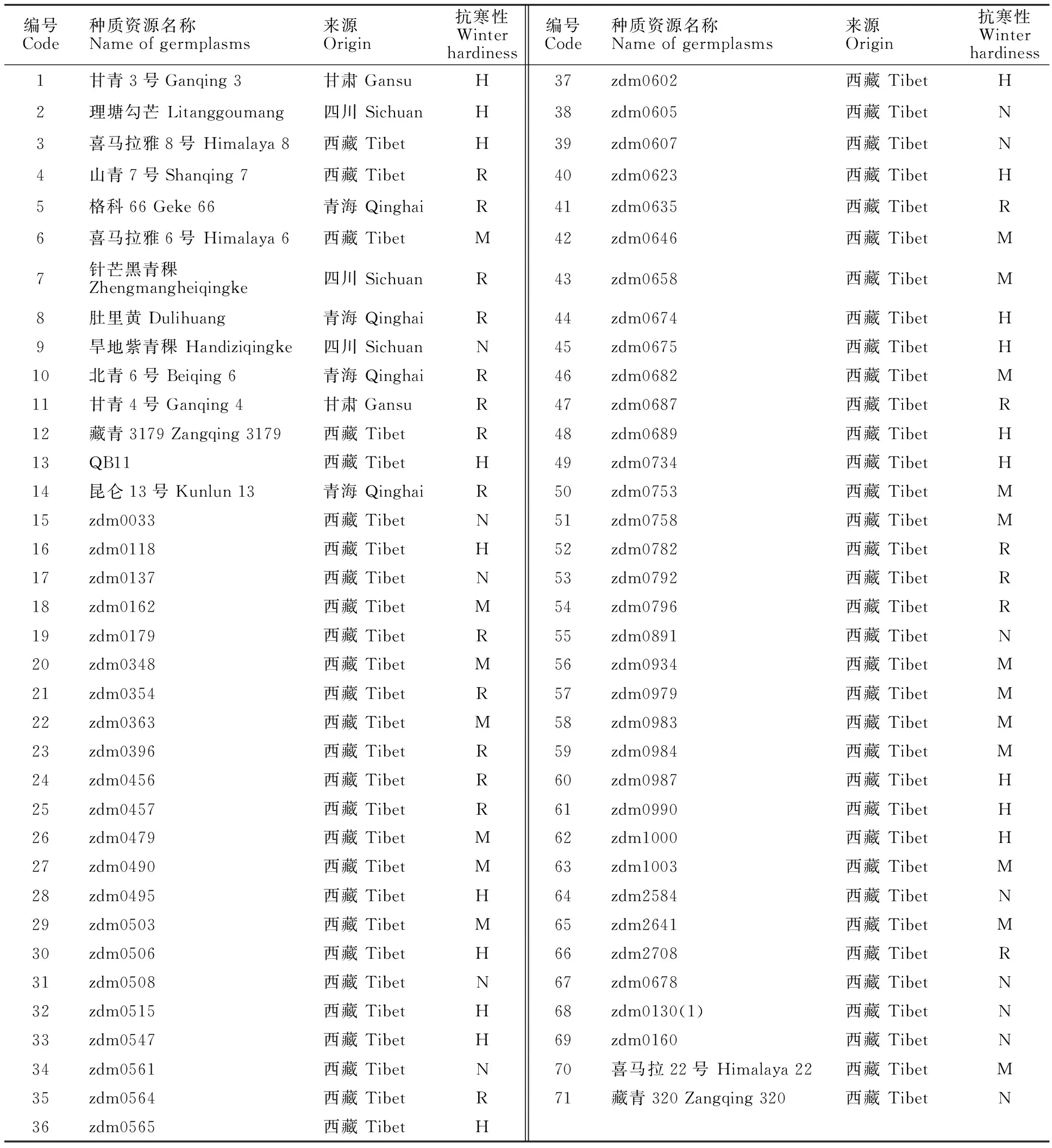
表1 供试的71份青稞种质资源Table 1 Hulless barley materials used in this study
H:高度抗寒;R:抗寒;M:中度抗寒;N:不抗寒。
H: Highly winter hardiness; R: Resistance; M: Medium winter hardiness; N: No winter hardiness.
1.4 数据分析
1.4.1 遗传多样性分析

1.4.2 群体结构分析
利用基于混合模型的Structure 2.3软件[24-25]分析青稞资源的群体结构。设定群体数K的估计值范围为1~11,将MCMC(Markov chain montecarlo)开始时的不作数迭代(length of burn-in period)设为 10 000次,将不作数迭代后的MCMC设为100 000次,每个K值重复数为20次。以似然值最大为原则确定最优群体数K的值,计算各材料的Q值,分析群体结构。
2 结果与分析
2.1 SSR标记的多态性
从表2可知,48对SSR引物在71份供试材料中共检测出230个条带,每对SSR引物可扩增出1~10个条带,平均每对SSR引物可扩增4.79个条带。其中,位于第二染色体的GBM1459检测到的条带数最多,为10个;分别位于第一和第二染色体的GMS021和Bmag0125检测到的条带数次之,为9个;而位于第五染色体上的GBM1176、GBM1438和位于第六染色体上的Bmag0173、Bmag0500检测到的条带数最少,仅为1个。此外,48对SSR引物在供试材料间的PIC值不同,其中位于第五染色体上的SSR标记Bmag0337的PIC值最高,为0.856 9,位于第一染色体上的SSR标记GBM1234的PIC值最低,为0.054 7,平均每对SSR引物的PIC值为0.489 8。
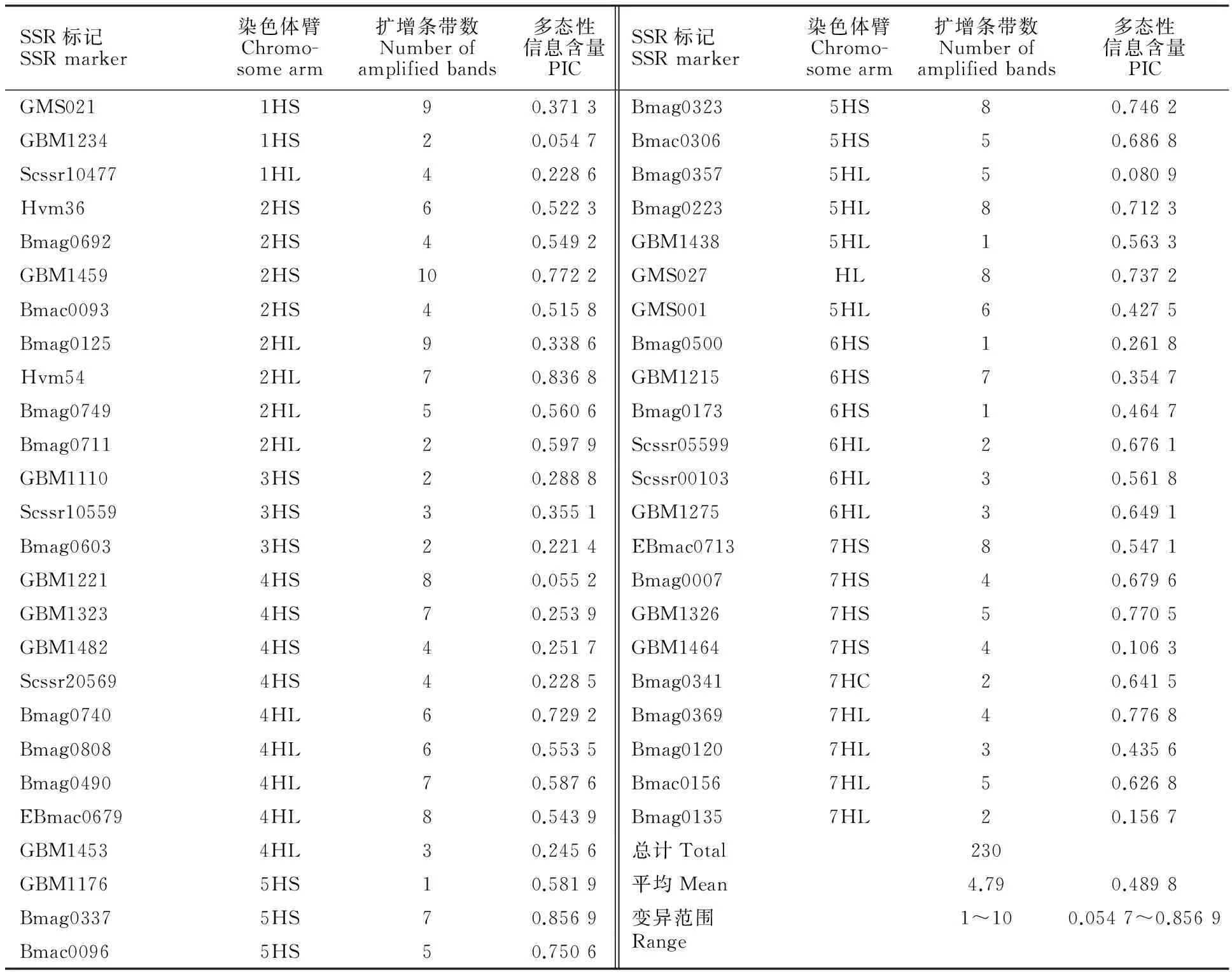
表2 用于本研究的SSR标记的名称、所在染色体臂、扩增条带数及多态性信息含量Table 2 Name,chromosome arm and amplified bands as well as polymorphic information content (PIC) for SSR markers used in this study
2.2 SSR标记揭示的品种间遗传相似系数
根据SSR标记数据,利用软件NTSYSpc 2.10计算遗传相似系数,其变化范围为0.469~0.924,平均值为0.745。其中,第 51号品种与第6号品种和第13号品种的遗传相似系数最低(0.469),说明二者之间的亲缘关系最远,适宜进行相互配组;而第12号品种与第10号品种的遗传相似系数最高(0.924),说明二者之间的亲缘关系最近,育种中不宜作亲本相互杂交。
2.3 群体结构
利用Structure 2.3软件分析了71份青稞种质的群体遗传结构。设定类群数K值在1~11之间,以每个K值为横坐标,以相应LnP(D)值为纵坐标作曲线图(图1)。从图1可知,LnP(D)值随着亚群数的增加而增加,曲线无明显拐点,无法确定适宜的亚群数。因此,参考Evanno等[26]的方法对71份供试青稞材料进行基于数学模型的群体结构分析,根据K所对应的最大似然值来选择合适的群体数目,用ΔK来确定合适的K值(图2),从图2看出,ΔK在K=2和K=4时曲线均具有拐点,但当K=2时,ΔK有最明显的峰值,各群体内青稞材料相似性最大。故根据似然值最大的原则选定K值为2为最终的群体数目。因此,71份青稞材料群体被分为2个亚群,群体结构分析结果如图3所示。第1个亚群包含第22、39、43、48、49、50和51号七份材料,全部为地方品种,其中,2份材料具高抗寒性,4份材料具中度抗寒性,1份材料不具抗寒性;而第二亚群中有13份材料为育成品种(占22.03%),其余为地方品种(占77.97%),各类型抗寒材料均存在,高抗寒、抗寒、中度抗寒和不抗寒材料分别占28.8%、32.3%、22.0%和17.0%。根据各材料的最大Q值分布看,71份材料中Q<0.6的有5份材料,即第14、27、38、58和67号品种,占7.0%,认为其有混合来源,表示其亲缘关系较复杂,而Q≥0.6的有66份材料,占93.0%,认为这些材料亲缘关系较为单一。

图1 K值与LnP(D)值的关系
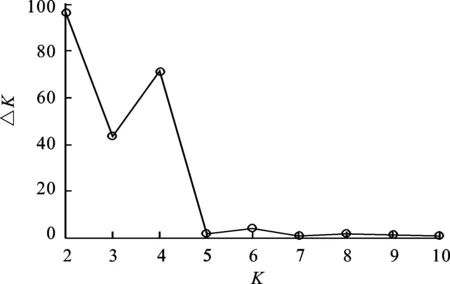
图2 ΔK值随K值的变化
2.4 聚类分析
从图4可知,在遗传相似系数约为0.740处,可以把参试的71个品种分为五大类,第51、43和14号品种各自单独聚为一类,分别为A、B和C类群,第22、27、39和50号品种聚为D类群,而其余64个品种聚为另一大类,为E类群。结合2013年田间农艺性状调查结果可知,A类群的第51号品种的穗长、穗粒数最低,分别为3.8 cm、44.2粒;B类群的第43号品种的小穗数最高(90个),而千粒重最低(35.6 g);C类群的第14号品种的生育期最短(86 d)、穗粒数最多(69.6粒);D类群的4个品种的株高的平均值最高(100.75 cm);E类群的千粒重的平均值最高(45.24 g)。在遗传相似系数约为0.748处,可将E类群分为两个亚类,其中第1、3、5和6号品种为一亚类(E1),其余聚为另一亚类(E2)。E1亚类的4个品种全为选育品种,其株高的平均值最低(75.25 cm)。
根据上述群体遗传结构分析结果,D类群中的4份材料有3份材料(第22、39和50号品种)在第一亚群,占75.0%,E类群中的64份材料中除第38、48、49、58和67号品种外有59份材料归入第二亚群,占92.2%。这表明聚类分析结果与群体结构分析结果存在一定的关联性。
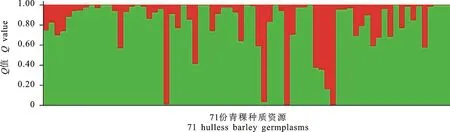
图3 参试青稞种质资源的群体遗传结构

图4 71份青稞种质资源的聚类图
3 讨 论
本研究先以两个生态差异较大且耐寒性有明显差异的青稞品种“藏青320”(不耐寒,育成品种)和“理塘勾芒”(高度耐寒,地方品种)为材料,对选自大麦7个连锁群上的200对SSR标记[22]进行了多态性引物筛选,共筛选出分别来自染色体1H-7H上的48对多态性SSR引物。然后以这48对SSR引物对71份青藏高原区域的青稞品种进行了遗传多样性分析,共检测到230个等位条带,变化范围为1~10个,以位于染色体5H上的SSR标记GBM1176、GBM1438和位于染色体6H上的SSR标记Bmag0173、Bmag0500检测到的等位条带数最少(仅1个),而染色体2H上的GBM1459检测到的等位条带数最多(达10个),平均每对SSR引物检测到4.79个等位条带。其多态性信息含量(PIC)变化范围为0.054 7~0.856 9,平均值为0.489 8,以来自染色体1H上的SSR标记GMM1234的PIC值最低(0.054 7),以染色体5H上的SSR标记Bmag0337的PIC值最高(0.856 9)。而Feng等[8]以来自大麦7个连锁群上的30对SSR标记研究了106份西藏野生大麦的遗传多样性,等位条带变化范围为1~14个,平均每对标记检测到7.6个,其平均PIC值为0.572 3。这表明西藏野生大麦的遗传变异较青藏高原的栽培青稞丰富。同样,Feng等[9]利用来自大麦7个连锁群上的35对SSR标记研究了来自西藏、青海及四川高原的65份青稞地方品种的遗传多样性,等位条带变化范围为2~16个,平均每对SSR标记检测到7.09个,材料间遗传基础较窄。孟亚雄等[15]用48对SSR标记研究了来自西藏、四川、青海、甘肃、北京、内蒙古的108份青稞地方品种和选育品种的遗传多样性,等位条带变化范围为2~6个,平均每对标记3.2个,认为材料间的亲缘关系较近。而潘志芬等[10]和吴昆仑[13]利用SSR标记分别研究了64份、55份青藏高原栽培青稞的遗传多样性,认为青藏高原栽培青稞具有较丰富的多样性。这可能与材料及SSR引物的选择有关,二者所选材料均含地方品种、育种品种,但潘志芬等[10]所用材料仅来自西藏和四川,而吴昆仑[13]所用材料除来自西藏、四川外,还包含青海、云南的材料;从SSR引物选择上,二者所选SSR引物均来自大麦7个连锁群,但潘志芬等[10]所选SSR标记有5对与性状未建立联系,其余15对分别与大麦生理、抗逆、抗病及品质性状相关,而吴昆仑[13]仅选择了14对SSR引物。前人认为,标记的正确选择对于种质资源的遗传多样性研究尤为重要[6, 27]。进一步根据遗传相似系数(GS)对71份青稞材料进行聚类分析,结果表明,在GS值约为0.740处,可将71份青稞材料聚为五大类,第51、43和14号品种各自单独为一类,分别为A、B和C类,第22、27、39和50号品种聚在D类群,而其余64个品种聚为一大类(E类),同样说明青藏高原地区的大部分青稞材料亲缘关系较近,其遗传基础狭窄。该结果与本实验室袁金娥[14]于2012年采用与大麦耐盐性相关的27对SSR标记对148份来自青藏高原区域的青稞材料进行多样性研究后所得结果一致。而杨 平等[28]利用SRAP(sequence-related amplified polymorphism)标记分析后得出四川高原青稞育成品种的遗传基础较为狭窄。同样,群体遗传结构分析显示,供试的71份材料仅可分为2个类群,也说明供试材料的遗传基础较窄。因此,青藏高原地区青稞耐寒育种应加强外来抗寒种质资源的引进,特别是野生大麦(包括裸大麦)材料及国外材料,丰富该区域的种质资源,拓展其遗传基础,避免遗传脆弱性,也是青稞育种者及青稞种质资源研究者的当务之急。
[1] 徐廷文.青稞穗原始体分化期及其营养条件问题的初步探讨[J].中国农业科学,1963,4(2):48.
XU T W.The preliminary discussion on the issues of spike primitive differentiation stage and nutritional condition in hulless barley [J].ScientiaAgriculturalSinica,1963,4(2):48.
[2] 巴桑玉珍,强小林.西藏青稞育种的成就与经验分析[J].西藏农业科技,2004,27(1):26.
BASANG Y Z,QIANG X L.Analysis on the achievement and experiences of hulless barley breeding in Tibet [J].TibetanAgriculturalScienceandTechnology,2004,27(1):26.
[3]ZENG X,LONG H,WANG Z,etal.The draft genome of Tibetan hulless barley reveals adaptive patterns to the high stressful Tibetan plateau [J].ProceedingsoftheNationalAcademyoftheSciencesoftheUnitedStatesofAmerica,2015,112(4):1095.
[4] 段瑞君,任有成,熊辉岩.低温胁迫对青稞幼苗抗寒性生理指标的影响[J].安徽农业科学,2009,37(34):16796.
DUAN R J,REN Y C,XIONG H Y.Effects of low temperature stress on chill resistance physiological indexes of highland barley seedlings [J].JournalofAnhuiAgriculturalSciences,2009,37(34):16796.
[5]SAGHAI MAROOF M A,BIYASHEV R M,YANG G P,etal.Extraordinarily polymorphic microsatellite DNA in barley: Species diversity,chromosomal locations and population dynamics [J].ProceedingsoftheNationalAcademyoftheSciencesoftheUnitedStatesofAmerica,1994,91(12):5466.
[6] 冯宗云,张义正,凌宏清.大麦基因组中的微卫星标记及其应用[J].遗传,2002,24(6):727.
FENG Z Y,ZHANG Y Z,LING H Q.Microsatellite markers and application in the barley genome [J].Hereditas,2002,24(6):727.
[7] 冯宗云,张义正,张立立,等.应用微卫星标记研究西藏野生二棱大麦的遗传多样性及地理分化[J].高技术通讯,2003,13(10):46.
FENG Z Y,ZHANG Y Z,ZHANG L L,etal.Genetic diversity and geographical differentiation ofHordeumvulgaressp.spontaneumin Tibet using microsatellite markers [J].HighTechnologyLetters,2003,13(10):46.
[8]FENG Z Y,LIU X J,ZHANG Y Z,etal.Genetic diversity analysis of Tibetan wild barley using SSR markers [J].ActaGeneticaSinica,2006,33(10):917.
[9]FENG Z Y,ZHANG L L,ZHANG Y Z,etal.Genetic diversity and geographical differentiation of cultivated six-rowed naked barley landraces from the Qinghai-Tibetan plateau of China detected by SSR analysis [J].GeneticsandMolecularBiology,2006,29(2):330.
[10] 潘志芬,邹弈星,邓光兵,等.青藏高原栽培青稞SSR标记遗传多样性研究[J].中山大学学报(自然科学版),2007,46(2):82.
PAN Z F,ZHOU Y X,DENG G B,etal.Genetic diversity of SSR markers in cultivated hulless barley from Qinghai-Tibet plateau in China [J].ActaScientiarumNaturaliumUniversitatisSunyatseni,2007,46(2):82.
[11] 张建华,杨晓洪,于亚雄,等.云南青稞(裸大麦)品种亲缘关系的SSR标记研究[J].麦类作物学报,2009,29(1):35.
ZHANG J H,YANG X H,YU Y X,etal.Study on genetic relationship of Yunnan naked barley by SSR markers [J].JournalofTriticeaeCrops,2009,29(1):35.
[12] 杨 菁,迟德钊,吴昆仑,等.青海省栽培青稞SSR标记遗传多样性研究[J].安徽农业科学,2010,38(8):4307.
YANG Q,CHI D Z,WU K L,etal.Genetic diversity of SSR markers in cultivatedHordeumvulgareL.in Qinghai province [J].JournalofAnhuiAgriculturalSciences,2010,38(8):4307.
[13] 吴昆仑.青稞种质资源的SSR标记遗传多样性分析[J].麦类作物学报,2011,31(6):1030.
WU K L.Genetic diversity analysis of hulless barley germplasm by SSR markers [J].JournalofTriticeaeCrops,2011,31(6):1030.
[14] 袁金娥.中国近缘野生大麦和栽培青稞的耐盐性鉴定及其与SSR标记的关联分析[D].雅安:四川农业大学,2012:56.
YUAN J E.Evaluation of salt tolerance and their association analysis with SSR markers in barley wild relatives and cultivated qingke(hulless barley) of China [D].Ya’an:Sichuan Agricultural University,2012:56.
[15] 孟亚雄,孟祎林,汪军成,等. 青稞遗传多样性及其农艺性状与SSR标记的关联分析[J].作物学报,2016,42(2):180.
MENG Y X,MENG W L,WANG J C,etal.Genetic diversity and associate analysis of agronomic characteristics with SSR markers in hulless barley [J].ActaAgronomicaSinica,2016,42(2):180.
[16] 何智宏,司二静,赖 勇,等.大麦亲本材料SSR标记遗传多样性及群体结构分析[J].麦类作物学报,2013,33(5):894.
HE Z H,SI E J,LAI Y,etal.Genetic diversity and population structure in barley parent materials using SSR markers [J].JournalofTriticeaeCrops,2013,33(5):894.
[17] 赖 勇,孟亚雄,王 晋,等.大麦遗传多样性及连锁不平衡分析[J].作物学报,2013,39(12):2154.
LAI Y,MENG Y X,WANG J,etal.Genetic diversity and linkage disequilibrium analysis in barley [J].ActaAgronomicaSinica,2013,39(12):2154.
[18] 王 晋,王世红,赖 勇,等.大麦SSR标记遗传多样性及群体遗传结构分析[J].核农学报,2014,28(2):177.
WANG J,WANG S H,LAI Y,etal.Genetic diversity and population structure analysis by using SSR markers in barley [J].JournalofNuclearAgriculturalSciences,2014,28(2):177.
[19] 司二静,赖 勇,孟亚雄,等.大麦遗传多样性及SSR标记与大麦条纹病抗性关联分析[J].农业生物技术学报,2015,23(2):193.
SI E J,LAI Y,MENG Y X,etal.Genetic diversity and association analysis of SSR markers with leaf stripe resistance in barley(Hordeumvulgare) [J].JournalofAgriculturalBiotechnology,2015,23(2):193.
[20] 张 宇,司二静,孟亚雄,等.不同来源大麦材料的农艺性状和籽粒蛋白质含量及群体遗传结构分析[J].麦类作物学报,2015,35(7):940.
ZHANG Y,SI E J,MENG Y X,etal.Analysis on agronomic traits,grain protein content and population structure of barley germplasm from different geographical origins [J].JournalofTriticeaeCrops,2015,35(7):940.
[21]GUO L L,LIU X J,LIU X C,etal.Construction of genetic map in barley using sequence-related amplified polymorphism markers,a new molecular marker technique[J].AfricanJournalofBiotechnology,2012,11(74):13859.
[22] 刘仙俊.大麦特异种质资源的分子生物学研究[D].雅安:四川农业大学,2011:94.
LIU X J.Study on molecular biology of special germplasm resources in barley [D].Ya'an:Sichuan Agricultural University,2011:94.
[23]ANDERSON J A,CHURCHILL G A,AUTRIQUE J E,etal.Optimizing parental selection for genetic linkage maps [J].Genome,1993,36(1):181.
[24]PRITCHARD J K,STEPHENS M,DONNELLY P.Inference of population structure using multilocus genotype data [J].Genetics,2000,155(2):945.
[25]FALUSH D,STEPHENS M,PRITCHARD J K.Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies [J].Genetics,2003,164(4):1567.
[26]EVANNO G,REGNAUT S,GOUDET J.Detecting the number of clusters of individuals using the software structure: a simulation study [J].MolecularEcology,2005,14(8):2611.
[27] 徐如虹,任明见,杨英苍,等.贵州省区试小麦品系遗传多样性的SSR标记研究[J].山地农业生物学报,2005,24(1):12.
XU R H,REN M J,YANG Y C,etal.Analysis on genetic diversity among Guizhou wheat lines based on SSR markers [J].JournalofMountainAgricultureandBiology,2005,24(1):12.
[28] 杨 平,刘仙俊,刘新春,等.利用SRAP标记研究四川高原青稞育成品种的遗传多样性[J].遗传,2008,30(1):115.
YANG P,LIU X J,LIU X C,etal.Genetic diversity analysis of the developed qingke (hulless barley) varieties from the plateau regions of Sichuan province in China revealed by SRAP markers [J].Hereditas,2008,30(1):115.
Genetic Diversity and Population Structure Analysis of Hulless Barley with Cold Tolerance from the Qinghai-Tibetan Plateau Revealed by SSR Markers
BASANG Yuzhen1,2,LIU Xinchun1,FU Guoyong1,LI Dongmei1, WANG Dandan1,QIANG Xiaolin2,FENG Zongyun1
(1.Barley and Hulless Barley Research Centre,College of Agronomy,Sichuan Agricultural University,Sichuan,Chengdu 611130,China; 2.Agricultural Research Institute,Tibet Agriculture and Animal Husbandy Sciences,Lhasa,Tibet 850032,China)
In order to understand the genetic basis of hulless barley with cold tolerance from the Qinghai-Tibetan Plateau,genetic diversity and population genetic structure of 71 hulless barley germplasm resources from the Qinghai-Tibetan Plateau were analyzed using 48 SSR markers with better polymorphic bands,which were selected from 200 pairs of SSR primers located on the seven linkage groups of barley.The results showed that a total of 230 allelic bands were identified in 71 hulless barley materials,varying from 1 to 10 with the average alleles per SSR locus of 4.79.The variation range of genetic similarity coefficients was 0.469~0.924 with the average of 0.745.The polymorphic information content (PIC) ranged from 0.054 7 to 0.856 9 with the mean of 0.489 8.These hulless barley materials could be clustered into two subgroups according to population genetic structure analysis.Cluster analysis based on UPGMA (Unweight pair group method using arithmetic averages) method indicated that 71 hulless barley accessions could be divided into five major groups at the genetic similarity coefficient (GS) level of 0.740,and the cultivar No.51,No.43 and No.14 were clustered into the Group A,B and C,respectively,and the cultivar No.22,No.27 and No.39 as well as No.50 were clustered as Group D,and the others as Group E including 64 hulless barley accessions.The cultivar No.51 from Group A had the lowest spike length (3.8 cm) and grain number per spike (44.2),and the No.43 from Group B had the highest spikelet number per spike (90) and lowest 1 000-grain weight (35.6 g),but the No.14 showed the shortest growth period (86 d) and maximum grain number per spike (69.6).The analysis based on population genetic structure and cluster analysis unanimously revealed that the genetic basis existed in most of hulless barley materials used in this study was narrow.
Hulless barley; Genetic diversity; SSR; Population structure; Cluster analysis
时间:2017-01-03
2016-08-09
2016-11-28
国家现代农业产业技术体系(大麦青稞)建设专项(CARS-05)
E-mail:36165329@qq.com
冯宗云(E-mail:zyfeng49@126.com)
S512.1;S332
A
1009-1041(2017)01-0041-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20170103.1625.012.html
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
自然灾害学报(2022年2期)2022-05-10
星星·诗歌原创(2022年1期)2022-04-16
环境卫生工程(2021年4期)2021-10-13
中国学校体育(2021年10期)2021-04-26
疯狂英语·新读写(2021年2期)2021-02-25
西藏人文地理(2020年4期)2020-11-19
翠苑(2019年4期)2019-11-11
华人时刊(2016年19期)2016-04-05
中国光学(2015年1期)2015-06-06
