
青稞主要性状的差异性与遗传分析
2017-03-01 07:59孟亚雄马增科任盼荣司二静汪军成张海娟马小乐王育才王化俊
麦类作物学报 2017年1期
孟亚雄,马增科,任盼荣,司二静,汪军成,杨 轲,张海娟,马小乐,王育才,王化俊
(1.甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室,甘肃兰州 730070;2.甘肃农业大学农学院,甘肃兰州 730070)
青稞主要性状的差异性与遗传分析
孟亚雄1,2,马增科1,2,任盼荣1,2,司二静1,2,汪军成2,杨 轲1,2,张海娟1,2,马小乐1,2,王育才1,2,王化俊1,2
(1.甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室,甘肃兰州 730070;2.甘肃农业大学农学院,甘肃兰州 730070)
为合理评价110份青稞品种(系)的农艺性状及遗传特性,对参试种质材料的株高、穗长、穗粒数、分蘖数和千粒重进行了调查统计,同时用分布于大麦全基因组的48对SSR标记进行了遗传多样性分析。结果表明,参试材料各农艺性状变异较大,变异系数为2.92%~42.33%,其中株高、穗长、分蘖数、穗粒数和千粒重变化范围分别为53~122 cm、2.5~7 cm、2~19个、20~90粒和25.46~60.34 g。SSR标记共检测到168个等位变异,变化范围为2~6个。110份青稞种质材料分成三大类群。SSR标记的1 128个位点成对组合中,不论是共线性成对组合还是非共线性成对组合,都存在一定的连锁不平衡(linkage disequilibrium,LD)。D′统计概率(P<0.01)支持的LD成对位点438个,占全部位点的38.8%,D′的平均值为0.36,整体LD水平较高。较高水平的LD位点(D′>0.5)主要分布在2H、4H、6H和7H连锁群上。
青稞;农艺性状;SSR标记;连锁不平衡
青稞是生长在青藏高原地区的裸大麦,是我国藏族同胞的主食,也是用来酿造青稞酒的主要原料,具有较高的营养价值和保健功效。近年来,随着对青稞的营养价值和保健功效的深入研究,青稞保健品及食品加工产业也得到了不断发展。青稞除了籽粒外,其秸秆也是青藏高原牧区的主要冬饲草,因此青稞生产对我国藏民的生活有着重要的意义。青藏高原地形复杂多样,种植栽培的青稞种质资源丰富多样,拥有大量可被利用和研究的基因资源[1]。和其他作物一样,现代青稞育种亦是以高产、优质和抗逆为主要育种目标,造成基因流失的现象越来越明显。因此,研究青稞种质材料的遗传多样性,发掘优良资源和有利基因,拓宽种质资源将成为解决该问题的主要途径。
目前,国内外学者已采用多种方式对作物种质资源的遗传多样性进行研究。其中,Dotlacil等[2]和Delacy等[3]认为,作物的形态学标记(农艺性状)比较直观,而且便于识别和易于掌握,且与生产实践直接相关,是一种颇为有效的作物遗传多样性分析的方法。其中,作物的株高、穗粒数、千粒重、分蘖数和穗长等农艺性状是作物育种中要重点考察的对象。另外,随着现代分子标记技术的发展和应用,为作物种质资源研究和有利基因挖掘提供了条件。其中,SSR标记具有重复性好、多态性高等优点,已被广泛应用于作物遗传多样性和基因发掘等方面[4-7]。通过遗传连锁分析发掘基因及鉴定基因功能需要构建作图群体, 要花费大量的时间和精力,而关联分析可以以自然群体为研究对象,不需构建作图群体就可检测出大量的等位变异。基于连锁不平衡( linkage disequilibrium,LD)的关联分析可以将作物表型性状的多样性与基因(位点)的多态性结合起来, 确定不同种质资源所携带的等位基因及其对目标性状的贡献。赖 勇等[8]运用SSR标记对221份大麦材料进行了遗传多样性分析和结构分析,结果显示,在2016个SSR位点中不管是共线性组合还是非共线性组合都存在一定程度的LD,其中栽培品种的LD水平高于地方品种, 且现代遗传改良的目标性状集中于2H、4H、6H和7H染色体。Kraakman等[9]用AFLP(amplified fragment length polymorphism)和SSR标记技术对148份春大麦的抽穗期、株高及叶锈抗性和大麦黄矮病抗性等进行了LD作图,找到了各性状的相关标记。
本研究拟对从西藏、四川、青海和甘肃等地收集的青稞种质材料的株高、穗长、穗粒数、分蘖数和千粒重等农艺性状进行统计调查,同时运用SSR标记技术对其进行遗传多样性分析,以期为合理利用青稞种质资源、选配亲本及发掘有利基因和标记辅助育种提供帮助。
1 材料与方法
1.1 试验材料
选用来自西藏、四川、青海、甘肃等地的110份六棱青稞品种(系)构成自然群体(表1)。每份材料取10粒饱满种子置于培养皿中,在室内避光培养,10 d后采黄化苗,液氮速冻置-80 ℃备用。2013年和2014年在甘肃农业大学黄羊镇育种试验站亲本圃,每份材料种植4行,行长1.2 m。在成熟收获时对试验所需的材料进行随机抽样,每份材料取10株,测定株高、分蘖数、穗长、穗粒数、千粒重等农艺性状,并取平均值。
1.2 SSR标记分析
采用CTAB法[10]从黄化苗嫩叶中提取各材料基因组DNA,其质量和浓度经紫外分光光度计法检测,于-20 ℃冰箱保存。根据Korff等[11]构建的遗传图谱,选取均匀分布于大麦1H~7H染色体的86对SSR标记(有关株高、穗长、穗粒数、分蘖数、千粒重的标记),进行PCR扩增。扩增体系为10 μL:2×Master Mix 5 μL、10 μmoL·L-1引物1 μL、60 ng·μL-1模板1 μL、ddH2O 3 μL。扩增程序参照赖 勇等[8]。扩增产物经8%聚丙烯酰胺凝胶电泳,银染显色后拍照。
1.3 数据统计分析
各农艺性状的数据在Microsoft Excel 2003中进行统计整理及简单的函数处理。SSR多态性分析和遗传结构分析参照赖 勇等[8]、Maccaferri等[12]和Evanno等[13]的方法。使用TASSEL 2.1软件计算共线、非共线SSR位点组合间的LD水平及支持概率,绘制LD配对检测矩阵图。
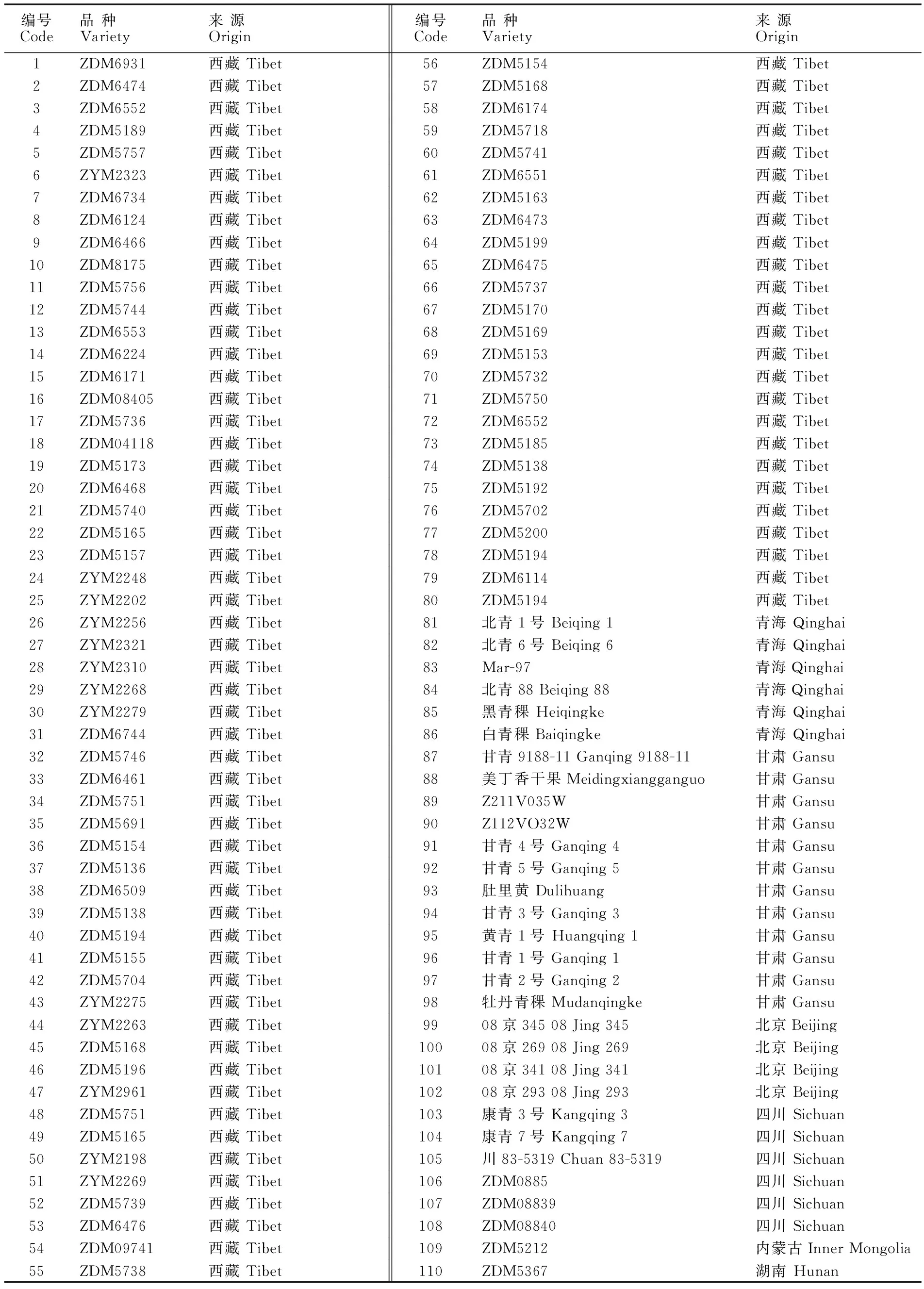
表1 供试大麦材料Table 1 Hulless barleys used in the study
2 结果与分析
2.1 青稞农艺性状的统计分析
对110份青稞种质材料的株高、穗长、分蘖数、穗粒数和千粒重进行调查和统计分析结果(表2)表明,参试青稞种质材料的株高变化范围为53.00~122.00 cm,平均值为96.80 cm,总体较高;穗长变化范围为3.00~12.00 cm,平均值为5.08 cm;分蘖数变化范围为2.00~29.00个,平均值为10.74个;穗粒数变化范围为20.00~90.00粒,平均值为59.35粒;千粒重变化范围为25.46~60.34 g,平均值为40.43 g,其中40 g以下的材料有56份。对110份青稞种质材料的5个农艺性状进行方差分析表明,株高、分蘖数、穗粒数、千粒重这4个农艺性状的差异均达到显著水平,只有穗长没有达到显著水平。供试材料的5个农艺性状的变异系数为2.92%~42.33%,其中,株高的变异系数最小,为2.92%,表明材料间株高变异幅度不是很大,可以选择的范围较其他材料窄,具有较为稳定的遗传机制,且受环境条件的影响相对较小;分蘖数的变异系数达到了42.33%,其次是穗粒数,变异系数为24.35%,较高的变异系数说明这几个性状容易受环境的影响,遗传稳定性较差。
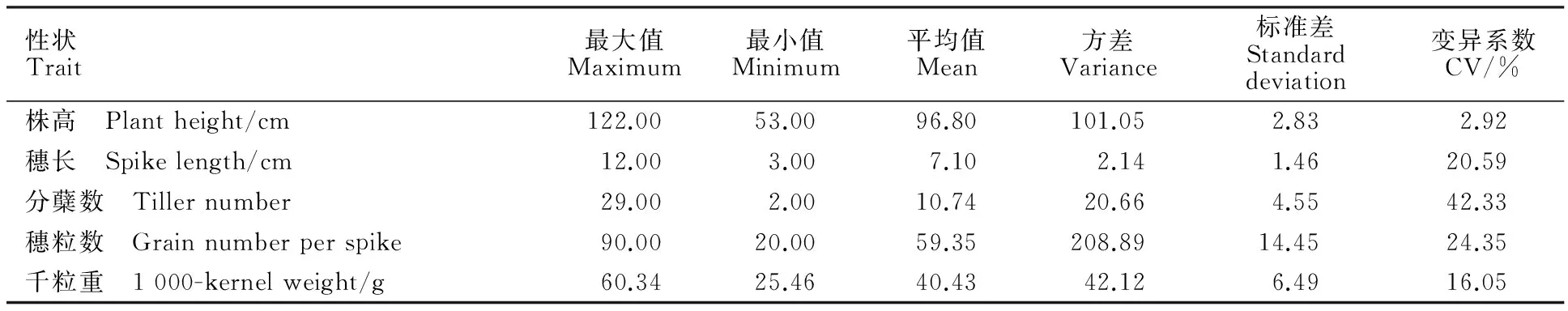
表2 青稞种质材料的农艺性状Table 2 Agronomic traits of hulless barley accessions
2.2 SSR多态性分析
用86对SSR标记对110份青稞种质材料进行了扩增,结果(表3)表明,其中48对SSR标记具有多态性,且这些标记在大麦7条染色体上均有分布。根据48对多态性SSR标记的检测结果可知,110份青稞亲本材料的遗传相似系数变化范围为0.225 0~1.000 0,平均值为0.510 6;在110份青稞种质材料中共检测到168个等位变异,每对标记所检测到的等位变异数在2~6之间,平均为3.2个,其中,Bmag7检测到的等位变异数最多,为6个,HVGLUEND等8个标记各检测到2个等位变异,变异数最少,3H染色体上检测出的等位变异数最多,为28个,1H染色体上检测出的等位变异数最少,为15个;SSR标记的Shannon’s信息指数的变化范围0.672 7~1.136 8,平均为1.050 7,标准差为0.099 4。
2.3 群体遗传结构分析
参照Evanno等[13]的研究方法,选取分布于大麦7条染色体上具有多态性的48对SSR标记对参试青稞的种质材料进行群体遗传结构分析,依据K值所对应的最大似然值选择确定合适的群体数目。结果如图1所示,当K值为3时,△K值出现了明显的峰值,各群体内青稞亲本材料的相似性最大,所以根据最大似然值的原则选定K=3为最终的群体数目。
由如图2所示的群体结构分析图可知,参试的110份青稞种质材料可以分为3个亚群,第1亚群有6份材料,第2亚群有73份材料,第3亚群有31份材料。说明参试的110份材料的群体结构比较简单,能够有效地降低群体结构对关联分析产生的影响。记录每一份材料的对应群体概率的Q值,可以为后期进行关联分析提供相应的协变量数据。
2.4 LD分析
应用软件TASSEL 2.1对48对SSR标记的1 128个共线、非共线的SSR位点组合间的LD水平及支持概率进行计算分析,并根据计算结果绘制了LD配对检测矩阵图(图3)。1 128个共线、非共线的SSR位点成对组合当中,均出现了不同水平的LD,且LD水平整体较高。当D′取值为0.2~0.4和0.4~0.6时,LD的成对位点数最多,分别为203和102;D′为0.8~1.0时最少,仅有13个(表4)。D′统计概率(P<0.01)支持的LD成对位点数为438个,占总位点数的38.8%,D′的平均值为0.36。由图3可知,在7个染色体群中均出现较高水平的LD成对位点(D′>0.5),但主要分布在2H、4H、6H和7H连锁群上,且在染色体2H和6H连锁群上较高水平的LD位点(D′>0.5)比较集中,分别出现了7个和11个。对共线性位点组合的LD状况进一步分析,发现在染色体2H、3H和6H连锁群LD成对位点明显多于其他几条染色体连锁群。

表3 48对SSR标记在110份青稞种质材料中的多态性Table 3 Polymorphism of fourty-eight SSRs among 110 hulless barley accessions

K:群体数目。
K:Number of population.
图1 ΔK值随K值的变化图
Fig.1ΔKwith change ofKvalues

图2 110份青稞亲本材料的群体遗传结构图
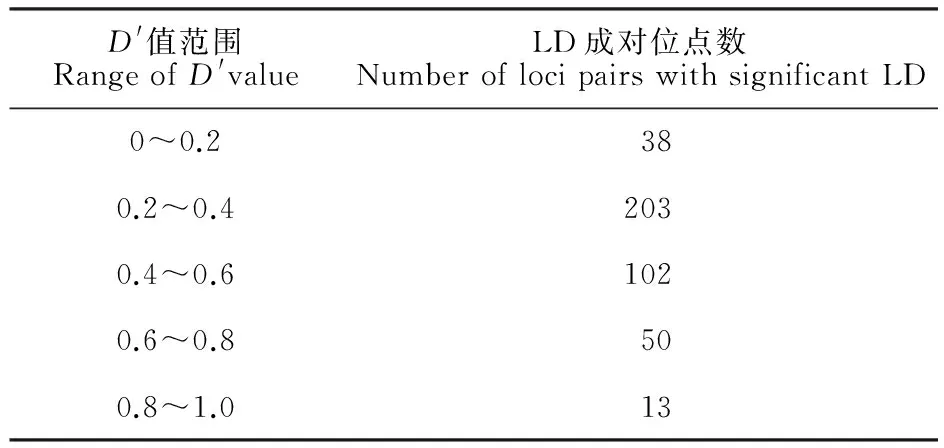
D'值范围RangeofD'valueLD成对位点数NumberoflocipairswithsignificantLD0~0.2380.2~0.42030.4~0.61020.6~0.8500.8~1.013
3 讨 论
3.1 根据青稞农艺性状评价种质资源的效果
农艺性状是评价种质资源最经济简捷的方法,由于其易于观测并在一定程度上也能反映出内在基因型的变异情况,因此已被广泛应用于作物种质材料遗传多样性研究和品种选育[14-15]。本研究对110份青稞品种(系)的株高、穗长、穗粒数、千粒重和分蘖数等性状进行了统计分析,结果显示,参试材料各性状变异较大,变化范围在2.92%~42.33%之间,这与前人的研究结果相一致[16-17]。就株高而言,多数参试材料在95cm 以上,变异系数小,说明在青藏高原栽培的青稞种质材料主要以高杆为主。千粒重是作物产量构成的三要素之一,本研究中千粒重的平均值为40.43 g,表明参试材料的千粒重总体上较高,因此可从中选取千粒重大的种质资源杂交以获得更高千粒重的品种(系)来提高青稞的产量。穗粒数和分蘖数等的变化范围较大,表现出极为丰富的多样性,可用于构建核心种质库。通过对农艺性状的综合评价,对优良性状多的材料直接用于育种亲本,提供一定的参考。然而,尽管农艺性状对作物种质的评价是有效的[19],但品质和抗性也至关重要,在后续的评价中应综合考虑。
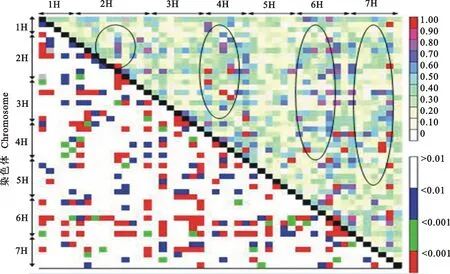
双向箭头指示连锁群区域;对角线上方为成对位点间的D′值;对角线下方为成对位点间LD的支持概率;椭圆圈显示较高水平的LD位点分布。
Double-headed arrows show the region of linkage groups;D′ value of the corresponding marker pair and itsP-value are shown in the upper and lower triangles,respectively; The oval circles show distribution of LD loci in high levels.
图3 大麦7个连锁群SSR位点的连锁不平衡分布
Fig.3 Distribution of LD among SSR loci on 7 linkage groups of hulless barley
3.2 青稞种质资源的群体遗传结构以及连锁不平衡分析评价
为了弥补农艺性状评价种质资源的局限性,利用分子标记技术分析不同材料间的遗传差异,明确它们的遗传关系,为后期青稞亲本杂交组合配置提供一定的参考[19-21]。本研究以SSR标记技术对来自西藏、青海、甘肃和四川等地的110份青稞品种(系)进行了遗传结构及连锁不平衡分析。结果显示,110份青稞种质材料被分为3个亚群,说明这些青稞材料的整体遗传多样性不够丰富。根据48对多态SSR标记的检测结果,110份青稞亲本材料的遗传相似性偏高,说明参试材料亲缘关系较近,主要原因可能是由于参试材料主要来源于青藏高原,这与赖 勇等[22]、冯宗云等[23]、杨 平等[24]的研究结果较一致。本研究中1 128个SSR位点的共线性组合和非共线性组合中,D′统计概率(P<0.01)支持的LD成对位点数为438个,占总位点数的38.8%,且主要集中在D′值的0.2~0.4范围内(203个),总体来看LD水平较高。在7个连锁群均有较高水平的LD(P>0.5)位点,但主要集中在2H、4H、6H和7H染色体上,这对关联分析和有利基因的发掘均有非常重要的意义。赖 勇[8]等选用大麦的64对SSR标记对大麦种质材料进行了分析,结果显示,共线性或非共线性位点组合均有一定程度的LD,且整体LD水平较高。Kraakman等[9]研究表明,栽培大麦的LD水平最高,地方品种次之,野生资源LD 水平最低。因此在后期进行关联分析挖掘青稞种质资源的有利基因过程中,需增加其他不同区域来源的材料,丰富遗传基础,提高关联结果的准确性。为了更有效地改善现有青稞遗传基础狭窄的问题,需要加强对其野生近缘种的开发利用,为培育优良青稞新品种提供宝贵的基因资源。
[1] 张赤红,张 京,赵会英,等.应用SSR标记对61个国家大麦遗传多样性的研究[J].植物遗传资源学报,2008,9(1):15.
ZHANG C H,ZHANG J,ZHAO H Y,etal.Study on genetic diversity of barley from 61 countries using SSR marker [J].JournalofPlantGeneticResource,2008,9(1):15.
[2]DOTLACIL L,HERMUTH J,STEHNO Z,etal.Diversity in European winter wheat land races and obeolete obsolete cultivars [J].CzechJouralofGenetics&PlantBreeding,2000,36(2):29.
[3]DELACY I H,SKOVMAN B,HUERTA J.Characterization of Mexican wheat land races using agronomically useful attributes [J].GeneticsResourcesandCropEvolution,2000,47(6):591.
[4]TANKSLEY S D,MCCOUCH S R.Seed banks and molecular maps: unlocking genetic potential from the wild [J].Science,1997,277(5329):1063.
[5]BHAGWAT A A,CREGAN P B,AKKAYA M S.Length polymorphisms of simple sequence repeat DNA in soybean [J].Genetics,1992,132:1131.
[6]MAROOF M S,BIYASHEV R M,YANG G P,etal.Extraordinarily polymorphic microsatellite DNA in barley: species diversity,chromosomal locations,and population dynamics [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1994,91:5466.
[7]WANG Z,WEBER J L,ZHONG G,etal.Survey of plant short tandem DNA repeats [J].TheoreticalandAppliedGenetics,1994,88:1.
[8] 赖 勇,孟亚雄,王化俊,等.大麦遗传多样性及连锁不平衡分析[J].作物学报,2013,39(12):2154.
LAI Y,MENG Y X,WANG H J,etal.Genetic diversity and linkage disequilibrium analysis in barley [J].ActaAgronomicaSinica,2013,39(12):2154.
[9]KRAAKMAN A W,MARTNEZ F,MUSSIRALIEV B,etal.Linkage disequilibrium mapping of morphological,resistance,and other agronomically relevant traits in modern spring barley cultivars [J].MolecularBreeding,2006,17:41.
[10]POREBSKI S,BAILEY L G,BAUM B R.Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components [J].PlantMolecularBiologyReporter,1997,15:8.
[11]EARL D A,VONHOLDT B M.STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method [J].ConservationGeneticsResources,2012,4:359.
[12]MACCAFERRI M,SANGUINETI M C,NOLI E,etal.Population structure and long-range linkage disequilibrium in a durum wheat elite collection [J].MolecularBreeding,2005,15:271.
[13]EVANNO G,REGNAUT S,GOUDET J.Detecting the number of clusters of individuals using the software STRUCTURE: as imulation study [J].EcologicalEntomology,2005,14:2611.
[14] 孟 霞,卓 嘎,栾运芳.西藏部分青稞主要农艺性状分析[J].麦类作物学报,2010,30(6):1043.
MENG X,ZHUO G,LUAN Y F.Analysis of main agronomic traits of several Tibetan naked barley [J].JournalofTriticeaeCrops,2010,30(6):1043.
[15] 陈晓静,沈会权,乔海龙,等.大麦种质资源形态特征及农艺性状的分析[J].江苏农业学报,2007,23(6):532.
CHEN X J,SHEN H Q,QIAO H L,etal.Analysis of agronomic trait and morphological characteristics of barley germplasm resources [J].JiangsuJournalofAgriculturalScience,2007,23(6):532.
[16] 李守明,魏凌基,梁 维,等.新疆大麦种质资源农艺性状和醇溶蛋白遗传多样性分析[J].新疆农业科学,2009,46(2):269.
LI S M,WEI L J,LIANG W,etal.Agronomic trait of germplasm resources and genetic diversity of hordein in Xinjiang barley [J].XinjiangAgriculturalSciences,2009,46(2):269.
[17] 陈丽华,张志斌,侯志强.青海省青稞主栽品种农艺性状分析[J].江西农业大学学报,2012,34(3):439.
CHEN L H,ZHANG Z B,HOU Z Q,etal.Analysis of agronomic characters of major hulless barley cultivars in Qinghai province [J].ActaAgriculturaeUniversitatisJiangxiensis,2012,34(3):439.
[18] 莫惠栋,顾世梁.江浙沪大麦品种农艺性状的聚类分析[J].中国农业科学,1987,20(3):28.
MO H D,GU S L.Jiangzhehu barley varieties of agronomic traits of clustering analysis [J].ScientiaAgriculturaSinica,1987,20(3):28.
[19]HARRIS B P,STOKESBURY K D E.The spatial structure of local surficial sediment characteristics on Georges Bank,USA [J].ContinentalShelfResearch,2010,30(17):1840.
[20]WANG M L,ZHU C S,BARKLEY N A,etal.Genetic diversity and population structure analysis of accessions in the US historic sweet sorghum collection [J].TheoreticalandAppliedGenetics,2009,120:13.
[21]COCKRAM J,WHITE J,LEIGH F J,etal.Association mapping of partitioning loci in barley [J].BMCGenetics,2008,9:16.
[22] 赖 勇,王晋民,任 龙,等.大麦SSR标记遗传多样性及连锁不平衡分析[J].核农学报,2016,30(10):1889.
LAI Y,WANG J M,REN L,etal.Genetic diversity and linkage disequilibrium analysis of barley using SSR markers [J].JournalofNuclearAgriculturalSciences,2016,30(10):1889.
[23] 冯宗云,张义正,凌宏清.应用微卫星标记研究西藏野生二棱大麦的遗传多样性及地理分化[J].高技术通讯,2003,13(10):46.
FENG Z Y,ZHANF Y Z,LING H Q.Genetic diversity and geographical differentiation ofHordeumvulgaressp.spontaneumin Tibet using microsatellite markers [J].ChineseHighTechnologyLetters,2003,13(10):46.
[24] 杨 平,刘仙俊,刘 新,等.利用SRAP标记研究四川高原青稞育成品种的遗传多样性[J].遗传,2008,30(1):115.
YANG P,LIU X J,LIU X,etal.The application of SRAP-PCR in genetic diversity study of Sichuan hulless barley cultivar [J].Genetic,2008,30(1):115.
Agronomic Characteristics and Genetic Analysis in Hulless Barley
MENG Yaxiong1,2,MA Zengke1,2,REN Panrong1,2,SI Erjing1,2,WANG Juncheng1,2,YANG Ke1,2,ZHANG Haijuan1,2,MA Xiaole1,2,WANG Yucai1,2,WANG Huajun1,2
(1.Gansu Provincial Key Laboratory of Aridland Crop Science/Gansu Key Laboratory of Crop Improvement & Germplasm Enhancement,Lanzhou,Gansu 730070,China; 2.College of Agronomy,Gansu Agricultural University,Lanzhou,Gansu 730070,China)
In order to provide a suitable evaluation for the agronomic traits and genetic characteristics of 110 hulless barley germplasm materials,the plant height,spike length,tiller number,grain number per spike and 1 000-kernel weight traits were analyzed statistically and fourty-eight pairs of SSR markers distributed on chromosomes 1H to 7H were used for genetic analysis.The results showed that there were large differences of agronomic characters: the variable coefficients ranged from 2.92% to 42.33%,and the variation range of plant height,ear length,tiller number,grain number per spike and 1 000-kernel weight trait was 53-122 cm,2.5-7,2-19,20-90,25.46-60.34 g,respectively.A total of 168 alleles were detected in 110 hulless barley accessions with 2 to 6 alleles per locus. All accessions were clustered into three major groups. There were linkage disequilibrium (LD) among linked loci and unlinked loci pairs,and 438 out of 1 128 loci (38.8%) had significant LD (P<0.01) with averageD′-value of 0.36. The LD (D′<0.5) level of hulless barley was high,and the loci were mainly distributed on chromosomes 2H,4H,6H and 7H.
Hulless barley; Agronomic characteristics; SSR marker; Linkage disequilibrium
时间:2017-01-03
2016-03-24
2016-12-10
国家自然科学基金地区项目(31460347);盛彤笙科技创新基金项目(GSAU-STS-1513);国家大麦青稞产业技术体系项目(CARS-05)
E-mail:yxmeng1@163.com
S512.3;S330
A
1009-1041(2017)01-0048-08
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20170103.1625.014.html
猜你喜欢
作文小学高年级(2022年5期)2022-06-16
星星·诗歌原创(2022年1期)2022-04-16
环境卫生工程(2021年4期)2021-10-13
疯狂英语·新读写(2021年2期)2021-02-25
落叶果树(2021年6期)2021-02-12
西藏人文地理(2020年4期)2020-11-19
甘肃教育(2020年18期)2020-10-28
天津农林科技(2020年3期)2020-08-13
翠苑(2019年4期)2019-11-11
华人时刊(2016年19期)2016-04-05
