
采后O3处理对采前CPPU处理猕猴桃果实乙烯代谢的影响
2018-08-17 02:12苏苗罗安伟李琳李圆圆白俊青李锐方沂蒙宋俊奇蔺志颖刘占德
中国农业科学 2018年15期
苏苗,罗安伟,李琳,李圆圆,白俊青,李锐,方沂蒙,宋俊奇,蔺志颖,刘占德
采后O3处理对采前CPPU处理猕猴桃果实乙烯代谢的影响
苏苗1,罗安伟1,李琳1,李圆圆1,白俊青1,李锐1,方沂蒙1,宋俊奇1,蔺志颖1,刘占德2
(1西北农林科技大学食品科学与工程学院,陕西杨凌 712100;2西北农林科技大学园艺学院,陕西杨凌 712100)
【目的】探究臭氧(ozone,O3)是否能有效减轻膨大剂(N-2-氯-4-吡啶基苯-N’-苯基脲,CPPU)对猕猴桃带来的负面影响,为市场上使用CPPU猕猴桃的贮藏提供信息。【方法】以采前盛花期28d使用20 mg·L-1CPPU处理和对照用清水蘸果处理的秦美猕猴桃为试验材料,研究不同浓度的O3处理(0、10、40、70 mg·m-3)对贮藏期间猕猴桃果实乙烯代谢过程中的蛋氨酸(methionine,Met)、S-腺苷蛋氨酸(S-adenosyl methionine,SAM)、1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)含量及其相关代谢酶1-氨基环丙烷-1-羧酸合成酶(ACC synthase,ACS)和1-氨基环丙烷-1-羧酸氧化酶(ACC oxidase,ACO)活性的影响。【结果】CK组(未使用CPPU也未使用O3处理的为CPPU对照组CK)在贮藏过程中Met、SAM含量以及SAM合成酶、ACS、ACO活性的下降速率均低于CK1组(使用CPPU但未使用O3处理的为臭氧处理对照组CK1);贮藏60 d时,CK1和各O3处理(10、40、70 mg·m-3)的Met含量分别为1.36、2.62、4.41和2.60 mg·(100 g)-1,O3处理显著高于CK1(<0.05);CK1和40 mg·m-3O3处理的SAM含量分别为15.48 mg·(100 g)-1和20.73 mg·(100 g)-1,具有显著性差异(<0.05),而CK1和10 mg·m-3O3处理组、70 mg·m-3O3处理组无显著性差异(>0.05);CK1和各O3处理(10、40、70 mg·m-3)的ACC含量分别为0.068、0.059、0.038和0.055 nmol·g-1,40、70 mg·m-3O3处理与CK1具有显著性差异(<0.05);CK1和各O3处理(10、40、70 mg·m-3)的ACS活性分别为0.084、0.069、0.054和0.080 nmol·(g·h)-1;ACO活性的峰值分别为0.062、0.046、0.029和0.051 nmol·(g·h)-1,O3处理和CK1之间存在显著性差异(<0.05);CK1和各O3处理(10、40、70 mg·m-3)的乙烯的峰值分别为18.42、15.99、9.86、11.69 μL·kg-1·h-1;呼吸高峰分别是18.77、16.15、12.24、15.48 mg·kg-1·h-1。【结论】CPPU增加了猕猴桃乙烯释放量,加速了果实软化,对猕猴桃贮藏带来负面影响,O3处理能有效抑制猕猴桃的乙烯代谢,延缓果实软化,因此O3处理能有效减缓因使用CPPU而导致的猕猴桃后熟软化进程。
臭氧;膨大剂;猕猴桃;乙烯代谢;呼吸速率
0 引言
【研究意义】猕猴桃果实耐贮性及其贮藏品质都与采后生理变化密切相关,特别是采后果实的乙烯代谢和呼吸作用。这是由于乙烯促进了果实的成熟、衰老,而呼吸作用则为果实采后正常生理代谢提供了能量及中间产物[1]。乙烯是植物体内的重要激素,参与植物生长发育和衰老的整个过程,包括果实的后熟软化[2]。在植物体内乙烯的合成途径为蛋氨酸(Met)→S-腺苷蛋氨酸(SAM)→1-氨基环丙烷-1-羧酸(ACC)→乙烯,SAM合成酶催化蛋氨酸合成SAM,ACC合成酶(ACS)催化SAM生成ACC,ACC在ACC氧化酶(ACO)作用下最终生成乙烯[3]。猕猴桃属于典型的呼吸跃变型果实,对乙烯极为敏感,采后极易软化;如何抑制乙烯合成、降低果实呼吸速率是猕猴桃贮藏保鲜的核心所在。膨大剂俗称“膨大素”、“大果灵”,化学名称为细胞激动素。常用膨大剂的有效成分为氯吡脲(CPPU),可以促进细胞分裂、分化、扩大和细胞形成[4],从而具有促进果实膨大的作用。膨大剂被广泛应用于柿子、甜瓜、苦瓜、猕猴桃、葡萄、番茄、苹果、梨、西瓜等果蔬中[5]。猕猴桃生产中为了增加单果重和亩产量,在幼果膨大期普遍使用较高浓度CPPU处理,但这会显著增加采后果实的呼吸速率和乙烯释放量,加快果实后熟和软化,缩短贮藏时间,为猕猴桃采后贮藏保鲜带来了极大地负面效应。O3在杀菌、消毒过程中只产生无毒无害的氧化物,并最终被还原为氧气,不存在二次污染,2001年被美国FDA列为可直接和食品接触的添加剂。O3因具有杀菌、抑制和消除乙烯、减缓果实的呼吸作用而在果蔬贮藏保鲜中逐渐得到应用[6];O3还能调节果蔬新陈代谢,延长果实的贮藏期,减少果实软化速度,抑制菌丝的生长等[7-10]。【前人研究进展】何靖柳等[11]对红阳猕猴桃进行不同保鲜处理,研究其贮藏期生理及品质变化的影响,结果表明,O3处理后贮藏至120 d的猕猴桃,其呼吸强度、硬度、腐烂率、可溶性固形物等指标均优于其他处理。李子龙等[12]对猕猴桃进行臭氧处理,探究臭氧对其酶活性的影响,试验结果表明O3浓度为6 g·L-1可以抑制猕猴桃果汁PPO酶的活性,可以防止褐变。HAN等[13]对桑果进行O3处理,发现臭氧能有效降低其呼吸速率和乙烯释放量和多酚氧化酶。【本研究切入点】虽然已经有较多的猕猴桃贮藏的研究报道,但是并未见O3处理对猕猴桃乙烯代谢途径的相关研究。【拟解决的关键问题】本文以生长期使用CPPU的猕猴桃为试材,探究O3处理能否有效减轻因使用20 mg·L-1CPPU对猕猴桃果实乙烯代谢带来的负面影响,为O3保鲜技术及其在猕猴桃贮藏保鲜上的应用提供科学依据。
1 材料与方法
1.1 材料与试剂
猕猴桃秦美品种采自陕西省杨凌示范区(管理良好的果园)。在猕猴桃盛花期后28 d,用20 mg·L-1CPPU进行蘸果处理,对照用清水蘸果处理(CPPU对照组CK);当果实生长至可溶性固形物含量达到6.0%—6.5%时采收。剔除有机械损伤、病斑及畸形果实,预冷24 h后入库。
氯吡脲(CPPU),四川省兰月科技有限公司;HgCl2,山东西亚化学股份有限公司;NaClO,广州华大化学试剂有限公司;PVP,上海蓝季科技发展有限公司;NaHCO3,广东光华科技股份有限公司;FeSO4,西陇化工股份有限公司;ACC、交联聚乙烯吡咯烷酮(PVPP)、二硫苏糖醇(DTT)、SAM、Tris、抗坏血酸钠、蛋氨酸均为北京索莱宝科技有限公司。
1.2 仪器与设备
GC-14C气相色谱,日本岛津;LC-20A高效液相色谱,日本岛津;HC-3018R高速冷冻离心机,安徽中科中佳科学仪器有限公司;有毒有害气体检测报警仪,深圳市瑞凯雷科技有限公司;O3发生器,青岛欣美净化设备有限公司。
1.3 方法
1.3.1 处理分组 将预冷后的猕猴桃装于网眼塑料筐中,分别放置于4个小型试验冷库中(每个冷库容积23 m3),每库20箱,每箱15 kg,库内温度(0±1)℃,RH 90%—95%。对使用20 mg·L-1CPPU处理的猕猴桃进行O3处理,将额定产量为20 g·h-1的O3发生器产生的O3通入冷库中,使3个冷库浓度分别达到10、40、70 mg·m-3(分别为O3处理1、处理2、处理3,记为T1、T2、T3;用O3检测仪对O3浓度进行检测);未使用CPPU也未使用O3的为CPPU对照组CK,使用CPPU但未使用O3处理的为臭氧处理对照组CK1;每15 d 处理1次,每次处理2 h;随机取样,乙烯含量和呼吸强度每15 d测定1次,Met、SAM、ACC、MACC含量以及SAM合成酶(SAMS)、ACS、ACO活性30 d测定1次,试验设3次重复。
1.3.2 测定指标
(1)蛋氨酸含量的测定参照Kühnreich等[14]的方法略有改动。
称取6.0 g猕猴桃研磨,于12 000×离心15 min,将上清液定容到10 mL容量瓶中,经0.22 μm滤膜过滤后进行HPLC检测。
高效液相色谱条件:采用C18柱,流动相为V(12.5 mmol·L-1磷酸)﹕V(乙腈)=97﹕3,柱温30℃,流速1 mL·min-1,检测波长210 nm,进样量10 μL。
(2)SAM含量的测定参照杨宇辉[15]略有改动。
称取2.0 g猕猴桃研磨,于10 000×离心15 min,
将上清液倒掉,在残渣中加入2.0 mL 1.5 mol·L-1的HClO4,常温下震荡90 min,于4℃过夜,12 000×离心10 min,经0.22 μm滤膜过滤后进行HPLC检测。
高效液相色谱条件:采用C18柱,流动相为V(0.01 mmol·L-1甲酸铵)﹕V(甲醇)=97﹕3,柱温30℃,流速1 mL·min-1,检测波长254 nm,进样量10 μL。
(3)ACC含量的测定参考曹建康等[16]的方法。
称取5.0 g猕猴桃研磨,转入试管中并加入10 mL 95%乙醇溶液,煮沸浸提20 min,冷却后于10 000×离心15 min,收集上清液,再向残渣中加入10 mL 80%乙醇,于70℃浸提30 min,离心收集上清液,合并两次上清液,用旋转蒸发仪浸提蒸发至干,向蒸发残留物中加入2.0 mL蒸馏水振荡溶解,即为ACC制备液。
取1.0 mL ACC制备液,置于容积为20 mL的样品瓶中,加入40 μL 25mmol·L-1HgCl2溶液,用橡胶塞密封样品瓶,冰浴10 min使温度平衡。再通过注射器加入0.2 mL经冰浴预冷的5% NaClO-饱和NaOH混合液(NaClO﹕NaOH=2﹕1,体积比)迅速振荡5 s后,放回冰浴平衡5 min,顶空取1 mL气体,用气相色谱法测定乙烯生成量。
(4)1-(丙二酰氨基)环丙烷-1-羧酸(MACC)含量的测定。
取1.0 mL ACC制备液,加入0.2 mL 12 mol·L-1盐酸,于沸水浴中加热3 h。冷却后加入1.2 mL 2 mol·L-1NaOH溶液中和,然后按照测定ACC的方法测定,即得总的ACC含量,MACC含量为总的ACC含量减去游离ACC含量。
(5)SAM合成酶活性的测定参照姚高峰等[17]方法略有改动。
称取10 g猕猴桃研磨,于12 000×离心15 min,将上清液倒掉,在残渣中加入10 mL Tris-HCl缓冲液,按冰浴30 s震荡1 min交替进行8次,于12 000×离心10 min,即为酶液。在室温下反应1 h,加入1.5 mL 20% HClO4,4℃放置0.5 h以上终止反应,12 000×离心10 min,经0.22 μm滤膜过滤后进行HPLC检测。
高效液相色谱条件:采用C18柱,流动相为V (0.01 mmol·L-1甲酸铵)﹕V(甲醇)=97﹕3,柱温30℃,流速l mL·min-1,检测波长254 nm,进样量20 μL。
(6)ACS活性的测定。
取5 g猕猴桃加入5.0 mL含有1 mmol·L-1EDTA、1 mmol·L-1PMSF、4 mmol·L-1DTT、3% PVPP和10 μmol·L-1磷酸吡哆醛的提取缓冲液进行研磨,于12 000×离心30 min,上清液即为酶提取液。
将1 mL酶提取液和1.5 mL反应缓冲液加入到20 mL样品瓶中,用橡皮塞密封样品瓶后,在30℃水浴中保温1 h。然后注射加入0.1 mL 25 mmol·L-1HgCl2溶液以终止反应,并置于冰浴10 min平衡温度,再用注射器加入0.2 mL经冰浴预冷的5% NaClO-饱和NaOH混合液(NaClO﹕NaOH=2﹕1,体积比)迅速震荡5 s后,放回冰浴平衡5 min,顶空取1 mL气体测定乙烯释放量。
(7)ACO活性的测定。
称取5 g猕猴桃,加入5.0 mL含有10%甘油、5% PVP、5.0 mmol·L-1DTT、30 mmol·L-1抗坏血酸钠和0.1 mmol·L-1FeSO4提取缓冲液,于12 000×离心30 min,上清液即为酶提取液。
将2.0 mL酶提取液和2 mL反应缓冲液(加入30 mmol·L-1NaHCO3)加入到20 mL样品瓶中,用橡皮塞密封样品瓶后,在30℃摇床震荡2 h。然后顶空取1 mL气体测定乙烯释放量。
(8)乙烯释放量的测定。
取1 kg果实放置在1.6 L的密封干燥器内1 h,抽顶空气体,通过气相色谱(GC-14C型气相色谱仪)进行测定。气相条件:FID检测器,柱温90℃,进样口温度160℃,GDX-102不锈钢填充柱,载气:N2(50 MPa),燃气:H2(75 MPa),助燃气:空气(50 MPa);对照及处理果实各设3组重复。
(9)硬度测定采用TAXT PLUS/50物性测定仪。
在TPA模式下,随机取4个果实进行测定,在猕猴桃果实赤道部位均匀取3点,削去果实表皮,沿果实赤道上120°等距离测定3次,重复3次。
1.4 数据处理方法
采用Origin8.0软件进行作图分析,SPSS 20软件对数据进行显著差异性检验,<0.05被认为具有显著性差异。
2 结果
2.1 O3处理对猕猴桃果实乙烯代谢途径中主要物质含量变化的影响
2.1.1 O3处理对猕猴桃果实Met含量的影响 Met是植物体内合成乙烯的前体物质。从图1-a中可以看出,在贮藏前期Met含量较高,随着贮藏时间的延长,Met含量呈下降趋势。在贮藏开始时CK的Met含量低于CK1,在贮藏过程中,CK的Met下降趋势小于CK1。在贮藏60 d后,Met含量下降减缓,60 d时CK1和各O3处理组(10、40、70 mg·m-3)的Met含量分别为1.36、2.62、4.41和2.60 mg·(100 g)-1,40 mg·m-3O3处理的Met含量是CK1的3.24倍;且在整个贮藏期,40 mg·m-3O3处理组Met均高于其他处理和CK1。
2.1.2 O3处理对猕猴桃果实SAM含量的影响 SAM是由Met转化生成的,随着Met含量的减少,SAM含量也随之减少(图1-b)。CK下降趋势低于CK1。贮藏60 d时CK1和40 mg·m-3O3处理组的SAM含量分别为15.48 mg·(100 g)-1和20.73 mg·(100 g)-1,具有显著性差异(<0.05),且在整个贮藏期内40 mg·m-3O3处理SAM含量均高于其他处理和CK,和对Met的影响一致,10、70 mg·m-3O3处理效果较差,差异不显著(>0.05)。
2.1.3 O3处理对猕猴桃果实ACC和MACC含量的影响 ACC是乙烯合成的直接前体物质,经过ACC氧化酶生成乙烯,这是乙烯合成途径中的限速步骤。图2-a中,随着贮藏时间的延长,ACC呈先上升后下降的趋势,在60 d时出现高峰。ACC含量增加是由SAM转化而来,在60 d之后SAM保持不变,而ACC向乙烯转化,故呈下降趋势。CK较CK1的ACC含量低,CPPU加快了ACC含量的增加,从而促进内源乙烯的释放;CK1与各O3处理组相比,O3能有效减少果实中ACC含量的增加,从而抑制乙烯释放量的增加。60 d时CK1和各O3处理(10、40、70 mg·m-3)的ACC含量分别为0.068、0.059、0.038和0.055 nmol·g-1,40、70 mg·m-3O3处理与CK1具有显著性差异(<0.05)。
在ACC代谢途径中,除向乙烯转化外,ACC还可以转化生成MACC(图2-b),在贮藏过程中MACC含量呈上升趋势。
不同浓度O3处理效果不同,10 mg·m-3O3处理由于浓度太低,没有达到预期效果;40 mg·m-3O3处理效果较好;70 mg·m-3O3处理因浓度过高,O3的强氧化活性可能损伤了细胞结构,造成ACC和MACC含量均高于40 mg·m-3O3处理。
2.2 O3处理对猕猴桃果实乙烯相关酶活性的影响
SAM合成酶是催化Met生成SAM的关键酶,ACS、ACO是乙烯合成途径中的关键酶和限速酶。抑制SAM合成酶、ACS和ACO酶的活性,均可以抑制乙烯的生成,延缓果实后熟软化。

图1 O3 对使用20 mg·L-1 CPPU秦美猕猴桃Met(a)、SAM(b)含量的影响

图2 O3 对使用20 mg·L-1 CPPU秦美猕猴桃ACC(a)、MACC(b)含量的影响
2.2.1 O3处理对猕猴桃果实SAM合成酶活性的影响 SAM合成酶催化Met生成SAM。图3显示,SAM合成酶在贮藏过程中呈先上升后下降的趋势,在贮藏30 d和120 d时分别达到高峰。在整个贮藏期,40 mg·m-3O3处理均低于其他处理和CK1,40 mg·m-3O3处理与CK1有显著性差异(<0.05),10、70 mg·m-3O3处理之间差异不显著(>0.05)。
2.2.2 O3处理对猕猴桃果实ACS活性的影响 ACS是催化SAM生成ACC的关键酶。图4-a中,贮藏前期ACS活性呈上升趋势,在60 d时达到高峰,之后下降,这与ACC含量的变化趋势一致(图2-a)。在贮藏过程中CK的ACS活性低于CK1。60 d时CK1和各O3处理组(10、40、70 mg·m-3)的ACS活性分别为0.084、0.069、0.054和0.080 nmol·(g·h)-1;且在整个贮藏期,40 mg·m-3O3处理ACS活性均低于其他处理和CK1,40 mg·m-3O3处理与CK1具有显著性差异(<0.05),10、70 mg·m-3O3处理之间差异不显著(>0.05)。
2.2.3 O3处理对猕猴桃果实ACO活性的影响 ACO以抗坏血酸和氧作为辅基,Fe2+和CO2作为辅助因子,将ACC氧化成乙烯。图4-b显示,ACO活性随着贮藏时间的延长呈先上升后下降趋势,在60 d时出现高峰。同样,在贮藏过程中CK的ACO活性低于CK1,CK1和各O3处理组(10、40、70 mg·m-3)ACO活性的峰值分别为0.062、0.046、0.029和0.051 nmol·(g·h)-1,处理组和CK1之间存在显著性差异(<0.05),40 mg·m-3O3处理对ACO活性的抑制效果较适宜,10、70 mg·m-3O3处理之间差异不显著(>0.05)。
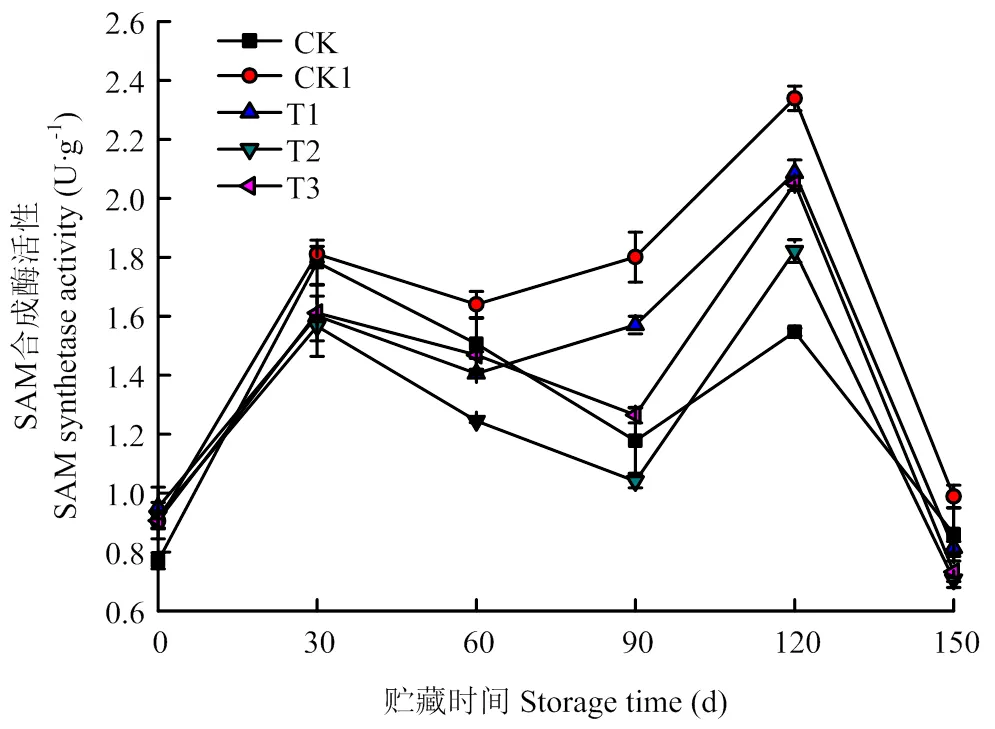
图3 O3对使用20 mg·L-1 CPPU秦美猕猴桃SAM合成酶活性的影响
2.3 O3处理对猕猴桃果实乙烯释放量、呼吸速率的影响
猕猴桃是典型的呼吸跃变型水果,乙烯能催化果实软化,加速果实衰老,乙烯释放量越高,果实软化越快。O3处理对果实乙烯释放量的影响如图5-a所示,CK乙烯释放量低于CK1,CPPU增加了猕猴桃乙烯的释放,从而影响贮藏时间。CK1和O3处理组(10、40、70 mg·m-3)果实的乙烯释放量均在45 d时达到高峰,其值分别为18.42、15.99、9.86、11.69 μL·kg-1·h-1,且乙烯高峰出现时间晚于呼吸高峰,乙烯释放量的变化同ACC含量变化一致(图2-a)。40 mg·m-3O3处理组果实的乙烯释放量在整个贮藏期始终最低,是较适宜的处理浓度。
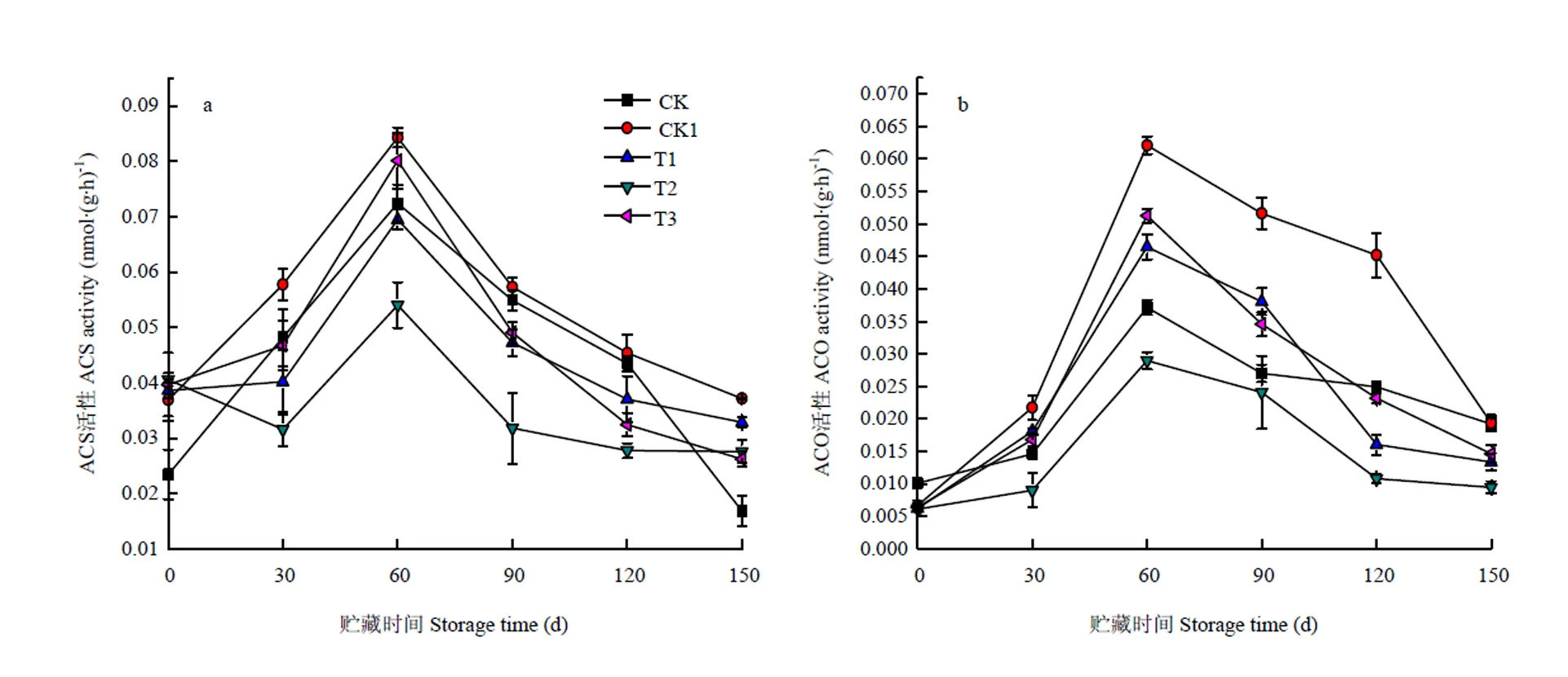
图4 O3对使用20 mg·L-1 CPPU秦美猕猴桃ACS(a)、ACO(b)的影响
如图5-b所示,O3处理和CK1的呼吸速率呈先上升后下降趋势,O3处理呼吸高峰出现在贮藏时间30 d,而CK1则在15 d出现呼吸高峰(CK的呼吸峰值要低于CK1),且呼吸峰值要高于O3处理组,CK1和O3处理组(10、40、70 mg·m-3)的呼吸峰值分别为18.77、16.15、12.24、15.48 mg·kg-1·h-1,O3不仅能降低呼吸峰值还能延迟呼吸峰出现的时间。随着贮藏时间的延长,O3处理果实呼吸强度始终低于CK1(<0.05)。40 mg·m-3O3处理效果适宜,贮藏期内果实呼吸强度最低;10 mg·m-3和70 mg·m-3O3处理对果实呼吸作用的抑制效果均较差。
2.4 O3处理对猕猴桃果实硬度的影响
果实硬度是影响贮藏效果的关键因素,随着贮藏时间的延长,硬度呈逐渐下降趋势(图6)。CK贮藏前期高于CK1,随着贮藏时间的延长,40 mg·m-3O3处理的硬度高于CK;O3处理果实的硬度高于CK1,贮藏时间为30 d时,各处理硬度分别下降了15.95%、7.61%、6.00%和10.17%。贮藏30—75 d时硬度呈现快速下降趋势,之后的贮藏期内硬度下降较为缓慢。在贮藏中期,40 mg·m-3O3处理的硬度显著高于CK1(<0.05),10、70 mg·m-3O3处理效果较差。
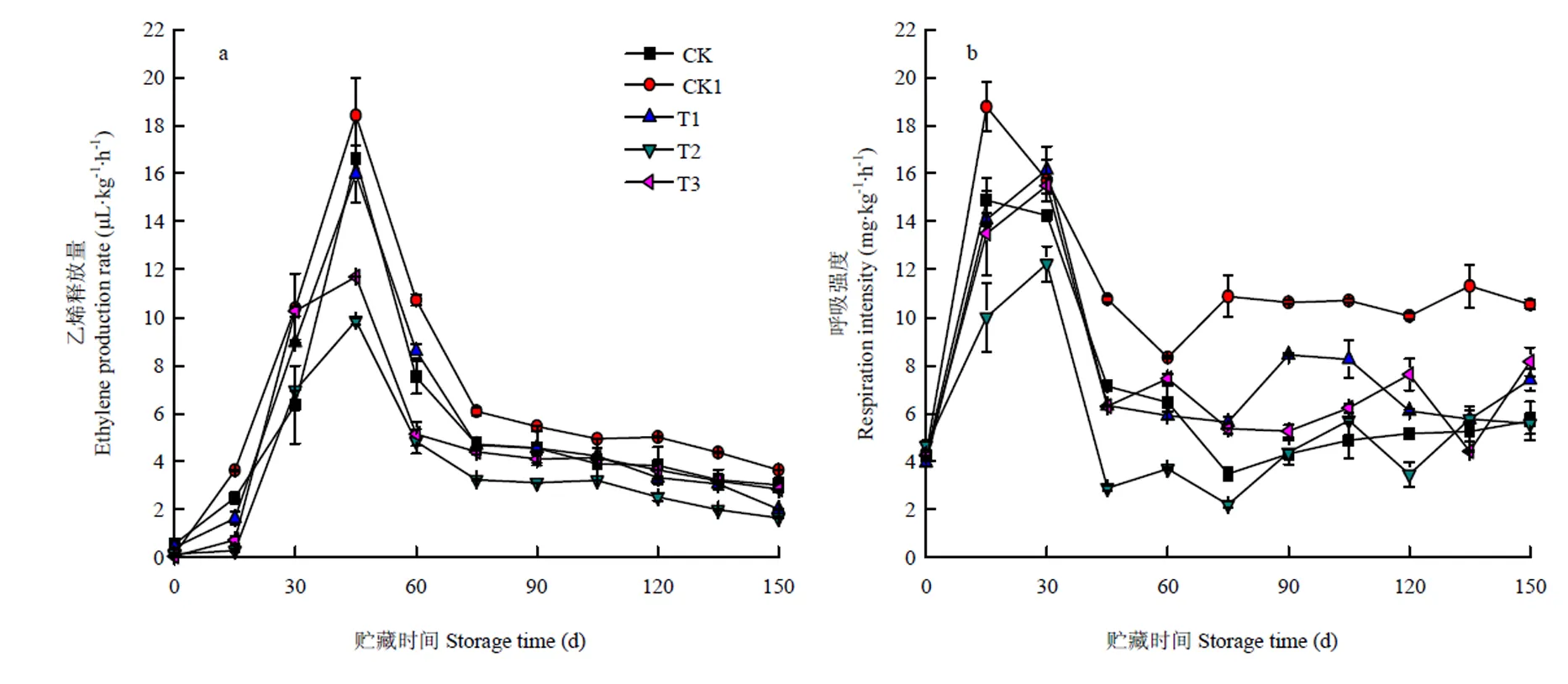
图5 O3对使用20 mg·L-1 CPPU秦美猕猴桃乙烯释放量(a)、呼吸强度(b)的影响
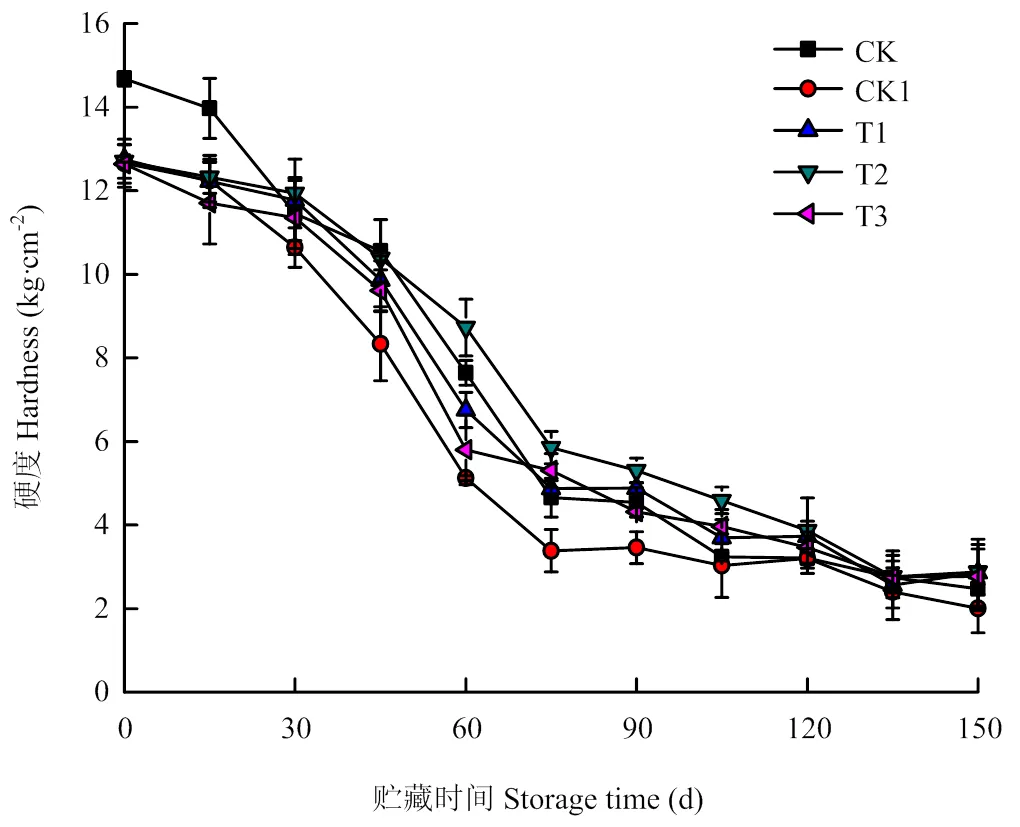
图6 O3对使用20 mg·L-1 CPPU秦美猕猴桃硬度的影响
3 讨论
乙烯与果实生长发育及其成熟衰老过程有着密切关系,对影响果实生命周期的变化起着很大作用。在采后的果蔬产品中,内源乙烯启动了体内一系列与衰老有关酶的活性,从而导致采后旺盛的新陈代谢,使果实迅速衰老变质[18]。在贮藏过程中经常采用保鲜剂来抑制乙烯代谢,环丙烯类物质1-OCP能有效抑制番茄的呼吸速率和乙烯释放量,但对贮藏过程中ACC含量影响较小,并在贮藏后期对果实带来了一定的伤害[19];Tassoni等[20]研究发现1-MCP抑制了乙烯的生成和ACS的表达,推迟了番茄的成熟,但对ACO影响不大。而O3与其他保鲜剂相比具有无污染、无残留等优点,是一种新型的绿色保鲜剂,并能很好的抑制呼吸速率、乙烯释放量等。
果农为了增加猕猴桃产量,采前使用CPPU处理,虽然增加了果实的单果重,但对猕猴桃贮藏带来了消极影响。宋小青等[21]研究发现CPPU处理有效促进了秦美猕猴桃果实乙烯的释放,造成果实软化加快,减少了猕猴桃的贮藏时间。而O3能使果蔬新陈代谢的有毒有害产物被氧化,从而达到延长贮藏期的效果。本试验研究发现采用适宜浓度O3处理能有效抑制使用了20 mg·L-1CPPU的秦美猕猴桃果实中内源乙烯的代谢。
猕猴桃的乙烯代谢是由Met经SAM合成酶催化生成SAM,再经过ACS催化生成ACC,最终经ACO催化生成乙烯。Met是乙烯代谢的前体物质,在贮藏中呈下降趋势,在贮藏后期,Met含量又有上升的趋势,这可能是甲硫基核糖向Met转化速率快于Met向乙烯转化速率[3]。SAM同Met含量变化趋势一致(图1-a)。这可能是因为在贮藏后期乙烯释放量减少,呼吸速率下降,消耗的Met和SAM减少。SAM合成酶催化Met向SAM转化,随着SAM合成酶活性的增加,SAM含量呈下降趋势(图1-b),这可能是因为Met(底物)含量不足,致使生成物SAM同底物Met含量变化趋势一致,但由于酶活性的增加,SAM含量相较于Met含量下降速率减缓。
Hoffman等[22]指出,ACC除了能转变成乙烯外,还能与丙二酸发生酰化形成MACC,并认为这是植物体内避免ACC积累的一种“去毒”调节。本试验中,我们发现O3处理抑制了ACC含量的积累,增加了ACC向MACC途径的转化,从而减少了乙烯的释放,延长了使用高浓度CPPU处理猕猴桃的贮藏时间。ACS是以磷酸吡哆醛为辅基的酶[23],O3处理能显著抑制ACS活性,这与MINAS[24]的研究结果相同,这可能是因为O3启动了ACS1基因的表达,从而抑制了ACS活性。ACO是乙烯合成途径中的最后一个酶,直接催化ACC合成乙烯,据KENTJ[25]研究发现,ACO可能也是乙烯生物合成途径中的一个限速酶,它在转录水平上调控着乙烯的生成速率。本文研究发现O3处理显著降低了ACO活性,这与MINAS[24]的研究结果相同,这可能是O3处理抑制了ACO1基因的表达,从而抑制了乙烯的生成。郭芹等[26]对番木瓜进行研究发现乙烯高峰出现在第6天,而ACC和ACS活性的高峰则在贮藏后第8天出现,表明ACS的活性加速ACC含量的积累,这与本试验研究结果相同。
猕猴桃是典型的呼吸跃变型果实,具有明显的乙烯释放高峰,O3处理能显著降低乙烯峰值,减少乙烯释放量,并降低果实呼吸速率,这与王玉萍等[27]的研究结果一致。O3处理不能推迟猕猴桃乙烯高峰出现的时间,但显著降低了乙烯高峰值,这是因为O3一方面能通过氧化消除库内的乙烯等有害气体,另一方面能抑制乙烯合成途径中相关酶的活性,从而减少乙烯释放量[28]。试验中还发现并非O3浓度越高越好,适宜的O3处理能有效减缓乙烯代谢,O3浓度过低起不到抑制的效果,而浓度过高产生大量氧气,使果蔬在一个富氧环境中,对乙烯代谢具有一定的促进作用。
4 结论
本研究表明O3处理能有效抑制使用了20 mg·L-1CPPU的秦美猕猴桃果实的呼吸速率;O3处理也能显著降低果实中的乙烯代谢,延缓果实的后熟和衰老,抑制了果实硬度的下降。因此,在实际贮藏中可以采用40 mg·m-3O3处理来改善因使用膨大剂而导致猕猴桃保鲜期大幅缩短的现象。
[1] 黄文俊, 钟彩虹. 猕猴桃果实采后生理研究进展. 植物科学学报, 2017, 35(4): 622-630.
HUANG W J, ZHONG C H. Research advances in the postharvest physiology of kiwifruit., 2017, 35(4): 622-630. (in Chinese)
[2] 蒋天梅, 殷学仁, 王平, 孙崇德, 徐昌杰,李鲜, 陈坤松. 乙烯调控非跃变型果实成熟衰老研究进展. 园艺学报, 2011, 38(2): 371-378.
JING T M, YIN X R, WANG P, SUN C D, XU C J, LI X, CHEN K S. Research advance in regulation of ethylene during ripening and senescence of non-climacteric fruit., 2011, 38(2): 371-378. (in Chinese)
[3] ADAMS D O, YANG S F. Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene., 1979, 76(1): 170-174.
[4] 蒋小平. 膨大剂在猕猴桃上应用的利弊. 北方果树, 2010(3): 43.
JIANG X P. Application of swelling agent in kiwifruit., 2010(3): 43. (in Chinese)
[5] 蔡金术, 王中炎. 低浓度CPPU对猕猴桃果实重量及品质的影响. 湖南农业科学, 2009(9): 146-148.
CAI J S, WANG Z Y. Effect of low concentration CPPU on fruit size and quality of kiwifruit., 2009(9): 146-148. (in Chinese)
[6] 李艳杰, 孙先鹏, 郭康权, 王英. 臭氧、保鲜剂对猕猴桃贮藏保鲜效果的比较. 食品科技, 2009, 34(2): 45-48.
LI Y J, SUN X P, GUO K Q, WANG Y. Comparison of the storage effect on kiwi fruit between ozone and preservative., 2009, 34(2): 45-48. (in Chinese)
[7] TZORTZAKIS N, TAYBI T, ROBERTS R, SINGLETON I, BORLAND A, BAMES J. Low-level atmospheric ozone exposure induces protection against, with down-regulation of ethylene-, jasmonate- and pathogenesis-related genes in tomato fruit., 2011, 61(2/3): 152-159.
[8] RODONI L, CASADEI N, Concellón A, ALICIA C A R, VICENTE A R. Effect of short-term ozone treatments on tomato (L.) fruit quality and cell wall degradation., 2010, 58(1): 594-599.
[9] MINAS I S, TANOU G, BELGHAZI M, JOB D, MANGANARIS G A, MOLASSIOTIS A, VASILAKAKIS M. Physiological and proteomic approaches to address the active role of ozone in kiwifruit post-harvest ripening.2012, 63(7): 2449-2464.
[10] CRISOSTO C H, SMILANICK J L. Effects of continuous 0.3 ppm ozone exposure on decay development & physiological responses of peaches & table grapes in cold storage., 2002, 24(1): 39-48.
[11] 何靖柳, 刘继, 秦文, 董红敏, 许晓丽, 王春霞. 贮藏处理后红阳猕猴桃的品质变化. 食品工业科技, 2014, 35(8): 318-322.
HE J L, LIU J, QIN W, DONG H M, XU X L, WANG C X. Research of qualities of ‘Red Sun’ kiwifruit during postharvest period., 2014, 35(8): 318-322. (in Chinese)
[12] 李子龙, 罗安伟, 桂海龙, 夏冉秋, 谌凯, 赵能. 臭氧处理对猕猴桃汁酶活性的影响. 食品工业, 2014(8): 67-70.
LI Z L, LUO A W, GUI H L, XIA R Q, CHEN K, ZHAO N. effect of ozone treatment on the enzyme activities of kiwi-fruit juice.2014(8): 67-70. (in Chinese)
[13] HAN Q, GAO H Y, CHEN H J, FANG X J, WU W J. Precooling and ozone treatments affects postharvest quality of black mulberry () fruits., 2017, 221: 1947-1953.
[14] Kühnreich R, HOLZGRANE U. Impurity profiling of l- methionine by HPLC on a mixed mode column., 2016, 122: 118-125.
[15] 杨羽辉. S-腺苷-L-甲硫氨酸的分离纯化及其硫酸盐制备的研究[D]. 杭州: 浙江工商大学, 2014.
YANG Y H. Isolation, purification of S-adenosyl-L-methionine and preparation of SAM sulphate[D]. Hangzhou: Zhejiang Gongshang University, 2014. (in Chinese)
[16] 曹建康, 姜微波, 赵玉梅. 果蔬采后生理生化实验指导. 北京: 中国轻工业出版社, 2007.
CAO J K, JIANG W B, ZHAO Y M.. Beijing: China Light Industry Press, 2007. (in Chinese)
[17] 姚高峰, 秦秀林, 储炬, 钱江潮. 高效液相色谱法分析胞内S-腺苷-L-甲硫氨酸含量及其合成酶活性的优化研究. 化学与生物工程, 2011, 28(6): 83-86.
YAO G F, QIN X L, CHU J, QIAN J C. Optimization of determination of intracellular S-Adenosyl-L-Methionine (SAM) content and SAM synthetase activity by HPLC., 2011, 28(6): 83-86. (in Chinese)
[18] 张红星, 王廿, 韩涛. 抑制ACC合酶和ACC氧化酶对植物贮藏保鲜的影响. 食品科学, 2009, 30(9): 269-271.
ZHANG H X, WANG N, HAN T. Effects of inhibiting 1- Aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase on storage of plants., 2009, 30(9): 269-271. (in Chinese)
[19] 孙炳新, 付琳, 程顺昌, 魏宝东, 冯旭桥. 3种乙烯抑制剂对番茄采后乙烯生物合成的影响. 中国食品学报, 2014, 14(1): 192-197.
SUN B X, FU L, CHENG S C, WEI B D, FENG X Q. Effect of different ethylene inhibitors on postharvest ethylene biosynthesis of tomato fruits stored at ambient temperature., 2014, 14(1): 192-197. (in Chinese)
[20] TASSONI A, WATKINS C B, DAVIES P J. Inhibition of the ethylene response by 1-MCP in tomato suggests that polyamines are not involved in delaying ripening, but may moderate the rate of ripening or over-ripening., 2006, 57(12): 3313-3325.
[21] 宋小青, 任亚梅, 张艳宜, 师俊玲, 樊明涛. CPPU、1-MCP处理对猕猴桃贮藏品质的影响. 食品与发酵工业, 2017, 43(4):266-271.
SONG X Q, REN Y M, ZHANG Y Y, SHI J L, FAN M T. Effects of CPPU and 1-MCP treatments on kiwifruit quality during cold storage.2017, 43(4): 266-271. (in Chinese)
[22] HOFFMAN N E, YANG S F, MCKEON T. Identification of 1- (malonylamino)cyclopropane-1-carboxylic acid as a major conjugate of 1-aminocyclopropane-1-carboxylic acid, an ethylene precursor in higher plants., 1982, 104(2): 765-770.
[23] KENDE H. Ethylene biosynthesis., 2003, 44: 283-307.
[24] MINAS I S, VICENTE A R, DHANAPAL A P, MANGANARIS G A, GOULAS V, VASILAKAKIS M, CRISOSTO C H, MOLASSIOTIS A. Ozone-induced kiwifruit ripening delay is mediated by ethylene biosynthesis inhibition and cell wall dismantling regulation., 2014, 229: 76-85.
[25] KENT J B. Shang Fa Yang: Pioneer in plant ethylene biochemistry., 2008, 175(1): 2-7.
[26] 郭芹, 王吉德, 李雪萍, 陈维信, 吴斌. 一氧化氮处理对采后番木瓜果实乙烯生物合成的影响. 广东农业科学, 2013, 40(3): 75-78.
GUO Q, WANG J D, LI X P, CHEN W X, WU B. Effects of nitric oxide treatment on ethylene biosynthesis in harvested papaya fruit.2013, 40(3): 75-78. (in Chinese)
[27] 王玉萍, 段琪, 饶景萍, 李萌. 1-MCP对不同品种猕猴桃果实冷害的调控作用. 西北农业学报, 2013, 22(3): 110-114.
WANG Y P, DUAN Q, RAO J P, LI M. Regulating action of 1-Methylcyclopropene on chilling injury of different kiwifruits., 2013, 22(3): 110-114. (in Chinese)
[28] 陈存坤, 高芙蓉, 薛文通, 朱婉贞, 董成虎, 王文生. 臭氧处理对新疆厚皮甜瓜贮藏品质和生理特性的影响. 食品科学, 2016, 37(20): 215-220.
CHEN C K, GAO F R, XUE W T, ZHU W Z, DONG C H, WANG W S. Effects of ozone treatment on storage quality and physiological characteristics of Xinjiang thick-skinned melon., 2016, 37(20): 215-220. (in Chinese)
(责任编辑 杨鑫浩)
Effect of Postharvest O3Treatment on ethylene metabolism of Kiwifruit Preharvest Treated with CPPU
SU Miao1, LUO Anwei1, LI Lin1, LI Yuanyuan1, BAI Junqing1, LI Rui1, FANG Yimeng1, SONG Junqi1, LIN Zhiying1, Liu Zhande2
(1College of food science and Engineering Northwest A&F University, Yangling 712100, Shaanxi;2College of Horticulture, Northwest A&F University, Yangling 712100, Shaanxi)
【Objective】The objective of this study was to investigate whether or not the ozone (ozone, O3) could effectively reduce the negative effects of the swelling agent (N-2-chloro-4-pyridine benzene-N'- phenyl urea, CPPU) on kiwifruit, which would provide useful information for using of CPPU on the market for the storage.【Method】First, the Qinmei kiwifruit was treated with 20 mg·L-1CPPU in the growing season, then treated with 10, 40, 70 mg·m-3ozone respectively in postharvest. The material contents in kiwifruit of methionine (methionine, Met), S-adenosyl methionine (S-adenosyl methionine, SAM), 1-aminocyclopropane-1- carboxylic acid (1-aminocyclopropane-1-carboxylic acid, ACC) and the related metabolic enzyme activity of ACC synthase (ACC synthase, ACS) and ACC oxidase (ACC oxidase, ACO) were studied. 【Result】The decreasing rate of Met, SAM and activity of SAM synthase, ACS, ACO under CK (without the use of CPPU and O3treatment as CPPU control CK group) during storage was lower than that under CK1 (using CPPU without using O3treatment for ozone treatment control CK1 group). At 60 days after kiwifruit storage, the contents of Met under CK1 and each treatment group (10, 40 and 70 mg·m-3ozone) were 1.36, 2.62, 4.41 and 2.60 mg·(100 g)-1, respectively. The contents under O3treatment was significantly higher than that under CK1 (<0.05). The contents of SAM under CK1 and 40 mg·m-3O3group were 15.48 mg·(100 g)-1and 20.73 mg·(100 g)-1, respectively (<0.05), but the contents under CK1 10 mg·m-3O3treatment, and 70 mg·m-3O3treatment no significant difference (>0.05). The ACC contents under CK1 and each treatment group (10, 40 and 70 mg·m-3ozone) were 0.068, 0.059, 0.038 and 0.055 nmol·g-1, respectively. There were significant differences (<0.05) between 40, 70 mg·m-3O3treatments and CK1. The ACS activities in CK1 group and each treatment group (10, 40and 70 mg·m-3) were 0.084, 0.069, 0.054 and 0.080 nmol·(g·h)-1, respectively. The peak of ACO activity was 0.062, 0.046, 0.029 and 0.051 nmol·(g·h)-1, respectively. There was significant difference between the treatment and the CK1 (<0.05). The peak values of ethylene in CK1 and each treatment (10, 40, and 70 mg·m-3ozone) were 18.42, 15.99, 9.86 and 11.69 μL·kg-1·h-1, respectively, and the peak of respiration was 18.77, 16.15, 12.24 and 15.48 mg·kg-1·h-1, respectively.【Conclusion】CPPU increased the kiwifruit ethylene production and accelerated fruit softening, had a negative impact on kiwifruit storage, while O3could effectively inhibit the ethylene metabolism of kiwifruit. O3treatment could effectively slow down the kiwifruit ripening due to the use of CPPU.
ozone; CPPU; kiwifruit; ethylene metabolism; respiration rate
2018-03-09;
2018-05-10
陕西省农业科技创新与攻关项目(2015NY051)、陕西省重点研发计划农业领域重点项目(2018ZDXM-NY-056)、杨凌示范区产学研用协同创新重大项目(2018CXY-04)
苏苗,E-mail:745396849@qq.com。通信作者罗安伟,E-mail:luoanwei@nwsuaf.edu.cn
10.3864/j.issn.0578-1752.2018.15.0013
猜你喜欢
水泵技术(2022年1期)2023-01-14
中国南方果树(2022年4期)2022-08-03
快乐语文(2021年36期)2022-01-18
今日农业(2021年11期)2021-11-27
保鲜与加工(2021年1期)2021-02-06
石油化工技术与经济(2021年4期)2021-01-13
动漫星空(兴趣百科)(2020年12期)2020-12-12
河北果树(2020年4期)2020-11-26
作文小学中年级(2019年9期)2019-10-14
儿童故事画报·发现号趣味百科(2014年1期)2014-03-31
