
光照亏缺对橡胶-作物农林复合系统中作物生产力的影响*
2020-04-30 05:34黄坚雄周立军袁淑娜林位夫
中国生态农业学报(中英文) 2020年5期
黄坚雄, 潘 剑, 周立军, 袁淑娜,4, 林位夫
光照亏缺对橡胶-作物农林复合系统中作物生产力的影响*
黄坚雄1,2,3, 潘 剑1,3, 周立军1,2, 袁淑娜1,3,4, 林位夫1
(1. 中国热带农业科学院橡胶研究所 海口 571101; 2. 中国热带农业科学院林下资源综合利用研究中心 海口 571101; 3. 农业农村部儋州热带作物科学观测试验站 儋州 571737; 4. 省部共建国家重点实验室培育基地-海南省热带作物栽培生理学重点实验室 海口 571101)
为探明农林复合系统中光照亏缺对不同间作作物生产力的影响, 以橡胶-豆薯和橡胶-花生农林复合模式为研究对象, 测定光照强度、作物生物量、光合生理等指标, 研究光照亏缺对不同喜光间作作物生产力的影响及可能的影响因素。结果表明, 复合系统中间作区域的光照强度日变化呈单峰型, 间作区域平均光照强度为裸地对照的51.3%~59.0%。间作作物的产量显著低于相应单作处理(<0.05); 间作豆薯产量可达22.9 t×hm–2, 为单作处理的85.3%; 而间作花生产量仅为单作处理的39.2%。光照亏缺环境对间作豆薯和花生不同位置叶片的RuBisCO活性没有显著影响, 但显著提高叶绿素(特别是叶绿素b)含量, 降低类胡萝卜素含量(<0.05), 其光合能力亦低于相应单作处理。复合系统中间作豆薯叶面积高于单作豆薯, 而间作花生叶面积则较单作花生显著降低59.1%~88.4%(<0.05)。综上所述, 豆薯比花生更适合在橡胶-作物农林复合系统中进行间作, 间作豆薯应该是通过维持较高的叶面积来捕获光能并制造更多的光合产物, 从而维持相对较高的生产力。
农林复合模式; 橡胶林; 间作作物; 光照亏缺; 生物量; 光竞争; 光合生理; 叶面积
天然橡胶是我国四大战略物资之一, 主要产自巴西橡胶树(), 我国主要在海南和云南两省种植, 是我国热带地区最主要的人工林。据统计, 2017年我国橡胶种植面积居世界第3位, 达115.9万hm2[1-3]。我国于20世纪五六十年代开始大规模种植橡胶, 80年代橡胶间作有较大规模发展。多年来, 橡胶间作生产主要是在定植后3~4年内的幼龄期, 后期由于封行出现持续阴蔽环境, 导致间作喜光作物长势极弱甚至渐渐枯萎死亡, 因此橡胶林下仅适宜间作少数非常耐阴的作物, 且间作面积较小, 因此长达二三十年橡胶生产期间的林下土地资源得不到充分利用[4-6]。以往生产上曾采用常见的橡胶树品种建立宽窄行模式, 以期达到避免持续阴蔽的林下环境, 但由于橡胶树树干枝条仍会渐渐向宽行延伸倾斜, 在成龄后几年内即可封行, 达不到避免持续阴蔽的作用[4,7], 国外长达17年的研究亦得出类似的结果[5]。橡胶育种周期长达30年以上, 导致可选择的品种较少, 直到1999年, 我国通过了直立单干型品种‘CATAS 7-20-59’的审定, 并进一步开展了通过种植配置改善微环境进行间作的相关研究[4]。通过选择直立橡胶品种并采用宽窄行种植的形式, 可避免橡胶树成龄后宽行中的间作区域出现持续阴蔽的环境(可保证有2~4 h的阳光直射), 避开橡胶树根系密集分布区域。尽管光照不足仍是该种植模式的主要限制因子, 但比常规胶园已明显改善了光照条件, 有利于在整个橡胶生产周期中间作相对较多类型的作物, 是一种值得推广的农林复合模式[1,8], 该技术模式在“十三五”期间入选了农业农村部主推技术目录。
农林复合模式是间作的一种重要形式, 搭配的常见短期作物有玉米()、大豆()、花生()、小麦()、丹参()、辣椒()等喜光作物[9-13]。光是影响植物光合产物合成和累积的主要因素之一, 是其生长发育和生理代谢的重要生态因子[14-15]。许多研究表明, 在农林复合系统中, 光照亏缺是限制间作作物生长的重要因素之一[16-18], 可导致农林复合系统中玉米、小麦、大豆等作物产量和生物量显著下降[19-23], 相应地, 净光合速率、光合色素含量、穗数、叶面积、核酮糖二磷酸羧化酶(RuBisCO)活性等与作物生产力息息相关的生理形态指标亦会受到影响, 如: 杨(spp.)-粮复合系统中, 较高的遮阴度会导致玉米和小麦光合速率降低, 减少作物穗数和千粒重; 南酸枣()-花生农林复合系统中, 光照不足使花生产量降低, 单株叶面积大幅减少[21,24-26]。
前期的研究结果表明, 与单作处理相比, 橡胶-作物农林复合模式中玉米、柱花草()、大豆、豆薯()等不同喜光间作作物之间的产量潜力差异较大, 如间作玉米可绝产, 而间作豆薯产量(等种植面积下)可达单作豆薯的94%[1,8,27]。在该橡胶-作物系统的光照条件下, 不同作物生产潜力存在较大差异的机制尚不清楚, 明确影响不同作物生产力的主要相关因素, 对推广橡胶-作物农林复合模式具有重要的理论和指导意义。因此, 本研究以生产力较高的豆薯和较低的花生为例, 测定其光合潜力、光合色素含量、叶面积变化、光合酶活性差异, 从光合生理的角度揭示该模式下不同作物对光照亏缺的响应机制, 以明确光照亏缺对不同作物生产力的影响。
1 材料与方法
1.1 试验地概况
试验地位于中国热带农业科学院试验场三队。该地年均气温20.8~26.0 ℃, 年降雨量约1 600 mm。橡胶树为宽窄行种植, 宽行行距为20 m, 窄行行距为4 m, 株距为2 m, 密度为420株×hm–2, 橡胶树品种为‘CATAS 7-20-59’, 为直立速生品种。该林段坡向为东西向, 坡度为8 ℃, 为防止水土流失, 橡胶种植方向为南北行向。橡胶树于2002年定植, 2010年进入成龄期并开始割胶, 2017年宽行中的冠幅约5.6 m, 株高约20 m, 胶园郁蔽度约为50%。本试验于2018年(即橡胶树成龄后的第9年)开始进行, 试验前期在该胶园种植过南瓜()、豆薯、地瓜()、柱花草、玉米、马铃薯()等一年生作物。单作地点位于农林复合间作小区附近的空旷地带。单作小区和间作小区的土壤质地均为粉砂黏壤土, 试验前0~20 cm土壤的肥力基本一致, 具体见表1[1,28]。
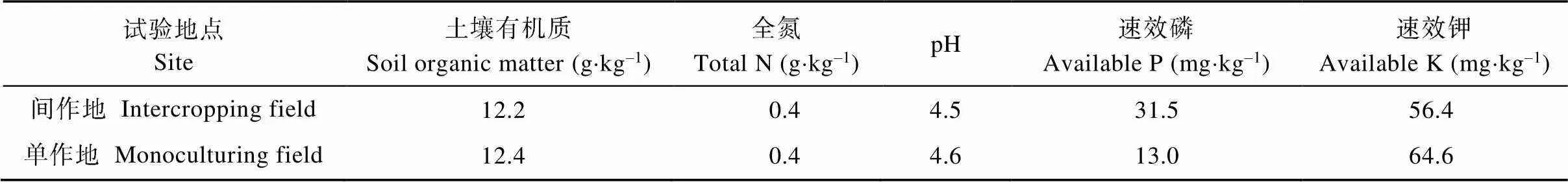
表1 橡胶-作物间作和作物单作试验地的土壤肥力
1.2 试验设计
胶园宽行长度为100 m, 行宽为20 m, 农林复合模式间作小区位于宽行中间的8.4 m区域。间作豆薯(IY)和花生(IP)小区的面积为8.4 m×8 m, 单作豆薯(Y)和花生(P)的小区面积为8 m×3 m。单作和间作处理的种植行向与橡胶树行向均为南北走向。豆薯为藤本植物, 品种为‘牧马山改良早熟1号’, 花生品种为‘琼旺2号’。单作和间作豆薯和花生的行株距分别为0.6 m×0.2 m和0.4 m×0.2 m, 间作小区豆薯和花生分别种植15行和21行。单作和间作处理均为3次重复。作物播种日期为2018年4月25日, 豆薯和花生收获日期分别为2018年9月29日和2018年7月31日。试验前以过磷酸钙、硫酸钾和牛粪做底肥, 牛粪用量为30 t×hm–2, 磷和钾肥用量分别为90 kg(P2O5)×hm–2和100 kg(K2O)×hm–2。豆薯和花生的施氮量(尿素)分别为250 kg×hm–2和180 kg×hm-2。豆薯氮肥在抽蔓期和结薯期追施, 比例为2∶3。花生氮肥在苗期和花期追施, 比例为1∶2。豆薯氮肥采用穴施方式追施; 花生苗期氮肥撒施, 花期氮肥条施并结合培土, 施肥后进行喷水。作物防虫、除草等管理均按当地生产习惯进行操作。
1.3 测定指标及方法
1.3.1 光照强度日变化
选择晴朗无云的日期测定8:00—17:00光照强度(µm×m–2×s–1)日变化。橡胶树行向为南北行向, 由于橡胶–树的遮挡, 间作小区东西侧接受光照起止时间不同, 且西侧位置接受光照的起止时间早于东侧, 故将间作区域划分成不同的观测点进行测定。从东往西方向, 分别观测小区的第0 m、4.2 m和8.4 m处的光照, 位置编号设为E、M和W(图1)。该位点对应豆薯的第1行、7行和15行, 对应花生的第1行、13行和25行, 单作小区设为CK。本试验以花生为例进行测定, 使用农业环境仪(浙江托普云农科技股份有限公司)测定光照强度, 测定3次重复, 每小时测定1次, 测定高度为离地50 cm, 测定时间为7月中旬。
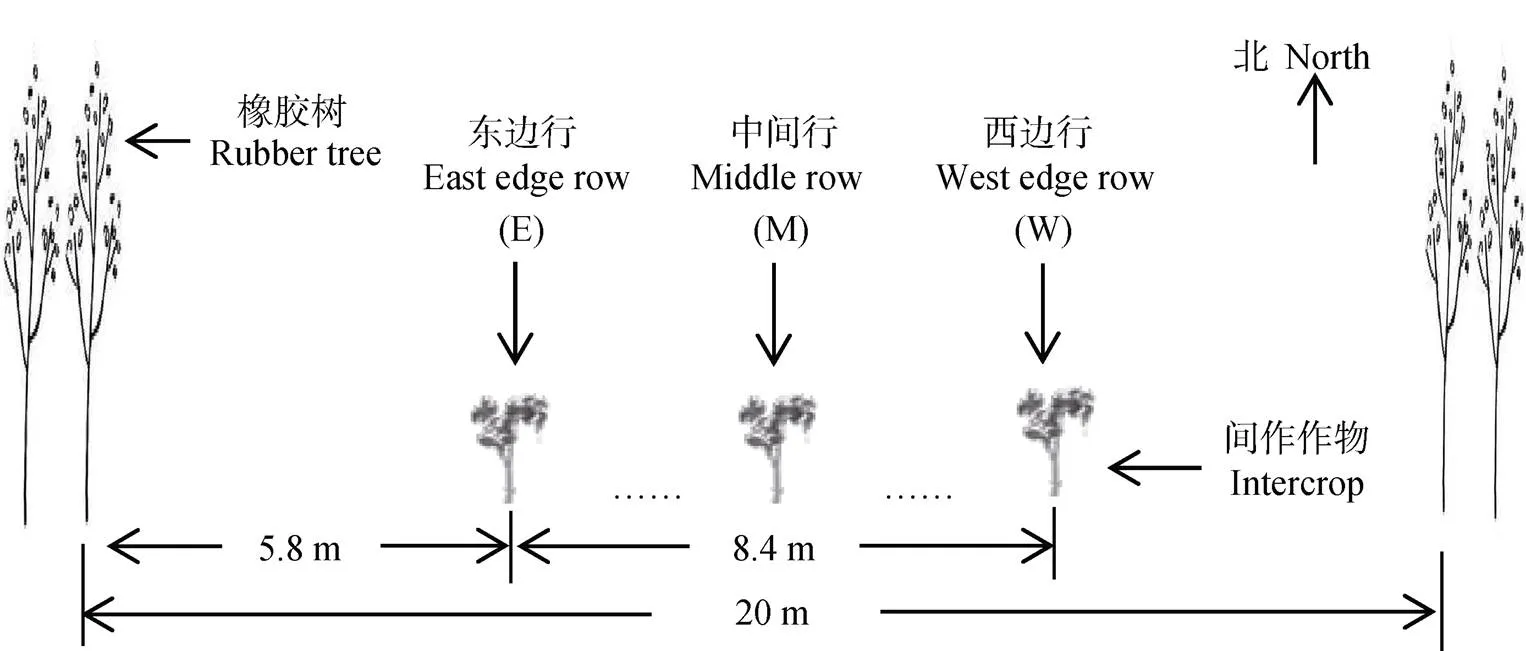
图1 农林复合系统种植与观测位置(E、M和W)示意图
1.3.2 生物量及产量
于豆薯和花生成熟期, 在间作小区中选择垂直于种植行向, 大小为2 m×8.4 m的亚区收获豆薯和花生的地上和地下部分, 用于测定生物量及产量。收获的作物测定总鲜重后取部分样品进行烘干, 用于折算单位面积生物量和产量。同时, 在与测定光照强度位置对应的种植行上分别收获豆薯和花生5穴, 用于测定复合系统中间作处理(I)不同位置(E、M和W)的作物生物量和产量。同理, 分别在单作豆薯和单作花生小区收获1 m×8 m的亚区, 并收获5穴植株, 用于测定相应的生物量和产量。其中, 豆薯测定的产量为鲜重产量, 花生则为干重产量。豆薯和花生植株地上和地下部分杀青后在85 ℃下烘至恒重后测定两种作物的生物量及花生产量。
1.3.3 叶绿素含量
于豆薯的苗期、发棵期、膨大期和成熟期, 和花生苗期、下针期、果荚期和成熟期, 分别选取不同处理不同位置(位置和编号同1.3.2)剪取稳定的10片倒3或倒4叶, 用泡沫箱加冰带回实验室, 剪碎叶片混匀, 称取0.2 g叶片放入盛有乙醇(95%,/)的试管后定容至25 mL, 避光放置24 h, 之后分别测定665 nm、649 nm和470 nm下的吸光度, 根据赵世杰等[29]的方法计算出不同光合色素含量。
1.3.4 光响应曲线
测定不同处理豆薯和花生的光响应曲线差异, 用于分析其光合能力。使用Li-6400XT于作物生长旺盛时期, 即豆薯膨大期和花生果荚期, 分别选取不同处理3个小区不同位置(位置和编号同1.3.2)的正常豆薯和花生植株5株, 测定豆薯和花生稳定的倒3或倒4叶的光响应曲线, 测定时设置流速为500 mol×s–1, 温度为30 ℃, 相对湿度为70%, CO2浓度为400 μmol×mol–1, 测定豆薯的光强梯度(µmol×m–2×s–1)为2 200、2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、20、0, 测定花生的光强梯度为2 000、1 800、1 500、1 200、1 000、800、600、400、200、100、50、20、0。测定时间为上午8:00—11:30。
1.3.5 叶面积
于作物生长旺盛时期, 即豆薯膨大期和花生果荚期, 分别选取不同处理不同位置(位置和编号同1.3.2)的正常豆薯和花生植株5穴, 将所有叶片摘下混匀, 随机选取100片叶片, 用打孔器取下样品叶片, 分别将所有叶片和样品叶片烘干后用万分之一天平进行称重, 根据打孔器直径和样品叶片重量折算叶面积。
1.3.6 RuBisCO活性
RuBisCO是最主要的光合酶, 其酶活性高低与光合速率大小密切相关。于作物生长旺盛时期, 即豆薯膨大期和花生果荚期, 分别选取不同处理不同位置(位置和编号同1.3.2)剪取稳定的10片倒3或倒4叶, 用泡沫箱加冰带回实验室, 剪碎混匀后称取0.2 g叶片, 加2 mL PBS缓冲液(0.01 mol×L–1, pH=7.4)在冰浴下研磨, 用6 mL PBS缓冲液冲洗至离心管, 在高速离心机上离心后(10 000转×min–1)提取上清液后保存于-20 ℃冰箱。RuBisCO活性采用酶联免疫吸附试验(ELISA)测定, 试剂盒(进口)采购自上海酶联生物科技有限公司, 根据试剂盒说明进行测定操作。
1.3.7 数据处理
采用Excel 2010软件和SPSS 17.0统计软件进行数据处理和统计分析, 采用最小显著差异法进行方差分析(<0.05)。
2 结果与分析
2.1 不同间作系统的作物产量和生物量
橡胶-豆薯和橡胶-花生间作系统中豆薯和花生的产量和生物量如图2所示。结果表明, 与相应的单作相比, 间作豆薯的生产潜力明显高于间作花生。尽管间作豆薯和花生的产量和生物量均显著低于相应的单作处理(<0.05), 但间作豆薯的产量(22.9 t×hm–2)和生物量(3 697 kg×hm–2)分别达单作处理的85.3%和84.4%(图2A, 2C), 而间作花生的产量和生物量分别为单作处理的39.2%和29.0%(图2B, 2D)。从不同位置看, 间作豆薯中间行(M)的产量和生物量与单作豆薯无显著差异, 而东边行(E)和西边行(W)则显著低于单作豆薯(图2E, 2G); 而间作花生不同位置的产量和生物量均显著低于单作处理(图2F, 2H,<0.05)。
2.2 不同间作系统的光照强度日变化与光响应曲线
由图3可知, 橡胶-花生间作系统不同位置的光合有效辐射呈单峰曲线, 且单作处理(CK)光照强度明显比间作处理不同位置高。间作区域E、M和W位置日平均光照强度分别为609.1 µmol×m–2×s–1、700.7 µmol×m–2×s–1和688.7 µmol×m–2×s–1, 分别为CK的51.3%、59.0%和58.0%, E和W处光照强度低于M处。中午时段(12:00—13:00)间作区域未受橡胶树遮挡时, 其光照强度与CK相近; 受遮挡时, 对应时间段CK的光照强度则在382.1~1 577.7 µmol×m–2×s–1, 而间作区域光照强度仅在60.7~171.7 µmol×m–2×s–1, 平均光照强度仅为对照的11.8%~15.7%, 显著低于CK(<0.05)。

图2 不同处理及不同位置豆薯(A, C, E, G)和花生(B, D, F, H)的产量和生物量差异
CK指单作, I指间作, E、M和W分别指间作小区的东、中和西行。不同小写字母表示各处理间(行间)差异显著(<0.05),=3. “CK” is the monocultured crop. “I” means intercropping. “E” “M” and “W” mean the intercropped crop in the east edge row, middle row and west edge row of the intercropped plot, respectively. Different lowercase letters indicate significant differences at< 0.05 level.= 3.
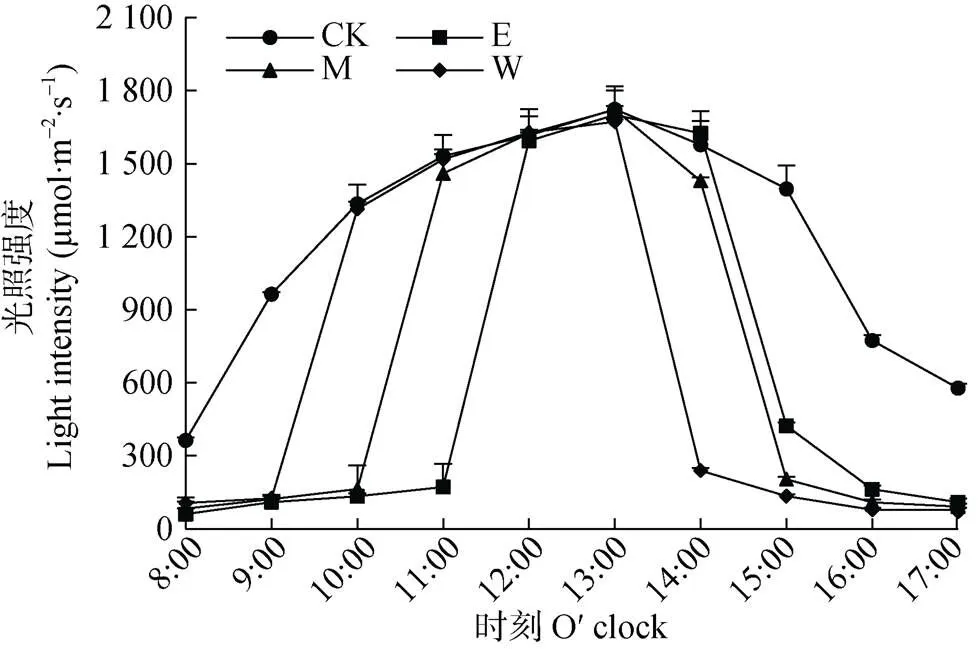
图3 与橡胶间作的花生不同观测位置(东边行, E;中间行, M; 西边行, W)和单作花生(CK)的光照强度日变化
通过比较不同处理豆薯和花生不同位置叶片光响应曲线(图4)可知, 当光照强度>200 µmol×m–2×s–1时, 间作处理的净光合速率(13.5~25.4 µmol×m–2×s–1)比单作处理(16.5~32.0 µmol×m–2×s–1)低5.0%~39.3%, 间作处理(除间作花生M外)的平均光合速率均显著低于单作处理(<0.05)。而光照强度≤200 µmol×m–2×s–1时, 单作和间作处理净光合速率则非常接近, 且无显著性差异。可见, 间作豆薯和花生不同位置叶片光合能力均低于相应单作处理。

图4 与橡胶间作的豆薯(A)和花生(B)不同观测位置(东边行, E;中间行,M;西边行,W)和相应的单作(CK)的光响应曲线
2.3 不同间作系统的叶片光合色素变化
根据间作系统豆薯和花生不同生育时期不同位置光合色素含量变化(图5和图6)可得出, 光照不足主要影响豆薯和花生叶片的叶绿素(叶绿素b)和类胡萝卜素含量。如图5所示, 不同生育时期单作处理(CK)的叶绿素a含量与间作处理不同位置的含量几乎无显著差异, 而叶绿素b含量则均显著低于间作处理不同位置的含量, 进而导致单作处理叶绿素含量均显著低于间作处理。相反, CK的类胡萝卜素含量总体上显著高于间作处理(<0.05)。类似地, 在单作和间作花生上亦有类似的变化趋势。总体而言, 农林复合系统中间作豆薯和花生总体上提高了叶绿素含量, 降低了类胡萝卜素含量, 而叶绿素含量的提高主要是由于叶绿素b含量的提高。
2.4 不同间作系统的叶面积和RuBisCO活性
由图7A可知, 与相应单作处理相比, 农林复合系统中间作豆薯和花生叶面积变化趋势明显不同。间作提高了豆薯的叶面积, 其中间作中间行的豆薯叶面积(6 086 cm2×cave–1)显著比单作高77.7%; 而间作中间行花生的叶面积反而较单作花生(47 240 cm2×cave–1)显著降低59.1%~88.4% (<0.05)。与叶面积变化趋势不同的是, 与单作处理相比, 农林复合系统中的间作豆薯和花生叶片RuBisCO活性无显著差异(图7B)。
3 讨论
农林复合系统中资源竞争是影响间作作物生产力的重要因素。现阶段的研究结果表明, 采用‘CATAS 7-20-59’橡胶的宽窄行种植模式, 在进入成龄的第9年(定植后第17年), 该模式仍可有效避免宽行出现持续阴蔽环境, 其间作区域光照日变化分布特点呈单峰式, 与苹果()-大豆(东西行向)、南酸枣-花生(东西行向)、杏树()-小麦、棉花(spp.)和玉米(南北行向)等农林复合系统的光照分布特点一致[30-32]。Reynolds等[33]在杨树()-作物农林复合系统(南北行向)2年的试验结果表明, 间作大豆和玉米的产量分别为对照处理的57.9%~62.1%和51.6%~ 69.3%。王兴祥等[34]在南酸枣-花生(东西行向)的研究结果表明, 间作花生产量仅为单作处理的44.3%~50.3%, 且离枣树越近, 花生产量越低。类似地, 在核桃()和李子()-农林复合系统中, 间作大豆和辣椒的产量亦大幅降低[16]。可见, 喜光作物通常较难在许多农林复合系统中获得相对较高的产量, 且离树越近, 间作作物产量越低。本研究中, 橡胶-花生的产量和生物量表现亦是如此, 而间作豆薯虽呈相似的趋势, 但其生产潜力明显比间作花生高, 亦比以上列举的间作系统的作物高。
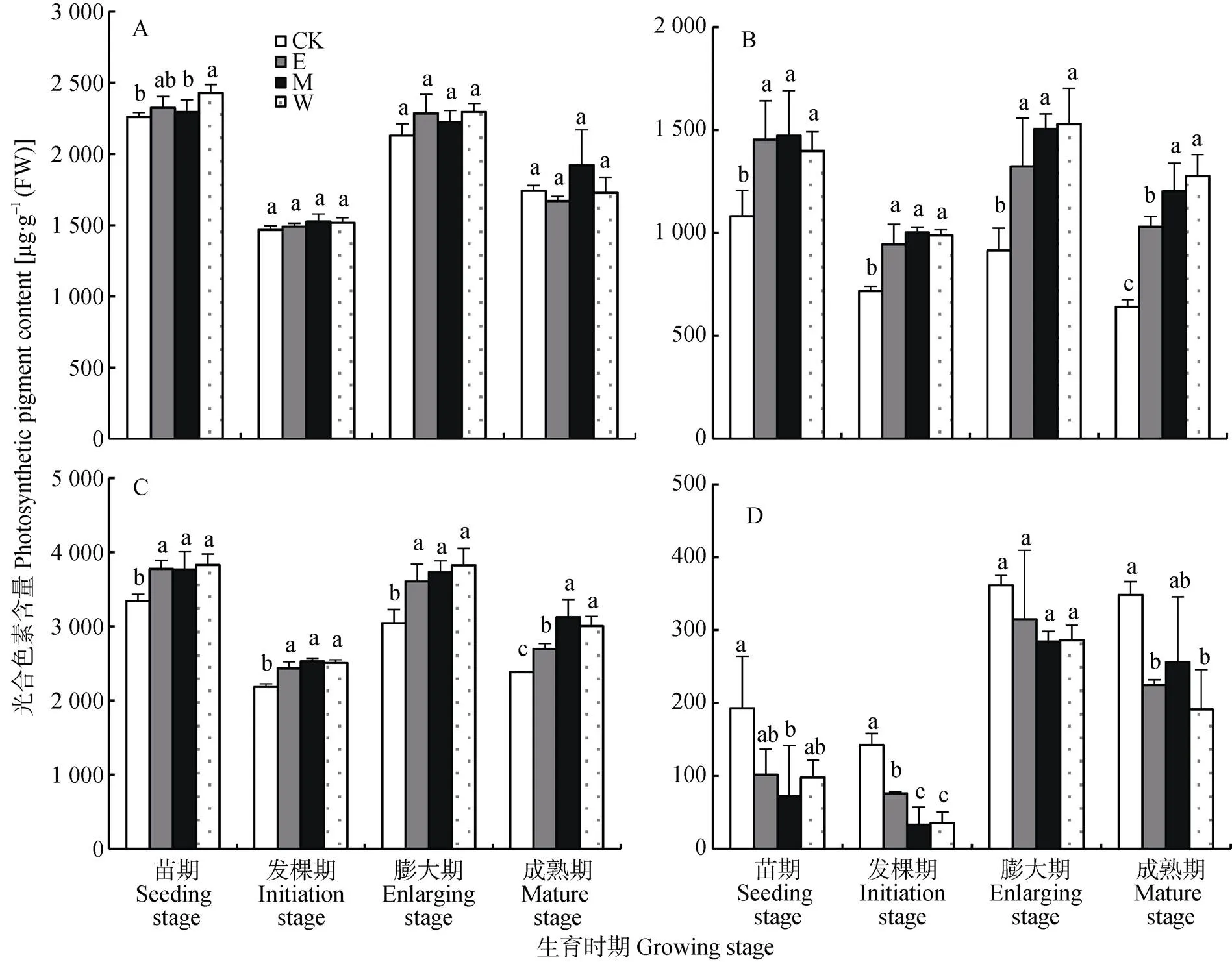
图5 不同处理不同生育时期不同位置间作豆薯光合色素含量(A:叶绿素a; B:叶绿素b; C: 总叶绿素; D:类胡萝卜素)
CK: 单作豆薯; E、M和W分别指间作小区中东、中和西行的豆薯。不同小写字母表示各处理间(行间)差异显著(<0.05),=3. “CK” is monocultured yam bean. “E” “M” and “W” mean the intercropped yam peanut in the east edge row, middle row and west edge row of the intercropped plot, respectively. Different lowercase letters indicate significant differences at< 0.05 level.= 3.
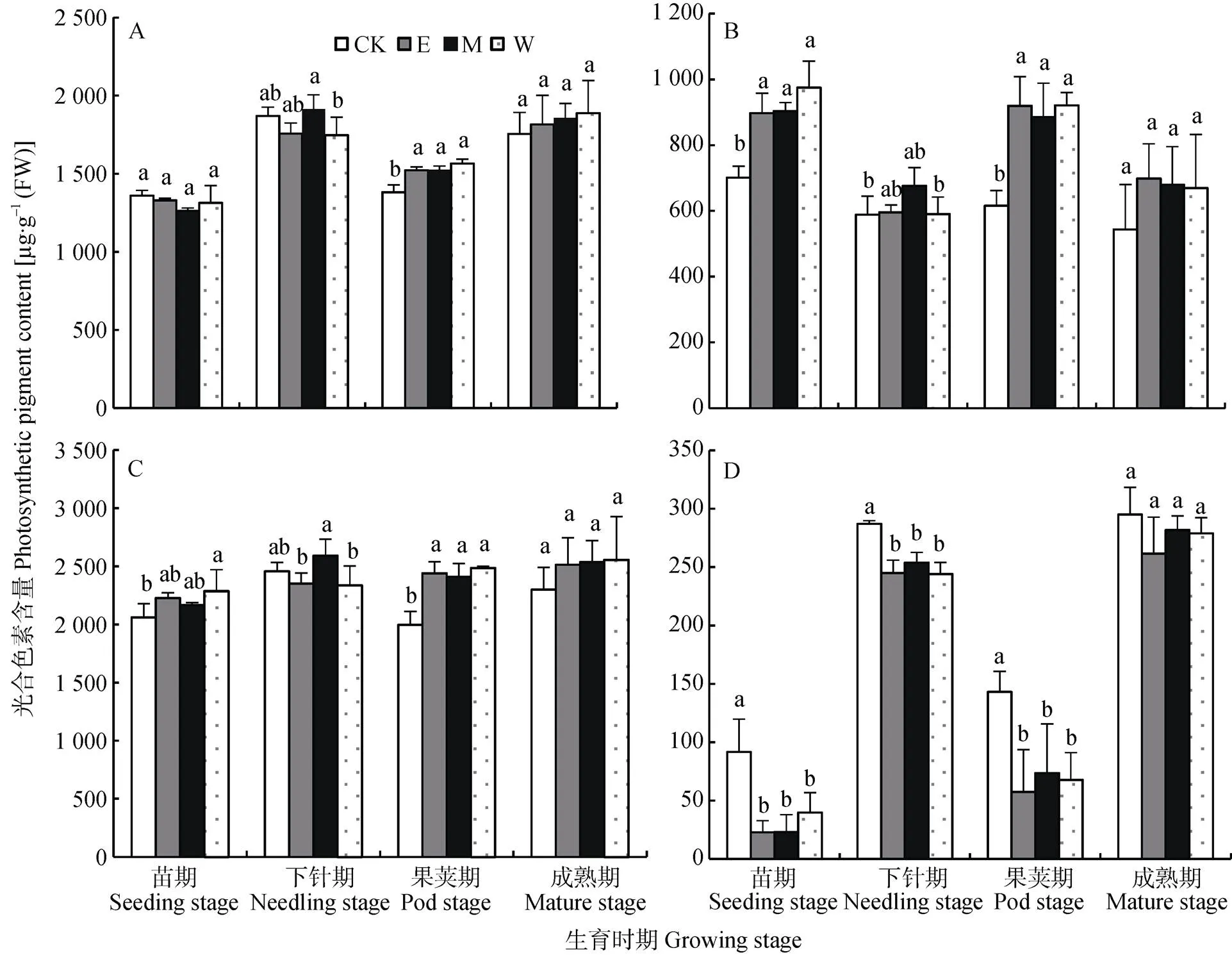
图6 不同处理不同生育时期不同位置间作花生光合色素含量(A:叶绿素a; B:叶绿素b; C总叶绿素; D:类胡萝卜素)
CK: 单作花生; E、M和W分别指间作小区中东、中和西行的花生。不同小写字母表示各处理间(行间)差异显著(<0.05),=3. “CK” is monocultured peanut. “E” “M” and “W” mean the intercropped peanut in the east edge row, middle row and west edge row of the intercropped plot, respectively. Different lowercase letters indicate significant differences at< 0.05 level.= 3.
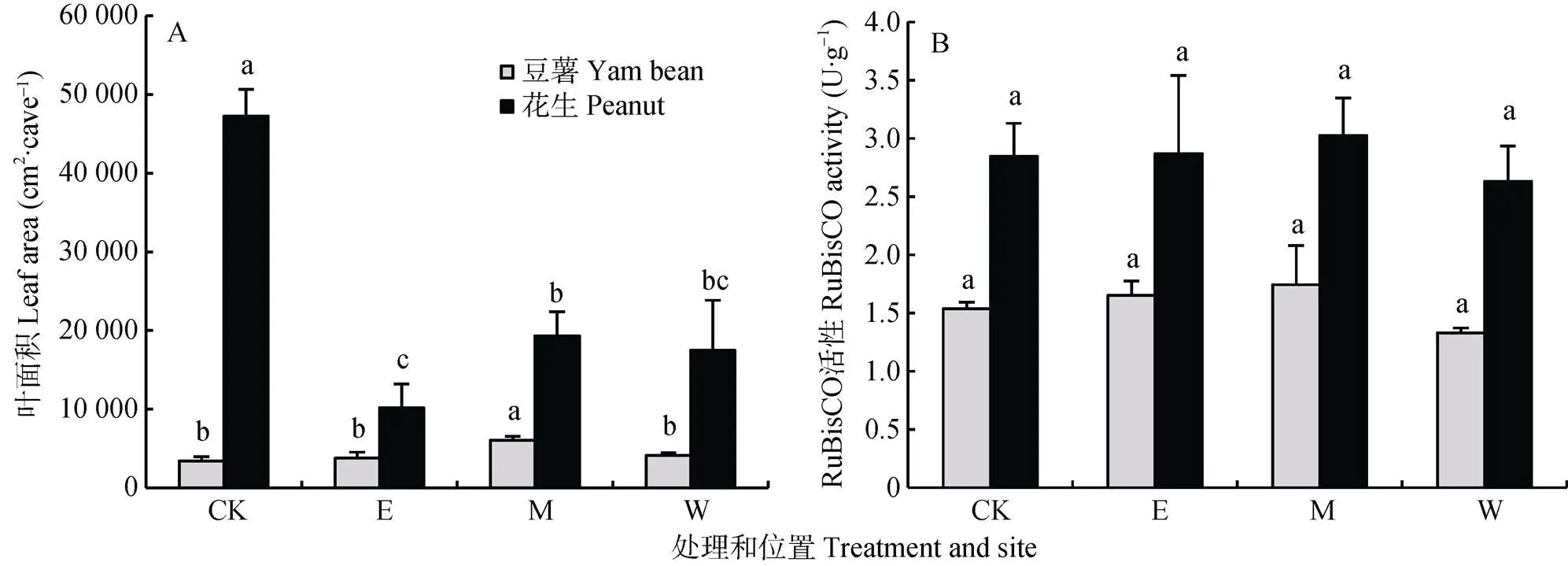
图7 与橡胶间作的豆薯和花生不同观测位置(东边行, E; 中间行, M;西边行, W)和相应单作(CK)的叶面积(A)和RuBisCO活性(B)
不同小写字母表示各处理间(行间)差异显著(<0.05),=3. Different lowercase letters indicate significant differences at< 0.05 level.=3.
光照不足可影响植物的物质代谢, 导致糖积累量的降低[35-36]。植物可通过形态及生理上的适应进行遮阴忍耐或通过垂直生长来逃避遮阴并最大化地截取光[37-38]。一些研究表明, 光照不足能促进植物提高叶绿素含量(特别提高叶绿素b来捕捉弱光环境的散射光)来增强捕光能力, 但仍导致其光合能力下降[39-42]。本研究表明间作豆薯和花生叶绿素含量变化及其光合能力与上述研究结果一致。另一方面, 在光照不足环境下不同植物叶面积和生物量变化趋势不同。如苇状羊茅()和鸭茅()两种牧草能在55%的遮阴条件下提高叶面积, 并能维持单株地上生物量[36]; 而在南酸枣-花生复合系统中, 间作花生受光照不足影响, 单株叶片数大幅减少, 叶面积降低, 其生物量和产量亦明显降低[25]。本研究得出间作豆薯能维持较大的叶面积, 而间作花生则显著降低叶面积。因此可推测, 从光合速率的角度难以说明间作豆薯和花生的生产力差异, 间作豆薯能维持较高的生产力应主要来源于叶面积的贡献。可见, 同样作为喜光作物, 豆薯比花生更能通过调节形态来适应光照亏缺的环境以达到维持较高生产力的目的。鉴于通常的喜光作物难以在当前模式下(南北行向)获得相对较高的产量, 因此建议在该模式中推广一些生产上必须有一定遮阴才能生长良好的作物, 如魔芋()(40%~70%的遮阴度非常有利生长)、山茶()(喜半阴, 忌烈日)等[43-44]。
在整个橡胶生产周期中, 开割前及初产期的叶量最大[45], 遮蔽效果最强, 而后随着生长特性、病虫害及风寒害的影响, 橡胶树叶量慢慢变少。因此可推断, 本研究中采用的模式在后期的光照环境中可以得到维持甚至改善。理论上, 该模式在东西行向种植方式下成龄后宽行的间作区域的光照表现将比南北行向种植方式更佳。该模式未出现封行现象, 由于生产周期长, 在近几年才开始大面积种植推广, 成龄后的东西行向宽窄行胶园改善光照及间作作物生产潜力的具体表现有待进一步研究。
4 结论
本研究通过测定橡胶-作物农林复合系统中喜光豆薯和花生的生产力、光照强度及作物光合生理等指标, 明确了豆薯在光照亏缺下仍能获得较高的产量潜力, 比花生更适宜在橡胶-作物农林复合系统中间作。光照亏缺环境均使豆薯和花生光合能力下降, 但叶面积变化却明显不同, 豆薯应该是通过维持较高的叶面积来捕获光能并制造更多的光合产物, 从而维持相对较高的生产力。
[1] 黄坚雄, 潘剑, 周立军, 等. 全周期间作模式胶园中间作大豆和玉米的生产力及其抗逆生理特性[J]. 中国农业大学学报, 2015, 20(5): 57–65 HUANG J X, PAN J, ZHOU L J, et al. Productivity and physiology of stress tolerance of soybean and maize intercropping in rubber plantation with paired row planting system[J]. Journal of China Agricultural University, 2015, 20(5): 57–65
[2] 宋艳红, 史正涛, 王连晓, 等. 云南橡胶树种植的历史、现状、生态问题及其应对措施[J]. 江苏农业科学, 2019, 47(8): 171–175 SONG Y H, SHI Z T, WANG L X, et al. History, current situation, ecological problems and countermeasures of rubber tree planting in Yunnan Province[J]. Jiangsu Agricultural Sciences, 2019, 47(8): 171–175
[3] 莫业勇, 杨琳. 2017年国内外天然橡胶产业发展形势[J]. 世界热带农业信息, 2018, (2): 1–3 MO Y Y, YANG L. Development situation of domestic and international natural rubber industry in 2017[J]. World Tropical Agriculture Information, 2018, (2): 1–3
[4] 林位夫, 曾宪海, 谢贵水, 等. 关于橡胶园间作的思考与实践[J]. 中国热带农业, 2011, (4): 11–15 LIN W F, ZENG X H, XIE G S, et al. Reflection and practice on intercropping in rubber plantation[J]. China Tropical Agriculture, 2011, (4): 11–15
[5] SNOECK D, LACOTE R, KÉLI J, et al. Association of hevea with other tree crops can be more profitable than hevea monocrop during first 12 years[J].Industrial Crops and Products, 2013, 43: 578–586
[6] 黄先寒, 兰国玉, 杨川, 等. 海南不同栽培模式下橡胶林灌草物种多样性研究[J]. 西北林学院学报, 2016, 31(5): 115–120 HUANG X H, LAN G Y, YANG C, et al. Shrub-grass species diversity of rubber plantations under different cultivation patterns in Hainan[J]. Journal of Northwest Forestry University, 2016, 31(5): 115–120
[7] 周光武. 橡胶树宽窄行种植形式的利弊[J]. 云南热作科技, 1992, 15(1): 16–17 ZHOU G W. Advantage and disadvantage of rubber plantation planted with double row system[J]. Journal of Yunnan Tropical Crops Science & Technology, 1992, 15(1): 16–17
[8] 林福宇. 全周期间作模式胶园的间作资源特性及其利用的初步研究[D]. 海口: 海南大学, 2014LIN F Y. Preliminary studies on the characteristics and utilizations of intercropping resources in the whole cycle intercropping rubber plantation[D]. Haikou: Hainan University, 2014
[9] 赵英, 张斌, 赵华春, 等. 农林复合系统中南酸枣蒸腾特征及影响因子[J]. 应用生态学报, 2005, 16(11): 2035–2040 ZHAO Y, ZHANG B, ZHAO H C, et al. Transpiration ofin agro-forestrial system and its affecting factors[J].Chinese Journal of Applied Ecology, 2005, 16(11): 2035–2040
[10] THEVATHASAN N V, GORDON A M. Ecology of tree intercropping systems in the north temperate region: Experiences from southern Ontario, Canada[J].Agroforestry Systems, 2004, 61(1): 257–268
[11] 廖文超, 毕华兴, 赵云杰, 等. 晋西苹果+大豆间作土壤水分分布及其对大豆生长的影响[J]. 中国水土保持科学, 2014, 12(1): 24–28 LIAO W C, BI H X, ZHAO Y J, et al. Distribution of soil moisture inandintercropping system and its impact on the soybean growth[J]. Science of Soil and Water Conservation, 2014, 12(1): 24–28
[12] 彭晓邦, 张硕新. 商洛低山丘陵区农林复合生态系统光能竞争与生产力[J]. 生态学报, 2012, 32(9): 2692–2698 PENG X B, ZHANG S X. Competition for light and crop productivity in an agro-forestry system in the hilly region, Shangluo, China[J]. Acta Ecologica Sinica, 2012, 32(9): 2692–269
[13] 张雯, 谢辉, 张平, 等. 扁桃||冬小麦间作模式下树冠结构对间作区域光环境的影响[J]. 中国生态农业学报, 2016, 24(6): 753–761 ZHANG W, XIE H, ZHANG P, et al. Effect of tree canopy structure on light condition in almond-winter wheat intercropping systems[J]. Chinese Journal of Eco-Agriculture, 2016, 24(6): 753–761
[14] 李影影, 熊耀康. 光照强度与南方红豆杉初生代谢产物相关性研究[J]. 中华中医药杂志, 2018, 33(6): 2659–2664 LI Y Y, XIONG Y K. Collection study between light intensity and primary metabolite content of(Lemee et Levl.) S. Y. Hu ex Liu[J]. China Journal of Traditional Chinese Medicine and Pharmacy, 2018, 33(6): 2659–2664
[15] 朱启林, 向蕊, 汤利, 等. 间作条件下施氮量对马铃薯光合特性的调控作用[J]. 生态学杂志, 2018, 37(5): 1391–1397 ZHU Q L, XIANG R, TANG L, et al. The effects of nitrogen application rate on photosynthetic characteristics of potato under intercropping[J]. Chinese Journal of Ecology, 2018, 37(5): 1391–1397
[16] 彭晓邦, 蔡靖, 姜在民, 等. 渭北黄土区农林复合系统光能竞争与生产力[J]. 应用生态学报, 2008, 19(11): 2414–2419 PENG X B, CAI J, JIANG Z M, et al. Light competition and productivity of agroforestry system in loess area of Weibei in Shaanxi[J].Chinese Journal of Applied Ecology, 2008, 19(11): 2414–2419
[17] 夏志光. 黑土区农林复合模式土壤水分变化规律与大豆产量研究[D]. 哈尔滨: 东北林业大学, 2009 XIA Z G. Study on soil moisture changes of the agroforestry models and soybean yield in black soil zone[D]. Harbin: Northeast Forestry University, 2009
[18] GILLESPIE A R, JOSE S, MENGEL D B, et al. Defining competition vectors in a temperate alley cropping system in the midwestern USA: 1. Production physiology[J].Agroforestry Systems, 2000, 48(1): 25–40
[19] FRIDAY J B, FOWNES J H. Competition for light between hedgerows and maize in an alley cropping system in Hawaii, USA[J].Agroforestry Systems, 2002, 55(2): 125–137
[20] MUTHURI C W, ONG C K, BLACK C R, et al. Tree and crop productivity in,and-based agroforestry systems in semi-arid Kenya[J].Forest Ecology and Management, 2005, 212(1/3): 23–39
[21] 孙尚伟, 夏新莉, 刘晓东, 等. 修枝对复合农林系统内作物光合特性及生长的影响[J]. 生态学报, 2008, 28(7): 3185–3192 SUN S W, XIA X L, LIU X D, et al. Effects of different pruning intensity on photosynthetic characters, growth and yield of crops in agroforestry[J].Acta Ecologica Sinica, 2008, 28(7): 3185–3192
[22] RIVEST D, COGLIASTRO A, VANASSE A, et al. Production of soybean associated with different hybrid poplar clones in a tree-based intercropping system in southwestern Québec, Canada[J].Agriculture, Ecosystems & Environment, 2009, 131(1/2): 51–60
[23] 段志平. 枣麦(棉)间作系统光合特性与产量形成机理研究[D]. 石河子: 石河子大学, 2018DUAN Z P. The study on the photosynthetic characteristics and yield formation mechanism in jujube/wheat (cotton) agroforestry systems[D]. Shihezi: Shihezi University, 2018
[24] 彭晓邦, 张硕新. 商洛低山丘陵区农林复合生态系统中大豆与丹参的光合生理特性[J]. 生态学报, 2013, 33(6): 1926–1934 PENG X B, ZHANG S X. Photosynthetic characteristics of soybean and salvia in an agroforestry system in the Hilly Region, Shangluo, China[J]. Acta Ecologica Sinica, 2013, 33(6): 1926–1934
[25] 王明珠, 张斌. 低丘红壤南酸枣-花生复合系统物种间水肥光竞争的研究——Ⅰ. 南酸枣与花生生育性状及产量研究[J]. 中国生态农业学报, 2003, 11(2): 44–46 WANG M Z, ZHANG B. Competitions between water, nutrient, and light oftrees and peanut in the low hilly land of red soil —Ⅰ. Growing and developing properties and their yields oftrees and peanut[J].Chinese Journal of Eco-Agriculture, 2003, 11(2): 44–46
[26] 邹长明, 王允青, 刘英, 等. 四种豆科作物的光合生理和生长发育对弱光的响应[J]. 植物生态学报, 2015, 39(9): 909–916 ZOU C M, WANG Y Q, LIU Y, et al. Responses of photosynthesis and growth to weak light regime in four legume species[J]. Chinese Journal of Plant Ecology, 2015, 39(9): 909–916
[27] 黄坚雄, 潘剑, 林位夫, 等. 全周期胶园间作对豆薯和大豆的产量、品质和土壤速效养分的影响[J]. 热带作物学报, 2016, 37(2): 247–253 HUANG J X, PAN J, LIN W F, et al. Effect of intercropping in rubber plantation with paired row planting system on yield, quality and soil rapidly available nutrient of yam bean and soybean[J]. Chinese Journal of Tropical Crops, 2016, 37(2): 247–253
[28] HUANG J X, CHEN Y Q, PAN J, et al. Carbon footprint of different agricultural systems in China estimated by different evaluation metrics[J].Journal of Cleaner Production, 2019, 225: 939–948
[29] 赵世杰, 史国安, 董新纯. 植物生理学实验指导[M]. 北京: 中国农业科学技术出版社, 2002 ZHAO S J, SHI G A, DONG X C. Techniques of Plant Physiological Experiment[M]. Beijing: China Agricultural Science and Technology Press, 2002
[30] 高国治, 王明珠, 张斌. 低丘红壤南酸枣-花生复合系统物种间水肥光竞争的研究——Ⅱ. 南酸枣与花生利用光能分析[J]. 中国生态农业学报, 2004, 12(2): 92–94 GAO G Z, WANG M Z, ZHANG B. Competition of the light, fertilizer and water betweentrees and peanut in the red soil of low hilly land — Ⅱ. Analysis of using light energy oftrees and peanut[J].Chinese Journal of Eco-Agriculture, 2004, 12(2): 92–94
[31] 张建雄. 南疆绿洲灌区杏农复合系下作物生理特性及生态效应研究[D]. 杨陵: 西北农林科技大学, 2010 ZHANG J X. Study on the ecological effect and physiological characteristics of micro-climate in apricot-crop intercropping system of the Oasis irrigation district in South of Xinjiang[D]. Yangling: Northwest A&F University, 2010
[32] 廖文超, 毕华兴, 高路博, 等. 苹果-大豆间作系统光照分布及其对作物的影响[J]. 西北林学院学报, 2014, 29(1): 25–29LIAO W C, BI H X, GAO L B, et al. Light distribution in apple-soybean intercropping and its impact on the crops[J]. Journal of Northwest Forestry University, 2014, 29(1): 25–29
[33] REYNOLDS P E, SIMPSON J A, THEVATHASAN N V, et al. Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada[J].Ecological Engineering, 2007, 29(4): 362–371
[34] 王兴祥, 张斌, 王明珠, 等. 低丘红壤复合农林系统光能竞争与生产力——以花生南酸枣间作为例[J]. 生态学杂志, 2002, 21(4): 1–5 WANG X X, ZHANG B, WANG M Z, et al. Competition for light and crop productivity in agro-forestrial systems in hilly red soil region-a case study ofwith[J].Chinese Journal of Ecology, 2002, 21(4): 1–5
[35] 周兴元, 曹福亮. 遮荫对假俭草抗氧化酶系统及光合作用的影响[J]. 南京林业大学学报: 自然科学版, 2006, 30(3): 32–36 ZHOU X Y, CAO F L. Effects of shading on the antioxidant enzymatic system and photosynthesis of centipedegrass[J]. Journal of Nanjing Forestry University: Natural Science Edition, 2006, 30(3): 32–36
[36] 李志刚, 侯扶江, 安渊. 不同光照强度对三种牧草生长发育的影响[J]. 中国草地学报, 2009, 31(3): 55–61 LI Z G, HOU F J, AN Y. Effect of different light intensities on growth and development of three kinds of forages[J]. Chinese Journal of Grassland, 2009, 31(3): 55–61
[37] 陈志成, 刘晓静, 刘畅, 等. 锐齿栎幼苗生长、光合作用和非结构性碳对遮阴和模拟光斑的响应[J]. 生态学杂志, 2017, 36(4): 935–943 CHEN Z C, LIU X J, LIU C, et al. Responses of growth, photosynthesis and nonstructural carbohydrate ofvar.seedlings to shading and simulated sunfleck[J]. Chinese Journal of Ecology, 2017, 36(4): 935–943
[38] HENRY H A L, AARSSEN L W. Inter- and intraspecific relationships between shade tolerance and shade avoidance in temperate trees[J].Oikos, 2001, 93(3): 477–487
[39] 张元帅, 冯伟, 张海艳, 等. 遮阴和施氮对冬小麦旗叶光合特性及产量的影响[J]. 中国生态农业学报, 2016, 24(9): 1177–1184 ZHANG Y S, FENG W, ZHANG H Y, et al. Effects of shading and nitrogen rate on photosynthetic characteristics of flag leaves and yield of winter wheat[J]. Chinese Journal of Eco-Agriculture, 2016, 24(9): 1177–1184
[40] 宋航, 杨艳, 周卫霞, 等. 光、氮及其互作对玉米光合特性与物质生产的影响[J]. 玉米科学, 2017, 25(1): 121–126 SONG H, YANG Y, ZHOU W X, et al. Effects of light, nitrogen and their interaction on photosynthetic traits and matter accumulation of maize[J]. Journal of Maize Sciences, 2017, 25(1): 121–126
[41] KUME A, AKITSU T, NASAHARA K N. Why is chlorophyll b only used in light-harvesting systems?[J].Journal of Plant Research, 2018, 131(6): 961–972
[42] 杨顺国, 郭贵敏, 王仙萍. 遮阴对花生生长发育及产量的影响[J]. 安徽农业科学, 2015, 43(35): 30–31 YANG S G, GUO G M, WANG X P. The influence of shading on the peanut growth and yield[J]. Journal of Anhui Agricultural Sciences, 2015, 43(35): 30–31
[43] 张东华, 汪庆平. 珠芽魔芋热区(橡胶)林下套种模式展望[J]. 热带农业工程, 2016, 40(3): 21–27 ZHANG D H, WANG Q P. Perspective estimation ofplantation under rubber trees in tropical areas[J]. Tropical Agricultural Engineering, 2016, 40(3): 21–27
[44] 陈鑫, 李桃祯, 王凌晖, 等. 不同郁闭度下山茶生长及光合特性差异[J]. 广西林业科学, 2017, 46(2): 160–164 CHEN X, LI T Z, WANG L H, et al. The differences of growth and photosynthetic characteristics ofunder different canopy densities[J]. Guangxi Forestry Science, 2017, 46(2): 160–164
[45] 陈帮乾, 吴志祥, 杨川, 等. 海南儋州地区橡胶林叶面积指数月动态研究[J]. 热带农业科学, 2015, 35(11): 1–6 CHEN B Q, WU Z X, YANG C, et al. Monthly dynamics of leaf area index of rubber plantation in Danzhou, Hainan Island, China[J]. Chinese Journal of Tropical Agriculture, 2015, 35(11): 1–6
Effect of light deficiency on productivity of intercrops in rubber-crop agroforestry system*
HUANG Jianxiong1,2,3, PAN Jian1,3, ZHOU Lijun1,2, YUAN Shuna1,3,4, LIN Weifu1
(1. Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China; 2. Research Center for Utilization of Under-forest Resources, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China; 3. Danzhou Investigation & Experiment Station of Tropical Crops, Ministry of Agriculture and Rural Affairs, Danzhou 571737, China; 4. State Key Laboratory Incubation Base for Cultivation & Physiology of Tropical Crops, Haikou 571101, China)
The objective of current study was to reveal the effect of light deficiency on the productivity of different intercrops in an agroforestry system. Based on rubber-yam bean and rubber-peanut agroforestry systems, the light intensity, crop biomass, photosynthetic physiology, and other parameters were determined to investigate the effect of light deficiency on the productivity of different sun-like intercrops and potential influencing factors. The daily light intensity was unimodal in the intercropped area in the agroforestry system. The average light intensity accounted for 51.3%-59.0% of bare land control. Yield of the intercrops was significantly lower than the yield of monocultural treatments (< 0.05). However, the yield of intercropped yam bean was 22.9 t×hm–2and reached 85.3% of the yield of monocultural yam beam, while intercropped peanut was only 39.2% that of monocultural peanut. There was no significant effect of light deficiency on ribulose-1,5-bisphosphate carboxylase/oxygenase activity of intercropped yam bean and peanut in different sites. However, the contents of chlorophyll (especially chlorophyll b) of intercropped yam bean and peanut were significantly increased, and the content of carotenoid was significantly reduced compared to monocultural treatments (< 0.05). Their photosynthesis rate also was decreased compared to monocultural treatments. Nevertheless, the leaf area of intercropped yam bean in the agroforestry system was higher than monocultural yam bean, while that of the intercropped peanut was significantly reduced by 59.1%-88.4% (< 0.05). The results conclusively indicate that yam bean is more suitable than peanut as an intercrop in the rubber-based agroforestry system. Intercropped yam bean could maintain relatively higher productivity, probably due to increased photosynthesis, by increasing leaf area to capture light.
Agroforestry system; Rubber forest; Intercrop; Light deficiency; Biomass; Light competition; Photosynthetic physiology; Leaf area
, HUANG Jianxiong, E-mail: lmc172@163.com
Dec. 6, 2019;
10.13930/j.cnki.cjea.190858
黄坚雄, 潘剑, 周立军, 袁淑娜, 林位夫. 光照亏缺对橡胶-作物农林复合系统中作物生产力的影响[J]. 中国生态农业学报(中英文), 2020, 28(5): 680-689
HUANG J X, PAN J, ZHOU L J, YUAN S N, LIN W F. Effect of light deficiency on productivity of intercrops in rubber-crop agroforestry system[J]. Chinese Journal of Eco-Agriculture, 2020,28(5): 680-689
S59
* 国家自然科学基金青年科学基金项目(31701388)和现代农业产业技术体系建设专项资金项目(CARS-34-YZ4)资助
黄坚雄, 主要从事橡胶林下间作技术与理论方面的研究。E-mail: lmc172@163.com
2019-12-06
2020-01-31
* This study was supported by the National Natural Science Foundation of China (31701388) and the Special Fund for the Industrial Technology System Construction of Modern Agriculture of China (CARS-34-YZ4).
Jan. 31, 2020
猜你喜欢
声屏世界(2022年17期)2022-12-18
声屏世界(2022年15期)2022-11-08
磷肥与复肥(2022年6期)2022-08-02
北方果树(2022年1期)2022-01-21
广东蚕业(2021年4期)2021-12-02
河北农业大学学报(2021年3期)2021-07-24
辣椒杂志(2021年4期)2021-04-14
干旱地区农业研究(2020年2期)2020-06-10
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
