
丙酮丁醇梭菌(Clostridium acetobutylicum)原生质体融合育种研究
2020-07-30 09:22王欢武芳牛昆
生物技术进展 2020年4期
王欢, 武芳, 牛昆
华北制药集团新药研究开发有限责任公司, 石家庄 050035
丙酮丁醇发酵工业中的菌种主要是梭状芽孢杆菌属(Clostridium),统称丙酮丁醇梭状芽孢杆菌(Clostridiumacetobutylicum),简称丙酮丁醇梭菌。传统菌株主要产生丙酮、丁醇、乙醇,三组份的含量比为3∶6∶1。丁醇是一种重要的化工原料,主要用于制造邻苯二甲酸二丁酯和脂肪族二元酸丁酯类增塑剂,广泛用于各种塑料和橡胶制品的生产。丁醇还可用来生产丁醛、丁酸、丁胺和醋酸丁酯,它们可用作树脂、油漆、粘接剂的溶剂,也可用作油脂、药物和香料的萃取剂及醇酸树脂涂料的添加剂。近年来美国科学家的研究表明,丁醇还是一种极具潜力的新型生物燃料。与甲醇和乙醇相比,丁醇在性能上与汽油更为接近,因此引起了研究者和企业更多的关注[1]。
目前生产上的主要问题集中于丁醇毒性问题,丙酮丁醇梭菌自身产生丁醇,但随着发酵液中丁醇浓度的提高却又破坏了适合梭菌细胞生长的pH,降低了细胞内的ATP水平,抑制了菌体对葡萄糖的吸收[2],解决这一关键问题,就菌株而言,必须提高梭菌自身的丁醇耐受性。
原生质体融合技术是通过酶解方法来去除两个亲本的细胞壁,在渗透压很高的环境下能够释放出原生质体,利用助溶剂将两个亲本原生质体在此条件下进行融合,促使聚集,先进行异核体融合,然后进行核融合,使其交换染色体,实现基因重组,这种技术打破了微生物的种间界限,实现了非亲缘菌株的基因重组[3-4];原生质体融合技术可以将DNA 或者RNA 有效并且完整地传递,使更多的远缘物种的基因得以重组[5]。原生质体融合的特点包括:杂交频度很高,即原生质体融合频率明显高于有性杂交;限制少,即不同种属间也能杂交;遗传物质传递更完善;提高多菌株融合的可能性。姜雄韬等[6]通过原生质体融合技术获得了产细菌素能力强的融合菌株。Sun等[7]通过原生质体融合技术获得了能在高温条件下产胞外多糖的融合菌株。Worawan等[8]用原生质体融合技术选育出了由Monascus属黄突变体的融合子,用于增强黄色颜料的生产。
对于丙酮丁醇梭菌(Clostridiumacetobutylicum)的原生质体实验只涉及到原生质体制备与再生过程,而对丙酸丁醇梭菌原生质体融合的介绍尚未见报道。本研究选用的两株目前生产上使用的菌株丙酮丁醇梭菌HY1214、HY1710,对两株菌的原生质体制备、再生及融合条件进行了研究,并利用PEG(聚乙二醇)介导遗传转化将外源DNA 整合到细菌基因组中,从而改变细胞膜透性以利于DNA 进入[9]。本研究希望获得丁醇耐受性较高的融合菌株,为后续研究提供参考。
1 材料和方法
1.1 材料
1.1.1菌株 丙酮丁醇梭菌(Clostridiumacetobutylicum)HY1214 (可耐受13 g·L-1丁醇)及HY1710(可耐受10 g·L-1丁醇)生产菌株,由本实验室保存。
1.1.2试剂和酶 融合试剂50%(体积分数)PEG1000、PEG4000、PEG6000(鼎国公司);PPM:葡萄糖10 g,无水硫酸镁0.08 g,一水硫酸锰0.006 3 g,七水硫酸亚铁0.008 3 g,磷酸氢二钾0.784 g,磷酸二氢钾0.224 g,水解酪蛋白0.8 g,对氨基苯甲酸0.001 g,生物素2 μg,蔗糖0.3 mol·L-1,二水氯化钙50 mmol·L-1,六水氯化镁50 mmol·L-1,调pH到7.5[10];溶菌酶(Sigma公司)。
1.1.3培养基 RCM培养基:酵母提取物3 g,牛肉膏l0 g,蛋白胨10 g,可溶性淀粉1 g,葡萄糖5 g, 半胱氨酸盐酸盐0.5 g,NaCl 3 g,NaAc 3 g, 定容到l 000 mL[11];SRA培养基:葡萄糖10 g,无水硫酸镁0.08 g,一水硫酸锰0.006 3 g,七水硫酸亚铁0.008 3 g,磷酸氢二钾0.784 g,磷酸二氢钾0.224 g,水解酪蛋白3 g,二水氯化钙4 g,六水氯化镁5 g,酵母提取物8 g,天冬酰胺1 g,半胱氨酸0.5 g,定容到1 000 mL[10];PDA培养基 :马铃薯6.0 g,葡萄糖 20.0 g[11],用于梭菌培养和再生;6%玉米醪糟培养基:60 g玉米面熬制30 min,定容到1 000 mL,用于发酵种子的制备;8%玉米醪糟培养基用于发酵验证;5%玉米醪糟平板加入1.5%的琼脂,用于丁醇耐受菌筛选。
1.2 实验方法
1.2.1原生质体制备、融合、再生条件的选择
首先将计划融合的两个生产菌株,在厌氧条件下,使用溶菌酶进行破壁,制备原生质体;将其中1株梭菌的原生质体在55 ℃下致死;再将两株菌的原生质体进行融合,使其基因组能够产生随机的置换或重组;对融合的原生质体进行再生,从而得到遗传性状有所改变的融合子[12-17]。
实验对原生质体的制备、融合、再生条件设置梯度筛选最优条件。①原生质体制备条件筛选。酶作用温度37℃,破壁浓度为2、3、4、5 mg·mL-1,破壁时间为10、20、30、40、50、60 min,分别进行镜检,破壁完成后使用细菌计数板对球形原生质体进行计数[11-16];②再生培养基种类筛选。本实验选用了RCM、PDA、SRA固体培养基作为原生质体再生培养基[14],将丙酮丁醇梭菌使用4 mg·mL-1的溶菌酶作用30 min,涂于3种培养基的平板上,以确定各种培养基再生能力的强弱。将经过PEG诱导融合的菌体使用PPM洗净后,以相同的量接种于RCM、PDA、SRA固体培养基上,厌氧箱中,37 ℃培养2~3 d后对平板上的菌落进行计数;③致死时间的确定。将丙酮丁醇梭菌使用4 mg·mL-1的溶菌酶作用30 min。离心产生的油状沉淀,根据实验目的将两株菌之一进行热致死,在55 ℃ 10、20、30、40、50、60 min时分别将菌体涂布于之前确定的SRA平板上[13],观察HY1214的致死时间;④PEG分子量与作用时间的确定。将55 ℃,30 min致死的HY1214原生质体与HY1710的原生质体混合均匀,分别使用50% PEG1000、PEG4000、PEG6000对原生质体诱导融合不同时间[10,15-16],离心,去上清,再使用PPM缓冲液将菌体洗涤一次,观察三种分子量的PEG作用不同时间的诱导融合能力;⑤Ca2+、Mg2+溶液的终浓度的确定。待原生质体经PEG4000 5 min诱导融合以及PPM洗净后分别加入不同量Ca2+、Mg2+的溶液(25、50、75、100、125 mmol·L-1)[5],将原生质体混匀,均匀涂布于SRA固体培养基上。厌氧箱中,37 ℃培养2~3 d后对平板上的菌落进行计数。通过这些条件的变化对丙酮丁醇梭菌原生质体制备、融合、再生的影响,来得到合适的原生质体操作条件。
1.2.2融合子的筛选 融合再生后,将平板上的菌落洗下后接种于含有13、15、18 g·L-1丁醇的液体SRA培养基中进行耐丁醇梯度筛选,厌氧条件下,37 ℃培养16 h,将有菌体生长的发酵液离心后,用余液将菌体悬起,涂布于5%玉米醪糟平板上,厌氧箱中,37 ℃培养2~3 d,挑选透明圈较大的菌株,制种后分别接入8%的玉米醪糟中进行发酵验证,通过气相色谱检测丙酮、乙醇、正丁醇产量,计算正丁醇以及总溶剂的转化率。
1.2.3残糖的测定 使用菲林试剂方法测定残糖[17],取发酵液检测发酵终点糖的利用情况。
1.2.4发酵产物检测 仪器:GC-7890A(Agilengt Technologies);色谱柱:DB-624(30 m×0.32 mm×1.8 μm,Agilengt Technologies);检测器:FID(300℃);载气:N2;温度模式:1阶程序升温,初温55 ℃,保持8 min,速率45 ℃·min-1,终温190 ℃,保持9 min;分流比:30∶1;进样方式:自动进样器(Agilengt Technologies),进样量:0.2 μL;定量方法:内标法,内标物:异丁醇(分析纯)。
2 结果与分析
2.1 原生质体制备条件
本实验选择溶菌酶进行破壁,溶菌酶作用温度37 ℃,设置不同的破壁浓度和时间分别进行镜检,破壁完成后使用细菌计数板对球形原生质体进行计数,绘制曲线如图1。

图1 不同浓度溶菌酶及作用时间对原生质体形成的影响Fig.1 Effect of different concentrations and action time of lysozyme on protoplasm formation
根据图1可以看出:当溶菌酶浓度小于4 mg·mL-1时,溶菌酶不能完全达到破壁目的,酶与细胞作用较弱;当溶菌酶浓度为5 mg·mL-1时,菌体短时间内被大量溶解,不易控制,而在4 mg·mL-1的溶菌酶作用下,出现一条比较清晰的原生质体形成曲线,形成的原生质体状态相对稳定,因此选择4 mg·mL-1的最终溶菌酶使用浓度,4 mg·mL-1的溶菌酶在作用30 min时,原生质体浓度达到最高值,因此决定使用4 mg·mL-1的溶菌酶作用30 min作为最佳原生质体制备条件。
2.2 各种再生培养基对原生质体再生的影响
本实验选用了RCM、PDA、SRA固体培养基作为原生质体再生培养基。将经过PEG诱导融合的菌体接种于RCM、PDA、SRA固体培养基上,37 ℃培养2~3 d后对平板上的菌落进行计数,培养基种类与菌落形成数如图2。

图2 不同培养基的对菌体再生的影响Fig.2 Effects of different media on protoplasm regeneration of Clostridium acetobutylicum
从图2中可以看出,在SRA固体培养基上形成的菌落数最多,SRA固体培养基上原生质体再生能力最强,因此选择SRA培养基作为再生培养基进行实验。
2.3 菌株HY1214原生质体致死时间的确定
将丙酮丁醇梭菌使用4 mg·mL-1的溶菌酶作用30 min。在55℃ 10、20、30、40、50、60 min时分别将菌体涂布于SRA平板上,观察HY1214的致死时间,由图3可见,发现HY1214菌株原生质体在30 min时致死率达到100%,决定选用55 ℃作用30 min作为HY1214菌株原生质体的致死条件。

图3 不同致死时间对菌株HY1214原生质体的致死效果Fig.3 Lethal effect of different lethal time on protoplasm regeneration of Clostridium acetobutylicum
2.4 PEG分子量与作用时间的确定
分别使用50% PEG1000、PEG4000、PEG6000对原生质体诱导融合不同时间[10],观察3种分子量的PEG作用不同时间的诱导融合能力,结果如图4。从图4可以看出,长时间的作用并不能增加PEG的诱导融合作用,反而对原生质体融合与再生不利,PEG4000在与原生质体混合物作用5 min时,产生的菌落最多,故采用PEG4000融合5 min作为最终的诱导融合条件。

图4 不同PEG分子量和作用时间对原生质体融合影响Fig.4 Effect of different PEG molecular weights and time of action on protoplasmic fusion
2.5 Ca2+、Mg2+对原生质体再生能力的影响
一定浓度的Ca2+、Mg2+有利于细胞膜的稳定性,对原生质体再生有一定作用[10]。在本实验中,待原生质体经PEG4000 5 min诱导融合以及PPM洗净后分别加入不同量的Ca2+、Mg2+溶液,菌体再生数量与Ca2+、Mg2+的溶液加入量关系如图5所示。从图5中可以看出,当Ca2+、Mg2+溶液的终浓度分别为50、75 mmol·L-1时,使得原生质体膜稳定程度最高,对原生质体再生能力促进最为明显,因此选用Ca2+、Mg2+溶液的终浓度分别为50、75 mmol·L-1作为最终的离子添加浓度以促进原生质体再生,达到实验目的。

图5 加入不同浓度的Ca2+、Mg2+对原生质体再生的影响Fig.5 Effect of different concentrations of Ca2+ and Mg2+ on protoplasmic regeneration
2.6 高产及新性状筛选
融合再生后,将平板上的菌落洗下后接种于含有13、15、18 g·L-1丁醇的液体SRA培养基中进行耐丁醇梯度筛选,通过气相色谱检测丙酮、乙醇、正丁醇产量,计算正丁醇以及总溶剂的转化率,结果见表1。

表1 融合菌株的产量、转化率与原始菌株的比较Table1 Yield and conversion rate of fusion strains compared to the original strain
在相同的总糖与发酵条件下,215I、215II与原始菌株相比,丁醇与总溶剂的转化率都有提高,因此产量也就得到了提高。215I的表现最突出:丁醇转化率比原始菌株提高了34.7%,产丁醇能力比原始菌株提高了32.2%,达到了本实验设计的最初目的,最直接地体现了产量的提高。
在不断的筛选过程中,发现有的融合子产生了新的物质,而且经过鲜菌体以及芽孢发酵重复验证后,产物非常稳定。结果见图6,通过对色谱图的比较可以看出融合菌株P1147在丙酮峰之后有一新物质峰出现,首先怀疑是酸类物质的相应峰,但在调节pH为中性后,该峰并没有消失,有可能该物质是醇或酯类物质,还需进一步进行鉴定。
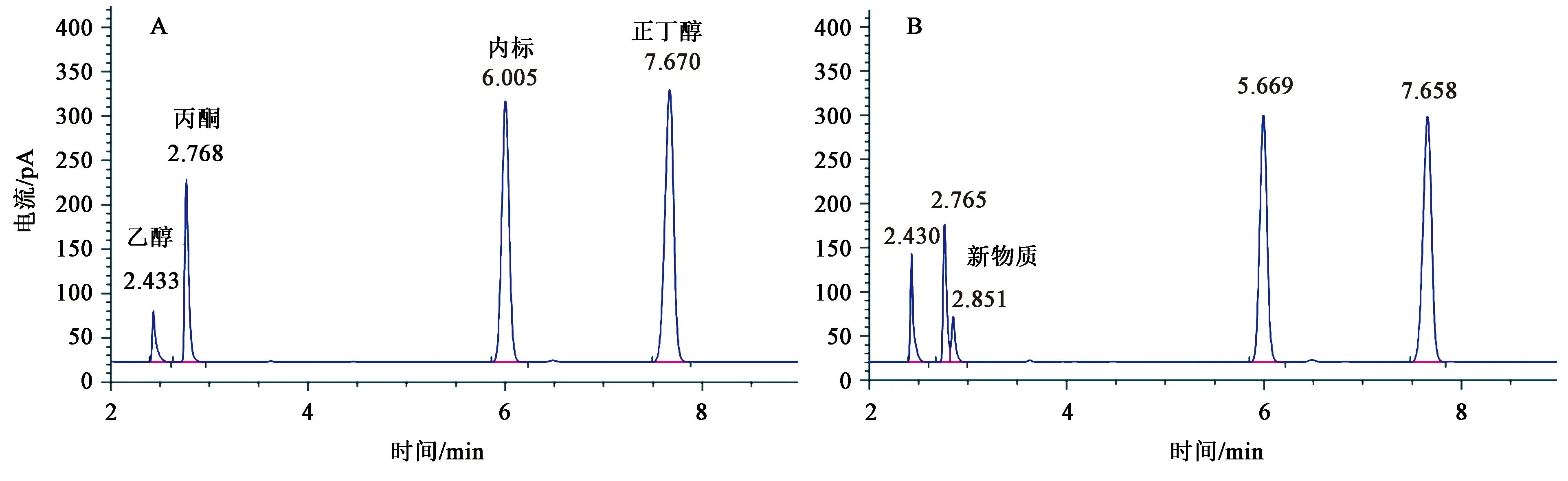
A:HY1710发酵液色谱图;B:P1247 菌株发酵液色谱图
3 讨论
实验中发现梭菌原生质体的制备与再生效果跟菌龄有很大关系,采用对数期中期[2]的菌体能够达到一个比较理想的破壁率和再生率,考虑可能的原因是:在对数期早期,细菌相对脆弱,对不利因素抵抗能力差,一旦失去细胞壁,较难再生;对数期中期菌体较为均一,细胞壁处于最薄的状态,易于与体外溶菌酶作用,同时细胞膜具有韧性,再生能力强;对数期后期原生质体包膜会形成褶皱,再生率下降。经过反复实验与OD值比较,发现实验菌株在16 h时最易破壁,原生质体形成率与再生率最高。
有文献报道,一定浓度的Ca2+、Mg2+作用于细胞膜表面的酶类与多糖类物质,能够影响渗透压,防止渗透压的剧烈变化对原生质体膜[18]的不良影响,对原生质体的再生起到了一定的稳定作用。因此在本实验中加入了Ca2+、Mg2+,所得结果与文献报道相同[10],加入后再生平板上的菌落数明显增多,Ca2+、Mg2+对原生质体的再生起到了促进作用。
本实验通过对实验室保存的生产菌株原生质体形成、融合以及再生条件的摸索,得到了适合该生产菌株的原生质体融合条件。目前国内关于丙酸丁醇梭菌原生质体融合方面的研究尚无报道,国外关于这方面的报道也只涉及到原生质体的破壁与再生,本文首次对丙酸丁醇梭菌原生质体融合进行了全面阐述。对于原生质融合关键是出发菌株的选择,本实验所选菌株HY1710的特点为发酵速度较快,但是耐丁醇能力较差,因此产溶剂能力较差,碳氮源的转化利用效率较低;菌株HY1214发酵速度缓慢,但细胞膜耐丁醇能力较强,相应的产溶剂能力较强,实验根据原生质体融合具有“去除不良性状,产生优异性状”的特点,通过两株菌之间的原生质体融合,使得耐丁醇能力得到显著提高,提高了碳源的转化效率,达到优化菌株的目的。通过原生质体融合实验,在300多株融合子中筛选得到了215I、215II、P1147 3株能够稳定遗传的、性状较为优秀的菌株,使得正丁醇的转化效率得到了提升,节约了发酵成本,提高了产量,但是我们也要看到:本实验中的正丁醇22%左右的转化率对比丙酮丁醇梭菌29%的极限丁醇转化率还有一段距离,需要进一步研究。另外通过融合育种产生的新产物,需要进一步的鉴定,就目前的数据分析,可能是一种醇类或酯类物质。
本实验是在单轮原生质体融合的基础之上所得到的结果,而目前比较流行的原生质体融合技术是通过多轮原生质体融合(即基因组重排技术)可以有效的提高突变几率[19],因此在今后的实验中要进行多轮原生质体融合,并结合原生质体的自身优点,进行基因操作与改造,必定会得到更加高产的生产菌株。
猜你喜欢
齐鲁工业大学学报(2021年1期)2021-12-30
新农业(2020年21期)2020-11-19
中国酿造(2020年10期)2020-11-04
中国酿造(2020年4期)2020-05-15
江苏农业科学(2018年21期)2018-12-05
特产研究(2015年1期)2015-04-12
食品工业科技(2014年23期)2014-03-11
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
中南林业科技大学学报(2012年8期)2012-12-29
中国兽药杂志(2012年4期)2012-11-06
