
miR-181通过调控TGFBR1表达抑制颅内动脉瘤中平滑肌细胞的表型转换*
2021-12-16 02:26王如科孙源源李克芬李一帆于国渊
国际检验医学杂志 2021年23期
王如科,孙源源,李克芬,李一帆,于国渊△
1.河北省邯郸市中心医院神经外科,河北邯郸 056001 ;2 河北省邯郸市第一医院影像科,河北邯郸 056001
颅内动脉瘤(IA)是脑动脉主要分叉的病理性扩张,是一种常见的脑血管疾病,急性病死率约为50%[1-2]。3%~5%的成年人患有未破裂的IA[3]。IA发生和破裂的机制仍不清楚,IA的病因可能与血流对血管壁的剪切力应激、炎症、氧化应激和凋亡等因素有关。炎症级联反应触发后大量炎症因子和氧化因子被释放,会导致许多病理过程,包括中膜弹力层中断、血管平滑肌细胞(VSMC)表型转换和细胞外基质重塑等[4-5]。当血管壁退化时,颅内动脉变得太薄弱,无法抵抗血液动态力,脑动脉管壁发生囊状扩张。VSMC是血管壁中主要的细胞类型,VSMC有两种表型:收缩型和合成型[6]。在炎症和氧化应激等病理刺激下,收缩型VSMC可以转换为合成型[7]。VSMC表型转换在几种心血管疾病中发挥重要作用[8]。microRNAs (miRNAs)是一种短链单链非编码RNA,广泛参与许多转录后调控过程,在调控各种细胞过程中发挥重要作用[9]。miRNAs除参与肿瘤的发生和发展外,也参与IA的发生和发展。有研究报道,miR-370-3p通过靶向IA中的KDR/AKT信号通路抑制平滑肌细胞增殖[10]。有研究发现,VSMC中miR-181的异常表达可能与动脉粥样硬化和脑血管疾病的发生相关[11-12]。例如,在VSMC中血管紧张素Ⅱ(Ang Ⅱ)降低了miR-181的表达水平,过表达miR-181抑制了Ang Ⅱ诱导的合成表型基因骨桥蛋白(OPN)的表达和平滑肌细胞与胶原的黏附[13],提示miR-181可能通过调节OPN介导的VSMC功能,参与动脉粥样硬化的病理生理过程。本研究旨在探究IA中miR-181在平滑肌细胞表型转换中的作用,采用TargetScan预测其靶蛋白并验证其作用机制。
1 资料与方法
1.1标本收集 收集邯郸市中心医院神经外科2016年11月至2019年5月确诊为IA并进行手术的患者80例,并收集同期健康体检中心年龄、性别匹配无IA家族史的健康对照者80例作为研究对象。手术中取出的IA组织在液氮中快速冷冻。收集接受开颅治疗的创伤患者的正常颞浅动脉。本研究经邯郸市中心医院伦理委员会批准,所有研究对象均签署知情同意书。
1.2主要材料 Trizol试剂购自美国Thermo公司,TaqMan反转录试剂盒购自日本Takara公司,实时荧光定量反转录聚合酶链反应(qRT-PCR)试剂盒购自美国 Biotuim 公司。miR-181模拟物(miR-181 mimics)由上海吉凯基因公司合成,人脑VSMC购自美国ScienCell公司。Lipofectamine 2000转染试剂购自美国Thermo公司。α-平滑肌肌动蛋白(SMA)、平滑肌22α蛋白(SM22α)、β-actin抗体均购自武汉三鹰生物技术有限公司。OPN、基质金属蛋白酶(MMP)-2、MMP-9抗体均购自美国Abcam公司。转化生长因子β受体1(TGFBR1)的3′非翻译区(3′-UTR)荧光素酶报告基因载体[TGFBR1野生型(wt)及突变型(mut)]由上海吉凯基因公司构建,双荧光素酶活性检测试剂盒购自上海翊圣生物科技有限公司。DMEM培养基、胎牛血清(FBS)均购自美国Gibco公司。辣根过氧化物酶标记的山羊抗兔二抗购自武汉三鹰生物技术有限公司。PVDF膜购自美国Mllipore公司。
1.3实时荧光定量聚合酶链反应(qPCR) IA患者和健康对照者均抽取空腹肘静脉血约5 mL,室温静置30 min,3 000 r/min 离心5 min,吸取上清液置于-80 ℃冰箱保存。标本收集完毕后,向200 μL上清液中加入1 mL Trizol裂解液吹打混匀后室温裂解5 min。每管中加入200 μL氯仿,室温放置15 min,12 000 r/min,4 ℃离心15 min。吸取上清液加入等体积异丙醇,室温放置10 min,12 000 r/min,4 ℃离心10 min,弃掉上清液,用1 mL 75%乙醇(DEPC水配制)重悬沉淀,8 000 r/min,4 ℃离心5 min,晾干沉淀。用去离子水溶解RNA。
将RNA进行反转录得到模板单链cDNA,然后采用 qPCR试剂盒对cDNA 进行qPCR。按照试剂盒说明书配制20 μL体系,收集荧光信号。每个样品做3个复孔。Ct值为荧光信号达到设定阈值的循环数。用2-ΔΔCt法计算miR-181的相对表达水平。miR-181引物F:GCGAACATTCATTGCTGTCG;R:GCAC TGGATACGACACCCAC。用U6作为内参,U6引物F:GCGCGTCGTGAAGCGTTCT;R:CACTGGA TACGACAAATATG。
1.4细胞培养 人脑VSMC快速复苏后,800 r/min离心5 min,弃掉冻存液,采用含10% FBS的DMEM高糖培养基重悬细胞,放置在37 ℃、5% CO2培养箱中培养,每隔2 d更换一次细胞培养基。当细胞生长至80%~90%融合时用0.25%胰酶进行消化,磷酸盐缓冲液(PBS)冲洗3次后,细胞计数,以每孔1×105个接种到6孔培养板中,放置在37 ℃、5% CO2培养箱中继续培养。
1.5细胞转染 人脑VSMC呈对数生长期时,采用0.25%胰酶消化细胞,PBS冲洗3次,采用含10% FBS的DMEM高糖培养基重悬制成单个细胞悬液。细胞计数后以每孔1×105个铺至6孔板中,分为miR-181 mimics和NC mimics,细胞贴壁18~24 h后,按照Lipofectamine 2000说明书将miR-181 mimics和NC mimics分别与无血清培养基混匀后加入各组待转染的细胞中,置于培养箱中培养48 h后进行后续实验。
1.6Western bolt 收集转染完成的细胞,每孔中加入100 μL蛋白裂解液,在冰上裂解20 min。加入适量上样缓冲液,以100 ℃水浴煮沸5 min使蛋白变性。按照50 μg总蛋白加入上样孔进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳,转膜封闭完成后,在4 ℃冰箱孵育一抗过夜,第2天洗膜后孵育相应的二抗,洗膜后进行增强化学发光法。条带用Image J进行灰度值半定量计算,采用β-actin作为内参,分析各目的蛋白的相对表达水平。
1.7荧光素酶报告基因实验 取对数生长期的人脑VSMC,接种于24孔板中(每孔细胞2×104个),培养至细胞达70%~80%融合时将TGFBR1的3′-UTR荧光素酶报告基因载体及NC mimics导入细胞,分为4组,每组设置3个复孔:miR-181 mimics+TGFBR1-wt共转染组;miR-181 mimics+TGFBR1-mut共转染组;NC mimics+TGFBR1-wt共转染组;NC mimics+TGFBR1-mut共转染组。培养24 h后弃去培养液,用预冷的PBS冲洗2次后,加入100 μL 1×被动裂解液,孵育15 min后每孔吸取20 μL上清液加入96孔发光检测板中,放入发光检测仪中读数,数据用GrahPad Prism 8进行作图与分析。
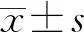
2 结 果
2.1IA患者和健康对照者血清miR-181表达水平比较 采用qPCR检测IA患者和健康对照者血清miR-181表达水平,结果显示,IA患者血清miR-181表达水平与健康对照者比较明显降低,差异有统计学意义(P<0.05),见图1。
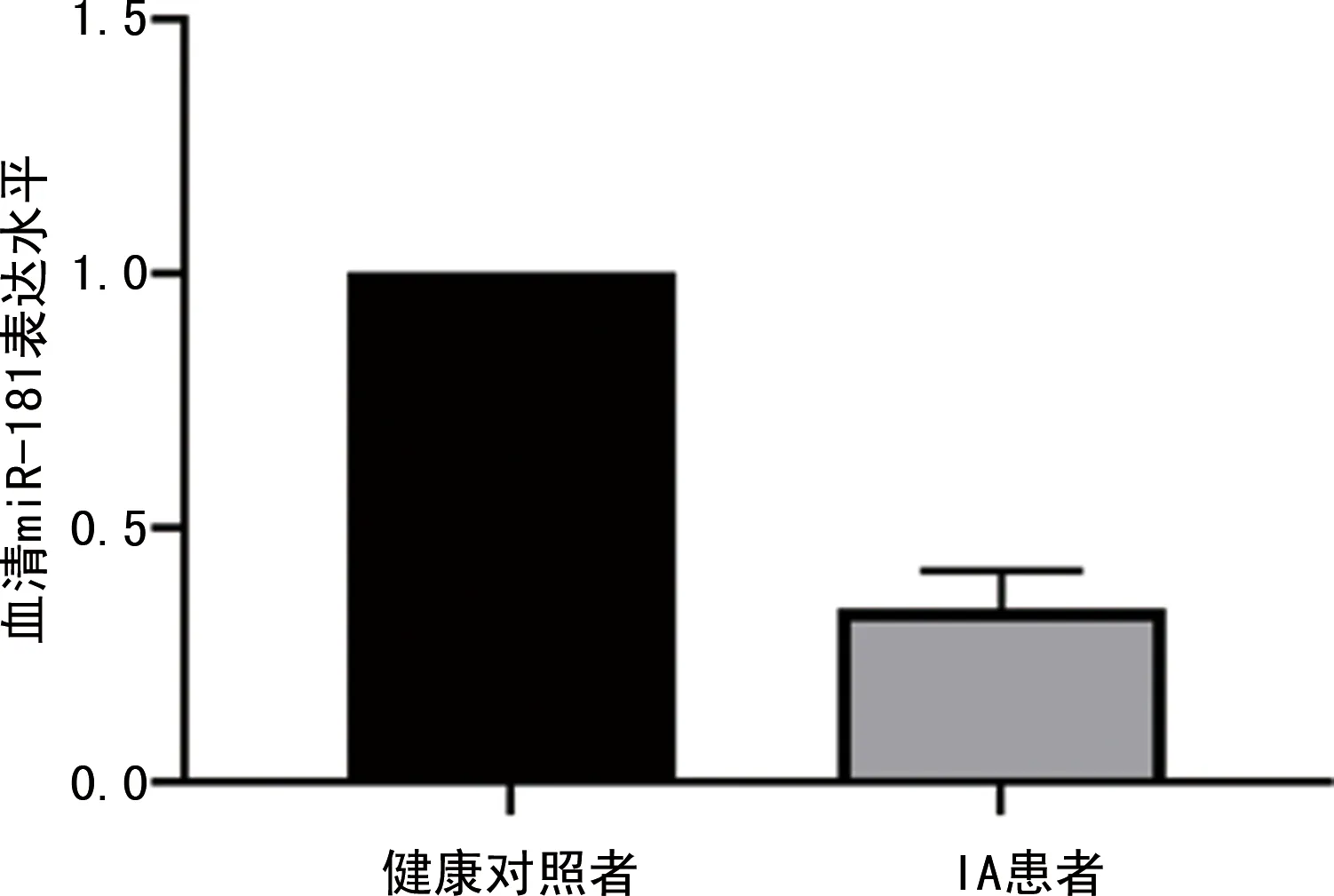
图1 IA患者和健康对照者血清miR-181表达水平
2.2miR-181抑制VSMC表型转换 人脑VSMC细胞转染miR-181 mimics和miR-181抑制剂(miR-181 inhibitor)后采用Western bolt检测VSMC收缩表型基因SMA和SM22α及合成表型基因OPN的表达水平,以及其分泌的MMP-2和MMP-9的表达水平,结果显示,转染miR-181 mimics升高了SMA和SM22α的表达水平,降低了OPN的表达水平,同时MMP-2、MMP-9的表达水平也降低。转染miR-181 inhibitor降低了SMA和SM22α的表达水平,上调了OPN的表达水平,同时MMP-2、MMP-9的表达水平也上调。见图2。
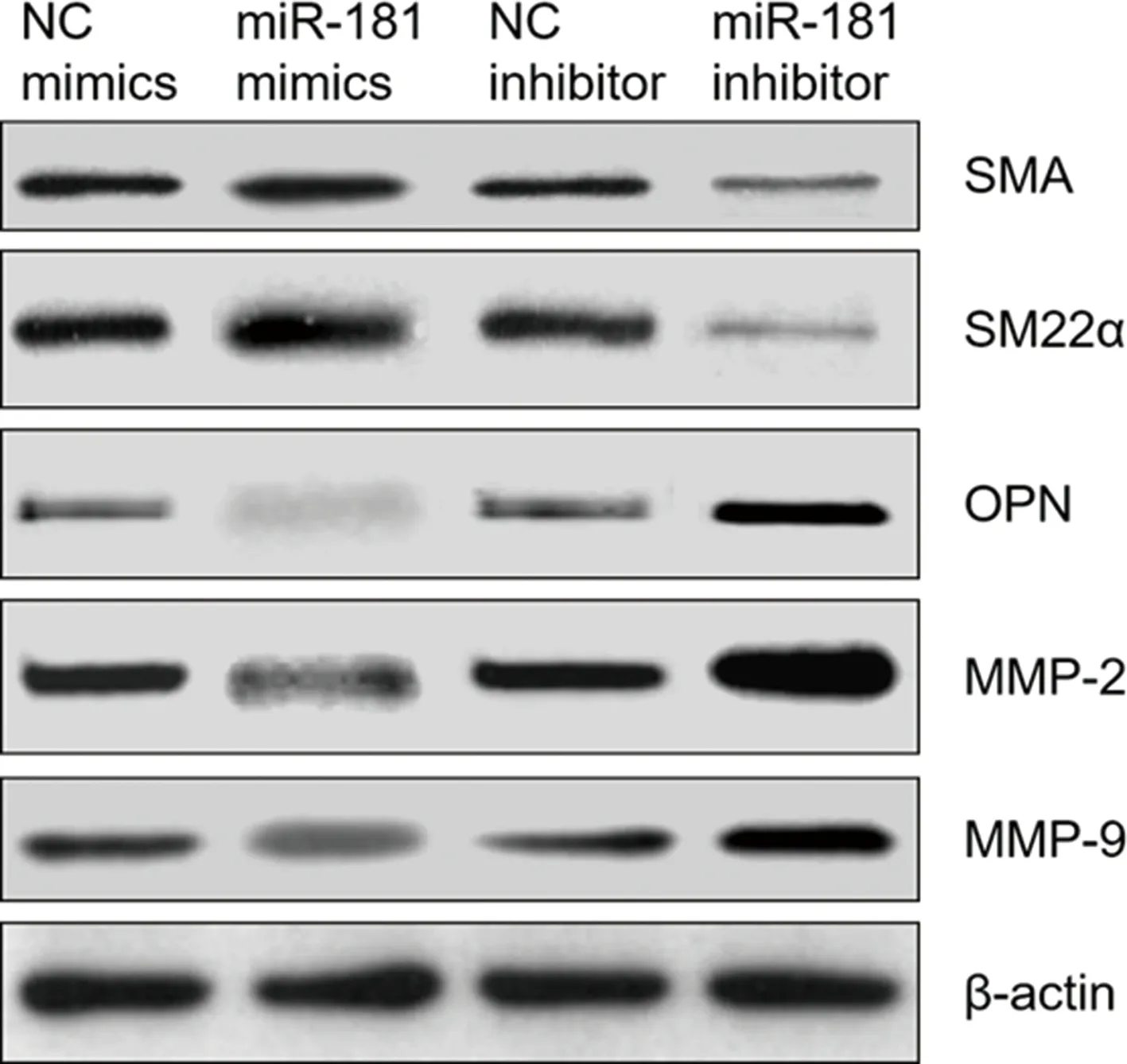
图2 miR-181对脑动脉VSMC表型转换的影响
2.3TGFBR1是miR-181的直接靶点 TargetScan软件预测结果显示,miR-181与TGFBR1的3′-UTR存在互补结合位点(图3A),miR-181可能通过靶向调控TGFBR1的表达水平而发挥作用。双荧光素酶报告基因检测结果显示,TGFBR1-wt与miR-181 mimics共转染组荧光素酶活性明显低于阴性对照组,差异有统计学意义(P<0.05);而TGFBR1-mut各组间荧光素酶活性比较,差异无统计学意义(P>0.05),图3B。
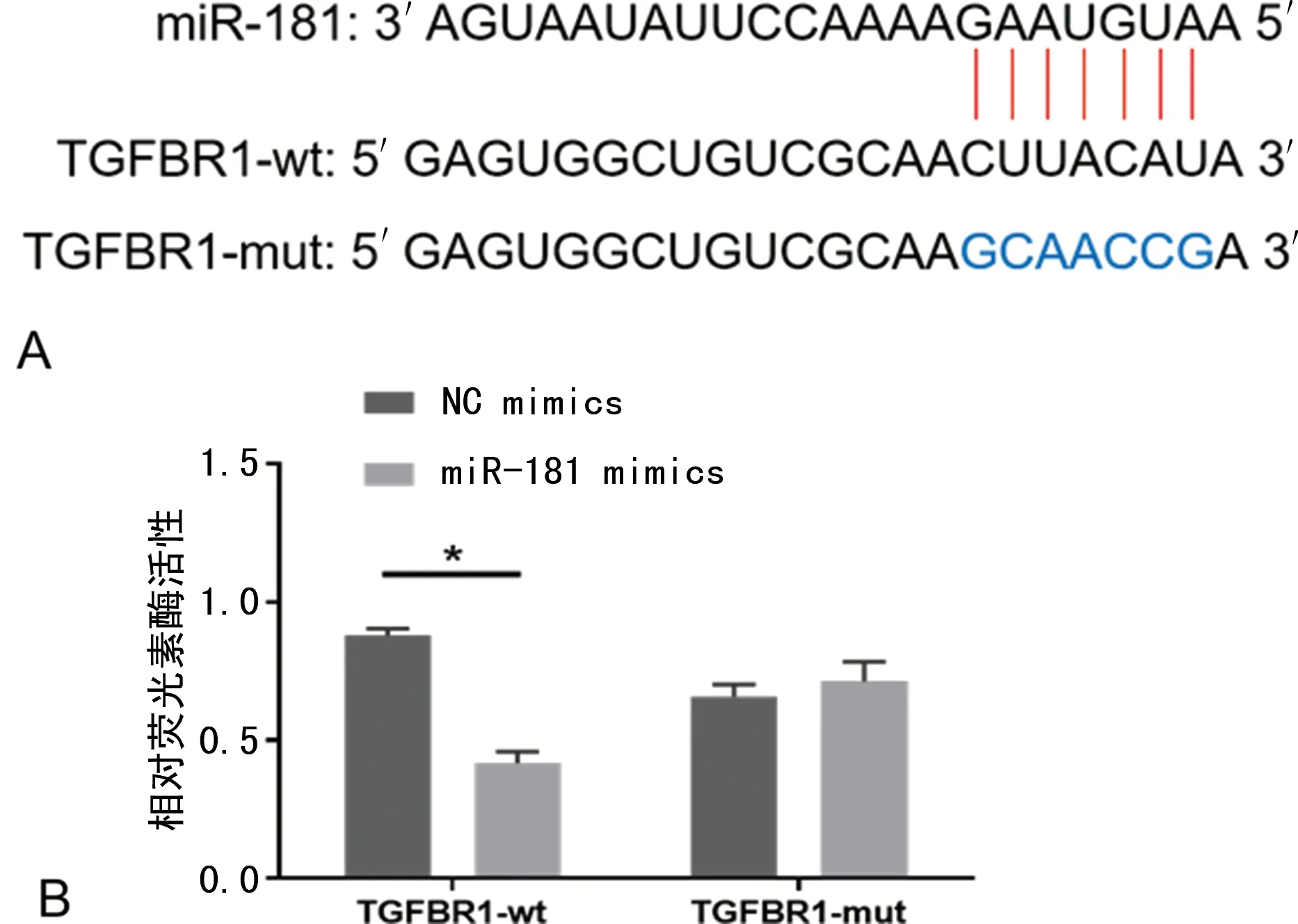
注:A为TargetScan软件预测TGFBR1是miR-181的靶基因;B为双荧光素酶报告基因检测结果,与NC mimics比较,*P<0.05。
2.4IA组织中TGFBR1蛋白表达水平 采用Western blot检测IA患者手术切除的IA组织中TGFBR1蛋白表达水平,并采用接受开颅治疗的创伤患者的正常颞浅动脉作为对照。结果显示,与对照者比较,IA患者IA组织标本中TGFBR1蛋白表达水平升高,见图4。

图4 IA组织中TGFBR1表达水平
2.5miR-181靶向调控TGFBR1的蛋白表达水平 Western blot分析表明,转染miR-181 mimics降低了TGFBR1的蛋白表达水平,转染miR-181 inhibitor升高TGFBR1的蛋白表达水平。见图5。
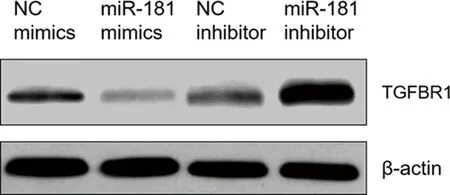
图5 miR-181靶向调控TGFBR1的蛋白表达水平
3 讨 论
虽然IA的发病机制尚不清楚,但氧化应激、炎症、血流动力学失调和细胞外基质降解均有助于IA的形成和发展。VSMC表型转换在IA、动脉粥样硬化和动脉瘤等疾病中发挥重要作用。有研究表明,miR-181通过调控VSMC表型转换参与动脉粥样硬化的发生和发展[11]。有研究表明,VSMC表型调控与IA的形成、生长和破裂相关[14-15]。VSMC由收缩型转换为合成型,合成型VSMC刺激促炎过程、细胞外基质重构和细胞凋亡,最终促进IA的形成和破裂。本研究结果显示,IA患者血清miR-181表达水平低于健康对照者。为了探究IA中miR-181对VSMC表型转换的作用,采用miR-181 mimics和miR-181 inhibitor转染人脑VSMC检测收缩表型的标记SMA、SM22α及 OPN的表达水平,结果表明,VSMC过表达miR-181基因细胞SMA、SM22α表达水平升高,OPN表达水平降低。同时抑制miR-181表达水平后SMA、SM22α表达水平降低,OPN表达水平升高。VSMC合成表型与收缩基因的表达降低有关,虽然VSMC失去了收缩能力,但有助于招募促炎细胞和血管壁细胞外基质重构。本研究结果提示,抑制miR-181基因VSMC MMP-2、MMP-9表达水平均升高。
miRNA可通过调控基因转录后水平抑制mRNA的降解和翻译。miRNA与其靶基因转录体序列互补,通过切断靶基因的mRNA分子和(或)抑制靶基因的翻译来发挥作用。几个miRNA可以共同调控某一基因的表达[16]。TargetScan软件预测结果表明,TGFBR1为miR-181的靶向调控基因。TGFBR1是一种特异性TGF-β受体,介导TGF-β的生理和病理功能,已有文献证明TGFBR1是动脉粥样硬化过程的主要成分[17]。TGFBR1在TGF-β介导的心血管疾病涉及的几乎所有细胞类型中均起作用,包括内皮细胞、VSMC、肌成纤维细胞和巨噬细胞[18]。本研究采用Western blot检测了IA患者手术切除的IA组织中TGFBR1的蛋白表达水平,并用接受开颅治疗的创伤患者的正常颞浅动脉作为对照,结果显示,与对照者比较,IA组织中TGFBR1的蛋白表达水平升高。因此猜测,在IA中miR-181通过调控TGFBR1的蛋白表达水平调控VSMC表型转换。
为了研究miR-181与TGFBR1的靶向调控关系,本研究采用TargetScan软件预测及荧光素酶报告基因检测验证上述猜测。结果表明,TGFBR1是miR-181直接调控的靶基因。通过Western blot检测发现,过表达miR-181后TGFBR1的蛋白表达水平降低;而抑制miR-181后TGFBR1的蛋白表达水平升高。
综上所述,IA患者血清中miR-181表达水平下降,过表达miR-181抑制VSMC收缩表型向合成型表型转换。miR-181直接靶点是TGFBR1,IA组织中TGFBR1的蛋白表达水平升高。因此,IA中miR-181可能通过调控TGFBR1的蛋白表达水平抑制VSMC的表型转换,这为治疗IA及控制蛛网膜下腔出血提供了新的靶点。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
实用手外科杂志(2022年2期)2022-08-31
中国现代医生(2022年21期)2022-08-22
中国临床解剖学杂志(2022年4期)2022-07-30
中国临床解剖学杂志(2022年2期)2022-04-22
农村科学实验(2022年2期)2022-03-12
中国临床解剖学杂志(2022年1期)2022-02-14
三农资讯半月报(2020年2期)2020-03-09
江苏农业科学(2019年23期)2019-03-03
小学生学习指导(中年级)(2018年12期)2018-11-29
