
流行性出血病的全球流行及分布
2022-11-07 02:10罗倩敏韦丽韩佃刚叶玲玲杨云庆董俊张冲李静尹尚莲李瑶瑶何昌霖董仙兰李丹丹唐晓萍艾军
中国动物检疫 2022年11期
罗倩敏,韦丽,韩佃刚,叶玲玲,杨云庆,董俊,张冲,李静,尹尚莲,李瑶瑶,何昌霖,董仙兰,李丹丹,唐晓萍,艾军
(1.昆明海关技术中心,云南昆明 650200;2.昆明海关技术中心大理分中心,云南大理 671000;3.海口海关技术中心,海南海口 570311;4.昆明市技术合同认定登记站,云南昆明 650021)
流行性出血病(epizootic haemorrhagic disease,EHD)是由流行性出血病病毒(epizootic haemorrhagic disease virus,EHDV)引起的,经库蠓叮咬传播的非接触性反刍动物传染病[1]。EHD主要影响野生和家养反刍动物。白尾鹿(Odocoileus virginianus)感染EHDV 后可发生严重的出血性疾病,临床表现为发热、乏力、食欲不振、流涎过多、面部水肿以及结膜和口腔黏膜充血、冠状炎和口炎,发病率和死亡率很高,是影响白尾鹿健康的重要疾病之一;骡鹿、黑尾鹿和叉角羚羊等感染EHDV后发生的症状较轻,死亡率低;牛、绵羊感染后,产奶量减少,发生流产或产畸形胎,从而引起巨大生产损失[2]。世界动物卫生组织(WOAH)将EHD 列入了须通报动物疫病名录[1]。本文对EHD的病原学特征及流行特征进行综述,以期为EHD防控提供借鉴。
1 病原学特征及传播媒介
1.1 病原学特征
EHDV 属于呼肠孤病毒科环状病毒属,在遗传学和形态学上都与蓝舌病病毒(BTV)相似。EHDV 基因组包含10 段线性双链(ds)RNA(Seg-1—Seg-10),编码7 种结构蛋白(VP1—VP7)和4 种非结构蛋白(NS1、NS2 和NS3/NS3a)。结构蛋白VP1、VP3、VP4、VP6 和VP7 形成病毒核心颗粒,而VP2 和VP5 形成病毒粒子外壳层。VP2 和VP7 分别是决定血清型和血清群的两种主要蛋白质[3]。Seg-3将EHDV 毒株分为两种地域型(topotype),中国、日本和澳大利亚毒株属东方型(eastern),美洲和非洲毒株属西方型(western)[4]。不同地域型同一种毒株在致病性方面也存在差异。例如,在日本出现的EHDV-2 毒株对牛有致病性,而在北美洲流行的同源毒株则没有[5]。
EHDV 在高温下极不稳定,50 ℃ 3 h,60 ℃15 min 即可失活;在pH6.0~8.0 下敏感,pH6.8~9.5稳定,pH4.0 以下以及pH10 以上迅速失活;对乙醚和氯仿等脂质溶剂具有抗性,容易被β-丙内酯、2%戊二醛、酸、2%氢氧化钠、2%~3%次氯酸钠、碘伏和酚类化合物灭活;试验[5]发现:在不同温度条件下,EHDV在血液和组织样本中有不同反应,-70、4 和20 ℃条件下EHDV 表现稳定,可长期保存活力,-20 ℃时表现不稳定;对紫外线和γ 辐射线具有抵抗力。
EHDV 最初在淋巴管内皮细胞和引流部位感染的淋巴结中复制。然后,通过血流传播到其他部位复制,如淋巴结和脾脏。体外研究[5]表明,EHDV 复制可能依赖于单核细胞。目前,全球各地分离到的EHDV 血清型至少有9 种,包括EHDV-1、EHDV-2、EHDV-4—EHDV-10(表1)。
1.2 传播媒介
EHD 是虫媒传染病,多种因素会影响虫媒摄入、维持和传播病原体的能力。雌性库蠓C.sonorensis可能会在吸食感染鹿的血液时摄入EHDV,但并非所有蠓都会传播病毒(表2 列出的为已经确认能够传播EHDV 的库蠓属)。虫媒携带并传播EHDV 的可能性受许多遗传和环境因素影响,如摄入的病原体数量、昆虫的免疫反应和媒介-病原体基因型相互作用等[12]。
2 全球流行及分布
2.1 美洲
据报告,在已知的血清型中,美洲仅存在EHDV-1、2 和6 型。C.sonorensis是北美洲唯一被确认的EHDV 传播媒介。EHDV 于1955 年首次在美国新泽西州的白尾鹿中被分离到,同年在美国密歇根州,1956 年在美国南达科他州,1962 年在加拿大艾伯塔省,1967 年在美国华盛顿州,1971 年在美国西南部的白尾鹿中均分离到该病毒[13]。

表1 全球EHDV 血清型分离情况[6-11]
2006 年在美国印第安纳州和伊利诺伊州发现的EHDV-6,是澳大利亚EHDV-6(CSIRO 753)和北美EHDV-2(艾伯塔省)组合的一种新型重组毒株[14]。2008 年在美国堪萨斯州和德克萨斯州,2009 年在美国密苏里州和密歇根州,2010 年在法属瓜德罗普,均分离到EHDV-6,表明该病毒已经越冬[5,15]。2012 年美国15 个州发生了大规模的多血清型EHDV 感染疫情,导致牛临床发病和死亡;随后,2013 年又暴发了小范围疫情,病死率高达26%[6]。
2013 年,在进口到特立尼达加勒比岛的无症状牛中发现EHDV-6。该病毒的10 个片段中,有8个来自于美国EHDV-6 印第安纳毒株(该毒株是含有澳大利亚EHDV-6 片段的重组毒株),另外的第4 段来自澳大利亚的EHDV-2 毒株,第8 段与美国新泽西EHDV-1 毒株核苷酸同源性最高(90.4%)。这种重组病毒可能来自美洲,在那里它从一种局部循环的EHDV 毒株中获得了第8 片段。经分析发现,该毒株在与美国当地流行的EHDV-2 毒株重组之前,就已在加勒比海地区传播,并可能从加勒比海或南美洲向北传播至北美洲[16]。
Allen等[17]对加拿大安大略省(2016—2019年)家畜血清进行EHDV 抗体检测,检出15 头牛(9.0%)呈EHDV-2 血清阳性。在法属圭亚那,分离出EHDV-1、EHDV-6;在厄瓜多尔,295 头牛血液样本中检测到EHDV 抗体,阳性率为81.3%,血清型为EHDV-1 和EHDV-6[18]。在墨西哥的白尾鹿中,EHDV 抗体阳性率为72%;在美国德克萨斯州的白尾鹿中,EHDV 抗体阳性率为90%[19]。在美国佛罗里达州,使用RT-qPCR 方法对2016—2020 年死亡的养殖白尾鹿进行EHDV 血清型鉴定,发现EHDV-1 占19%,EHDV-2 占55%,EHDV-6占27%(表3)。
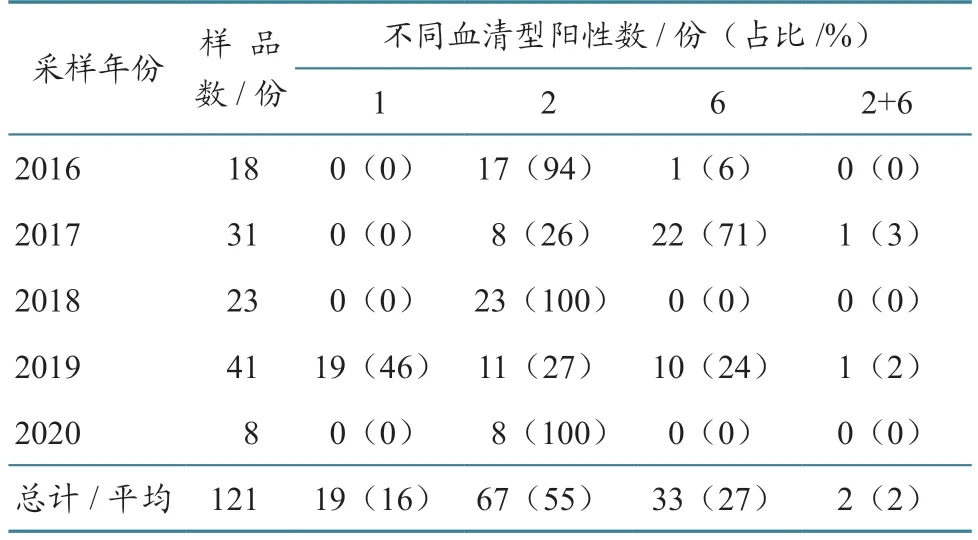
表3 2016—2020 年对佛罗里达州鹿场死亡白尾鹿EHDV 的血清型鉴定[20]
据WOAH-WAHIS 数据库统计,2010 年以来美洲有4 个国家或地区报告发生EHD,分别是法属瓜德罗普、法属圭亚那、加拿大、巴西(表4)。
根据WOAH 报告和相关文献报道,美洲EHDV 主要流行于北美洲南部和南美洲北部的加勒比地区,包括美国、加拿大、法属圭亚那、墨西哥等国家或地区。这与它们发达的牛羊养殖业以及湿润的大陆性气候有关。美洲是白尾鹿原产地,其中北美洲国家不仅有野生白尾鹿,还有提供鹿角等传统装饰物的小型农场人工饲养白尾鹿。这是美洲地区EHDV 流行并向外传播的重要因素之一。
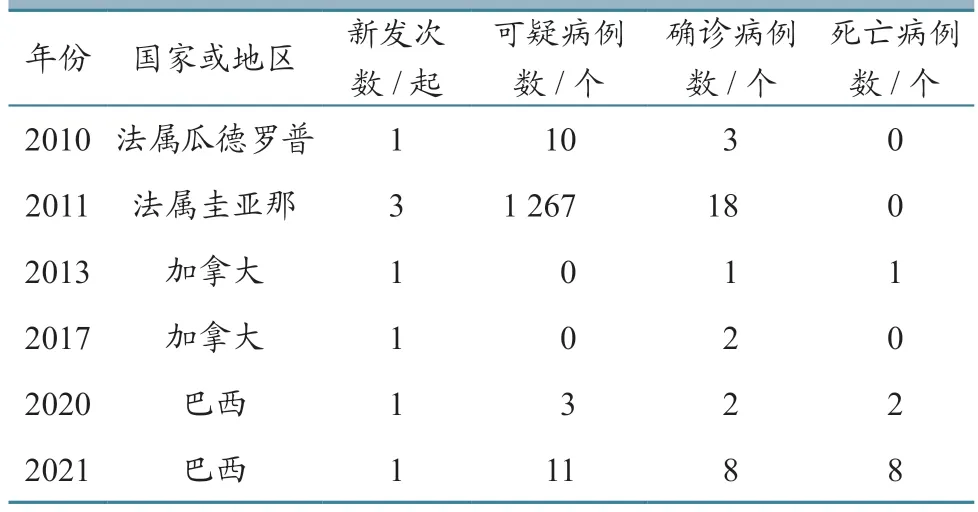
表4 EHDV 在美洲的流行情况统计
2.2 亚洲
1959 年在日本分离的EHDV-2(茨城)是首次证明对牛有致病性的EHDV 毒株[14]。当时该病毒导致4 000 头牛感染死亡,并在受影响地区的39 000 头牛身上造成高发病率[21]。2006 年从以色列牛中分离出来的EHDV-7 被证实起源于非洲[22]。该病毒由西、北向南传播至位于以色列、巴勒斯坦、约旦交界处的死海,并沿约旦裂谷向北传播[5]。2015 年、2016 年在以色列分别发现了EHDV-6 和EHDV-1,其中EHDV-1 与尼日利亚毒株亲缘关系密切[23]。EHDV-2(茨城)、EHDV-6和EHDV-7 分别在日本、土耳其和以色列引起牛病,而相同血清型在澳大利亚则不能引发牛病[5,24]。有报道[15]称,在巴林分离到的EHDV-6(318 株)与“西方型”病毒同源性较高,而与“东方型”病毒同源性较低。
和东华等[25]对我国云南省边境地区牛进行EHDV 血清学调查,发现8 个市(县)的抗体阳性率为72.7%~98.75%,检出6 种血清型,分别为EHDV-1、2、5、6、7、8。寇美玲等[26]对云南省进行EHDV 流行调查,发现11 个县(市)的抗体阳性率为49.3%~91.3%,检出6 种血清型,分别为EHDV-2、5、6、7、8、10。我国分离到的EHDV-1、6、10 与日本毒株亲缘关系较近[4,10,27]。研究人员对土耳其南部的绵羊和牛进行了血清调查,发现EHDV-1 阳性率分别为0.4%和0.9%,EHDV-2阳性率分别为6.5%和4.5%[19]。
据WOAH-WAHIS 数据库统计,2006 年以来,亚洲通报EHDV 感染的国家共有3 个,分别是土耳其(乌沙克、穆拉、马尼萨、伊兹密尔、代尼兹利、布尔杜尔、艾丁、安塔利亚)、以色列(耶路撒冷区、耶路撒冷分区、塔拉巴、以色列专属经济区、北部区、中央区、海法、南部区、戈兰)和日本(熊本、鹿儿岛)。具体数据见表5。
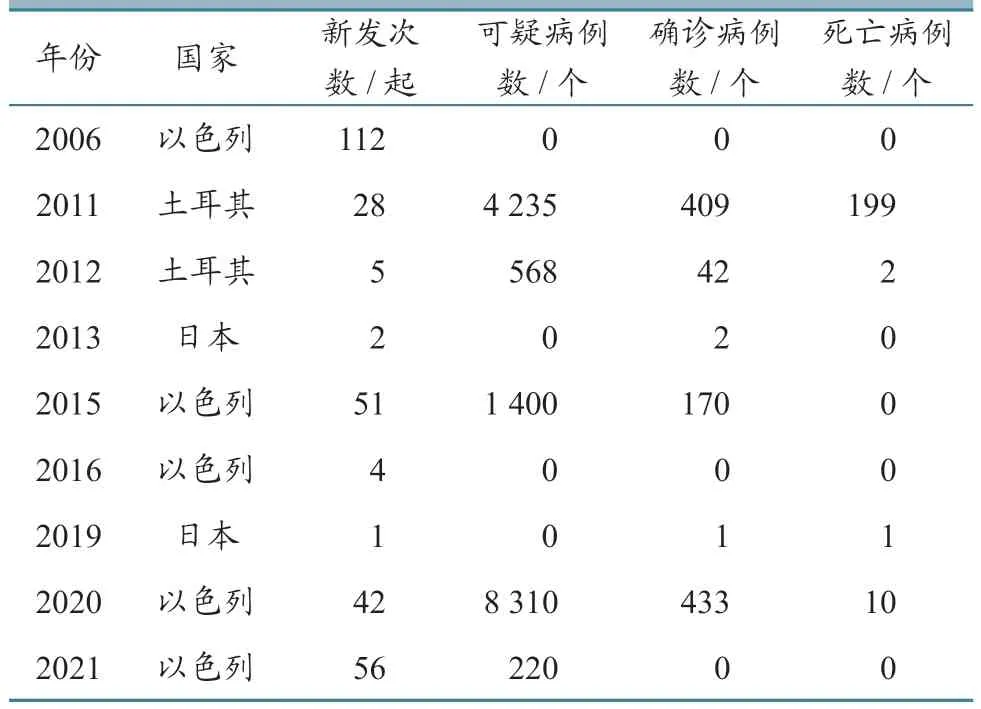
表5 EHDV 在亚洲的流行情况统计
综上,亚洲EHDV 流行主要集中在日本、以色列、土耳其,它们都是亚洲牛羊养殖业发达的国家。疫情也佐证了EHDV 能够大规模侵袭牛羊,造成疫病发生和死亡的事实。我国在云南省边境地区,从牛体内分离到6 个EHDV 血清型,牛抗体阳性率也较高,表明我国边境地区已遭受境外EDHV 侵袭。
2.3 非洲
2004 年,摩洛哥报告了由EHDV-6(318 株)引起的疫情。2006 年,同一血清型导致该国再次暴发疫情,发病率为18.0%,病死率为2.2%;同年EHDV-6 在阿尔及利亚和突尼斯引起疫情[5]。2006 年对在突尼斯传播的EHDV-6 毒株S2、S3、S6、S7和S10核苷酸序列比较发现,这5 个片段与在同一地理区域(非洲、阿拉伯半岛和印度洋)分离的EHDV 毒株具有共同的起源[28]。Sailleau等[15]对留尼汪岛分离到的EHDV-6 毒株基因组第2 段序列进行分析,发现2003 年与2009 年在该岛引起流行的毒株有高度同源性。这些数据表明,相同的毒株已经在岛上传播了7 年。经序列比较发现,留尼汪岛流行的EHDV-6 毒株与2006—2007 年在地中海盆地流行的EHDV-6 毒株密切相关。
肯尼亚小牛中的EHDV 抗体阳性率为64%,留尼汪岛牛羊中的EHDV 抗体阳性率为38%[29]。2016 年在埃及牛群中检出了EHDV 阳性抗体,血清型为EHDV-1[30]。Cosseddu等[31]研究发现,毛里塔尼亚骆驼中的EHDV 阳性率高达73%。Mahmoud等[14]调查了利比亚的EHDV 血清流行情况,检测了2015 年采集的855 份血液样本,发现EHDV 总体血清流行率约为4%,其中小型反刍动物血清阳性率显著高于牛,绵羊的血清流行率高于其他物种,由此可知绵羊可能在EHDV 流行中发挥重要作用。Mejri等[32]在突尼斯不同地区采集了牛和单峰驼血清样本进行EHDV 抗体筛查,发现2012—2013 年采集的牛样品中EHDV 抗体阳性率最高(表6)[32]。
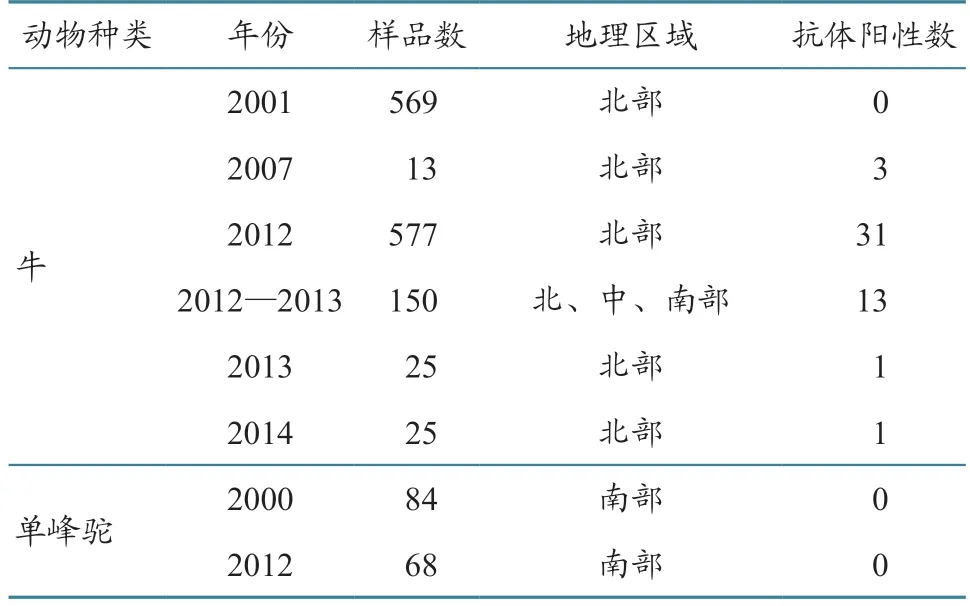
表6 突尼斯2007—2014 年EHDV 流行情况[32] 单位:份
依据WOAH-WAHIS 数据统计,非洲共有5个国家或地区通报发生EHDV 感染,分别是突尼斯(宰格万、托泽尔、希里亚奈、西迪布兹德、斯法克斯、斯贝特拉、尼拜尔、莫娜斯提尔、马努巴、马赫迪那、卡赛林、凯鲁万、坚杜巴、比泽太、本阿鲁斯、巴贾、阿里亚纳)、摩洛哥(东部区、中南区、中北区)、阿尔及利亚(麦迪亚、拉格瓦特、杰勒法)、法属留尼汪、纳米比亚(哈达普)。具体数据见表7。
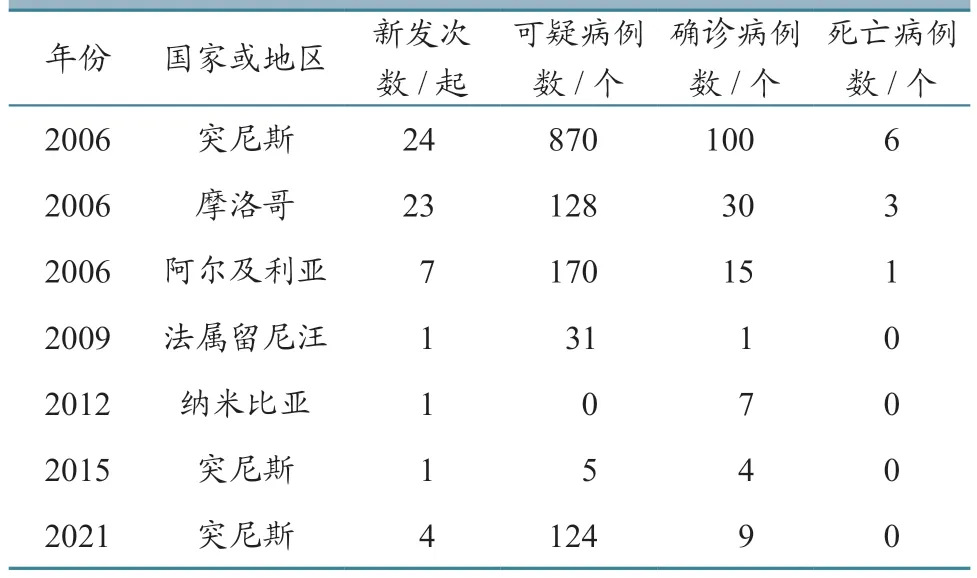
表7 EHDV 在非洲的流行情况统计
综上所述,非洲EHDV 流行集中在北非地区的地中海沿岸国家,主要影响牛、骆驼和小型农场的绵羊。北非地区EHDV 的流行对地中海周边国家和欧洲其他区域形成严重威胁。在印度洋的法属留尼汪,EHDV-6 流行时间很长,它与地中海盆地国家流行毒株具有同源性。
2.4 欧洲和大洋洲
至今,WOAH-WAHIS 数据库中未有欧洲和大洋洲通报发生EHDV 感染的相关数据。但据相关文献记载,澳大利亚目前存在6 种EHDV 血清型,包括EHDV-1、2、5、6、7 和8[16,33]。库蠓C.imicola是较常见的传播EHDV 的媒介物种,其虽然是一种亚非物种,但已在南欧建立了自己的地位,目前广泛分布于葡萄牙,西班牙巴利阿里群岛,法国南部大陆、科西嘉岛,意大利大陆、撒丁岛、西西里岛,希腊大陆,土耳其安纳托利亚以及附近的几个希腊岛屿。C.obsoletus和C.punctatus广泛分布于北欧和中欧的大部分地区。C.schultzei在非洲和中东广泛分布,但在欧洲,只有希腊南部大陆的南阿提卡才有记录[5]。
欧洲虽然没有EHD 病例报告,但在其周边亚非国家却有疫情发生,对欧洲形成传入风险。在地中海盆地以及留尼汪岛流行的EHDV-6 和EHDV-7对牛具有高致病性,表明除EHDV-2 外,其他血清型也可能对家畜致病,并可能产生相当大的经济损失[21]。此外,在地中海盆地,已报告了4 种EHDV 血清型,如土耳其的EHDV-1、EHDV-2 和EHDV-6,阿尔及利亚、摩洛哥和突尼斯的EHDV-6,以色列和约旦的EHDV-7[14]。这些血清型在地中海盆地的出现也显著增加了EHDV 入侵中欧和北欧的风险[20]。
值得注意的是,EHDV 与20 世纪90 年代末流行的BTV 存在惊人的相似性。在此期间,一些最初在阿尔及利亚、突尼斯、土耳其和以色列传播的BTV 血清型能够穿越地中海,通过西部、东部和南部走廊入侵南欧[5]。
2.5 全球分布变化
EHDV 在世界范围内广泛传播,2011 年以前EHDV 影响北美洲和中美洲,南非和中非,以及中东、大洋洲的澳大利亚和东南亚地区[5]。截至目前,EHDV 在原有区域不断向外扩散。据现有数据[5]以及上述情况汇总:北美洲的EHDV 由美国向北扩散到加拿大达艾伯塔省、安大略省;在南美洲,EHDV 已经扩散至厄瓜多尔、法属瓜德罗普、特立尼达和多巴哥、法属圭亚那和巴西,流行的3 种EHDV 血清型至今未变(表8)。在非洲,EHDV已扩散至法属留尼汪、西非毛里塔尼亚以及北非东北部的埃及和利比亚,血清型由原来的3 种增加至4 种。在亚洲,EHDV 已扩散至中国、日本、以色列,血清型由原来的3 种增加至7 种。在大洋洲,EHDV 目前仍局限在澳大利亚,没有扩散现象,血清型也没有增加。全球EHDV 血清型流行变化见表8。
3 全球传播风险因素
3.1 野生宿主种群规模扩大
最初,EHDV 的易感宿主主要是野生鹿,尤其是北美洲的白尾鹿。白尾鹿原产于美洲,曾濒临灭绝,2015 年被列入《世界自然保护联盟濒危物种红色名录》。目前白尾鹿在南美洲数量仍然稀少,但在北美洲如美国、加拿大,由于对白尾鹿的保护,其数量逐渐增多,增加了感染和传播EHDV的风险。
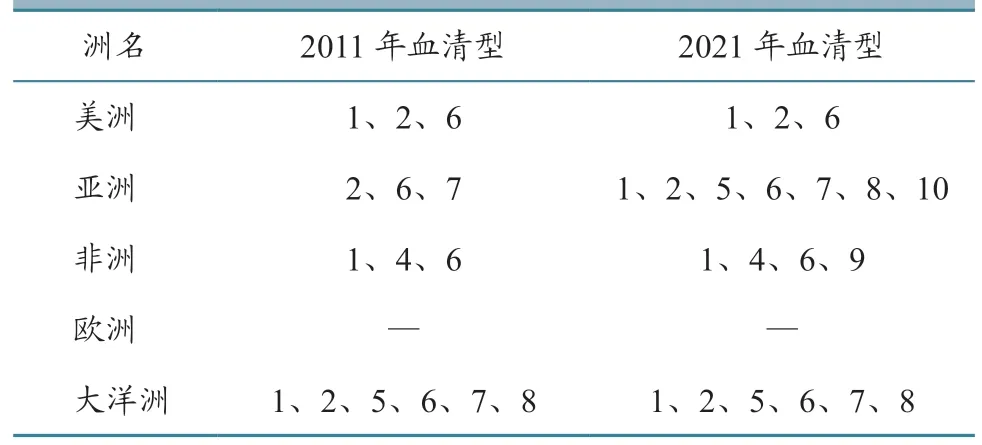
表8 全球EHDV 血清型流行分布变化
3.2 传播方式发生变化
EHDV与BTV在全球的传播区域和传播媒介相似,两者的共同媒介包括C.imicola、C.brevitarsis、C.bolitinos、C.obsoletus、C.gulbenkiani、C.magnus、C.sonorensis和C.stellifer。随着EHDV 在全球的持续流行传播,这两种疫病在一个国家的共同传播正频繁发生[5]。由于虫媒种群扩大,病毒血清型增加,EHDV 对牛羊的致病性也开始提高。牛羊规模化养殖量大、养殖密集,随商业贸易可远距离调运,一旦染疫,EHDV 不仅可在区域内迅速传播,也可跨洲传播,因此养殖牛羊将成为其主要传染源和传播主体。
3.3 病毒基因发生重配
EHDV 属于环状病毒。环状病毒具有巨大的遗传多样性以及相似遗传基因序列,不仅本身可发生变异产生新的病毒,而且其他入侵的基因与病毒本身存在的基因发生重组,促进了新重组子代的出现和传播。
3.4 人为因素促进病毒变异
人类对病毒病的防控行为,如使用减毒活疫苗,过度环境消杀等,可促使病毒变异或出现流行特征变化,以适应新环境生存。这包括:病毒通过媒介蠓传播能力的变化,与地方性病毒重新组合,产生新的遗传变异后代,毒株变异表现新的流行特性,等等。
3.5 气候变化引起传播媒介活跃、范围变广
气候变暖导致EHDV 传播媒介地理范围分布变广,EHDV 在原有区域不断向外扩散,使原来没有节肢动物传播病毒的地区也出现了EHDV,从而导致EHD 全球分布加速。
4 全球控制措施
人类行为导致的动物迁徙和全球贸易流通的加速,使EHDV 迅速形成了全球流行传播趋势。WOAH-WAHIS 数据库收集汇总了2017—2021 年全球对EHD 的15 项控制措施:宰前宰后检 疫(ante and post-mortem inspections)、疫 病报告(disease notification)、常规监测(general surveillance)、监测(monitoring)、移动控制(movement control)、筛检(Screening)、野生动物宿主控制(control of wildlife reservoirs)、病媒控制(control of vectors)、病媒生物监测(vector surveillance)、目的性监测(targeted surveillance)、区域区划(zoning)、边境防控(precautions at the borders)、清群和无害化处理(selective killing and disposal)、扑杀(stamping out)、宰杀(slaughter)。不同国家或地区实施的措施略有不同(图1),其中日本实施的措施最多(8项),多项措施的综合利用使日本长期未发生过大规模疫情;其次澳大利亚为7 项,自1977—1992年分离出6 种血清型以来,至今的30 年内,血清型没有发生任何变化,有效阻断了新血清型EHDV侵入。
5 启示

图1 2017—2021 年EHDV 在全球国家/地区层面的控制措施
近年来,EHDV 在全球流行传播,各种EHDV 血清型及其载体的地理分布范围不断扩大,感染的严重程度和频率逐渐增加,给反刍动物养殖带来了新挑战。目前我国边境地区已发现牛羊EHDV 感染,为有效防范EHDV 传入,建议我国:(1)强化EHDV野生宿主动物活动区域管控监测,防止野生动物与家养动物接触;(2)加强低洼灌木天然草场改造,削弱传播媒介生存环境;(3)加强边境地区的EHD 监测,严厉打击动物走私、逃检行为,降低病毒传入风险;(4)研发EHDV检测技术,提高检测水平,做好EHDV 灭活疫苗研发储备。
猜你喜欢
首都食品与医药(2022年21期)2022-12-06
动物医学进展(2022年9期)2022-11-26
检验医学与临床(2022年17期)2022-09-14
健康护理(2022年3期)2022-05-26
中国典型病例大全(2022年11期)2022-05-13
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
医药前沿(2021年25期)2021-10-16
养猪(2020年6期)2021-01-27
医学概论(2021年19期)2021-01-21
