
福建闽南地区与广西北海地区波纹巴非蛤的形态差异与遗传多样性分析
2023-12-22 12:29周治东张丽艳霍云龙李海平何雪蒨张跃平
应用海洋学学报 2023年4期
周治东,张丽艳,霍云龙,李海平,何雪蒨,张跃平*
(1.福建海洋研究所,福建 厦门 361013; 2.福建省海岛与海岸带管理技术研究重点实验室,福建 厦门 361013;3.福建省海陆界面生态环境联合重点实验室,福建 厦门 361005;4.自然资源部第三海洋研究所,福建 厦门 361005)
波纹巴非蛤(Paratapesundulatas,旧称:Paphiaundulatas)是我国东南沿海地区一种重要的经济贝类,多栖息于低潮区至水深10 m左右的泥砂底质中,俗称“花甲”、“油蛤”等[1]。在我国,波纹巴非蛤自然分布于浙江南部、福建、广东、海南和广西沿岸海域[2-3],其滋味鲜美,营养丰富,纹理清晰,色泽明艳,深受广大消费者喜爱[4-6]。近年来,随着市场需求量的增加,波纹巴非蛤的采捕压力过大,波纹巴非蛤自然资源遭受了毁灭性的破坏[7]。因此,我国已在福建、广西等东南沿海地区大力发展波纹巴非蛤养殖产业[8-9]。然而,近年来福建闽南沿海波纹巴非蛤的浅海增养殖大量采用广西外来苗种,种群结构受人为活动的干扰严重,这种非自然的交流导致不同种群间进行基因交流,致使本地波纹巴非蛤的种质资源遭受破坏,遗传多样性保护迫在眉睫[10]。
迄今为止,国内有关波纹巴非蛤形态学[7,10]、分子遗传学[10]的研究较少,而有关微卫星分子标记的研究多集中在微卫星引物开发方面[11]。本研究对采集于福建闽南地区及广西北海地区的波纹巴非蛤种群样品进行形态学和分子遗传学研究,探讨不同群体间的形态差异和遗传分化程度,以期为波纹巴非蛤遗传特性研究、种质资源的保护及养殖繁育提供理论和方法依据。
1 材料与方法
1.1 样品采集
波纹巴非蛤样品于2020年12月至2021年12月采自福建闽南地区(厦门同安湾海域、云霄东山湾海域、惠安大港湾海域)及广西北海(北部湾海域),以下简称厦门群体、云霄群体、惠安群体、北海群体,每个群体样品数量不少于30枚(图1)。随机选取外壳完好的波纹巴非蛤个体进行形态测量,测量完毕后每个群体随机挑选24枚取其斧足及闭壳肌用95%的酒精固定,以备后续分子遗传学研究。

图1 4个群体波纹巴非蛤样品外壳形态Fig. 1 Shell shape of four Paratapes undulatas populations
1.2 形态学测量方法
采用精度为0.01 mm的游标卡尺对波纹巴非蛤的形态特征参数进行测量:分别获得壳长(Ls)、壳高(Hs)、壳宽(Ws)、韧带长(Lh)、前腹缘距(Af)、后腹缘距 (Ab)6个形态学参数(图2);用精确至0.001 g的电子天平称取壳质量(Ms),共计847个测量数据。
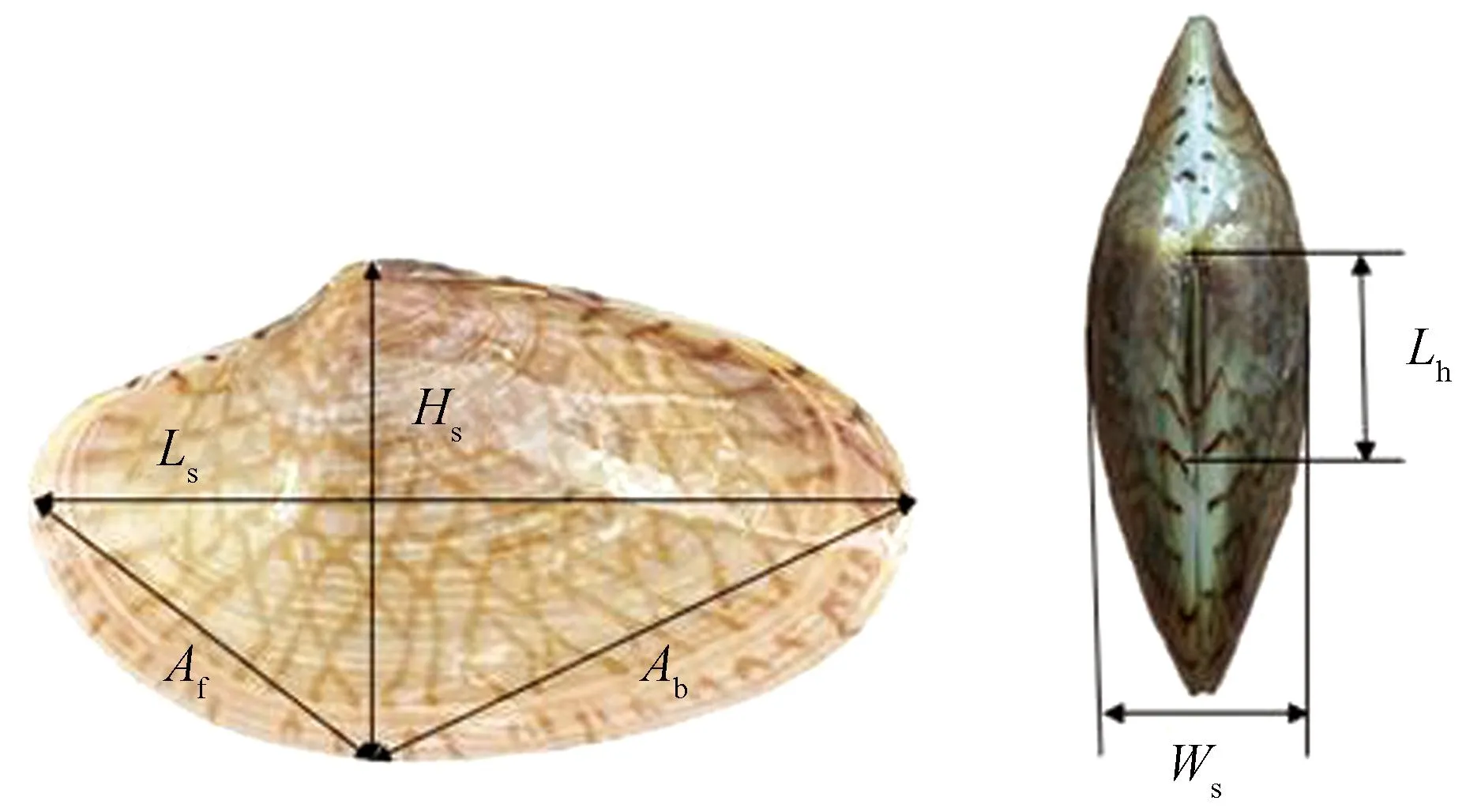
图2 波纹巴非蛤形态学测量参数示意图Fig. 2 Diagram of morphological measurement parameters of Paratapes undulatas
1.3 分子遗传学实验方法
采用天根海洋动物组织基因组DNA提取试剂盒提取其基因组DNA,基因组DNA于4 ℃保存。本研究从已发表的波纹巴非蛤微卫星引物中筛选出10对扩增结果稳定且多态性较好的引物[10-12](表1),用于后续分析。
PCR反应体系为25 μL,包括17.15 μL双蒸水、2.5 μL 10×PCR Buffer、1 μL模板DNA(50 ng/μL)、2 μL dNTPs(2.5 mmol/L)、1 μL反向引物、1 μL荧光标记的M13-R、0.1 μL含有M13-R的正向引物、0.25 μL Taq酶。PCR反应程序:95 ℃变性5 min;95 ℃ 45 s;退火(54~64 ℃)45 s;72 ℃ 45 s;循环30次;72 ℃延伸10 min;4 ℃保存。反应结束后,将PCR产物送青岛派森诺基因科技有限公司进行基因分型[13-16]。
1.4 数据分析
1.4.1 形态学数据计算分析
将各性状测量值处理成5个比例数据(Hs/Ls、Ws/Ls、Lh/Ls、Ms/Ls、Af/Ab)作为校正值对各群体的形态学差异进行计算分析。采用SPSS 13.0软件计算和分析,利用聚类分析、主成分分析和判别分析等方法对4个群体的形态学特征进行比较。其中聚类分析采用欧式距离的最长距离系统聚类法;主成分分析将5个比例性状计算得到两个综合性指标,即两个互不关联的主成分;判别分析对所有样本进行逐个判别,计算判别准确率及综合判别率[17-20]。
1.4.2 分子遗传学数据分析
使用GeneMarker 2.2.0对荧光PCR基因分型数据进行读取,并辅以人工矫正。使用PopGene软件统计每个微卫星位点的等位基因数、多态信息含量指数、观测杂合度以及期望杂合度[21]。使用F-stat 2.9.3软件计算每对群体间遗传分化指数,同时进行1 000次置换检验分析其显著性[22]。使用Population 1.2软件计算群体之间的遗传距离,并使用邻接法(Neighbor-Joining, NJ)基于遗传距离构建系统发育树[23]。使用LEA包对进行Structure分析[24]。通过BOTTLENECK 1.2.02软件采用无限等位基因模型(IAM)、逐步突变模型(SMM)和双相模型(TPM)3种模型检测各群体是否经历过瓶颈效应[25]。若等位基因频率分布为L型,说明所检测的群体近期没有经历过瓶颈效应事件。
2 结果与分析
2.1 形态学研究结果
2.1.1 形态参数测量比较
4个波纹巴非蛤群体的表观形态特征参数测量结果如表2所示。对测量数据进行F检验表明:各群体除壳长外的6个度量形态特征均与壳长呈显著正相关(P<0.01);且群体间各个度量形态特征数据差异显著(P<0.01)。
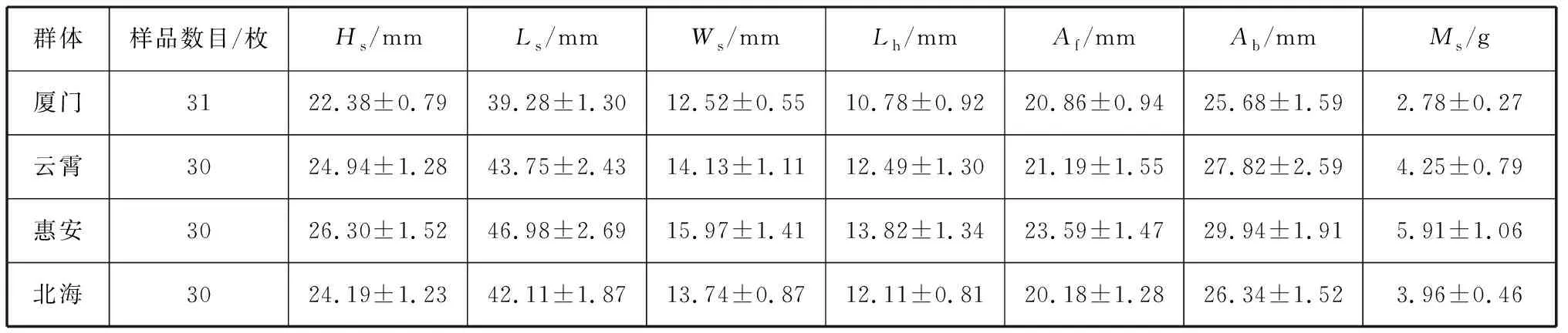
表2 4个波纹巴非蛤群体各参数测量结果Tab. 2 Summary of measurements of various parameters of four Paratapes undulatas populations
2.1.2 聚类分析
对4个群体样本的5个形态特征校正值进行聚类分析结果表明,北海与惠安群体形态特征最为接近,二者首先聚类,后与云霄群体聚类,而厦门群体则自成一支(图3)。
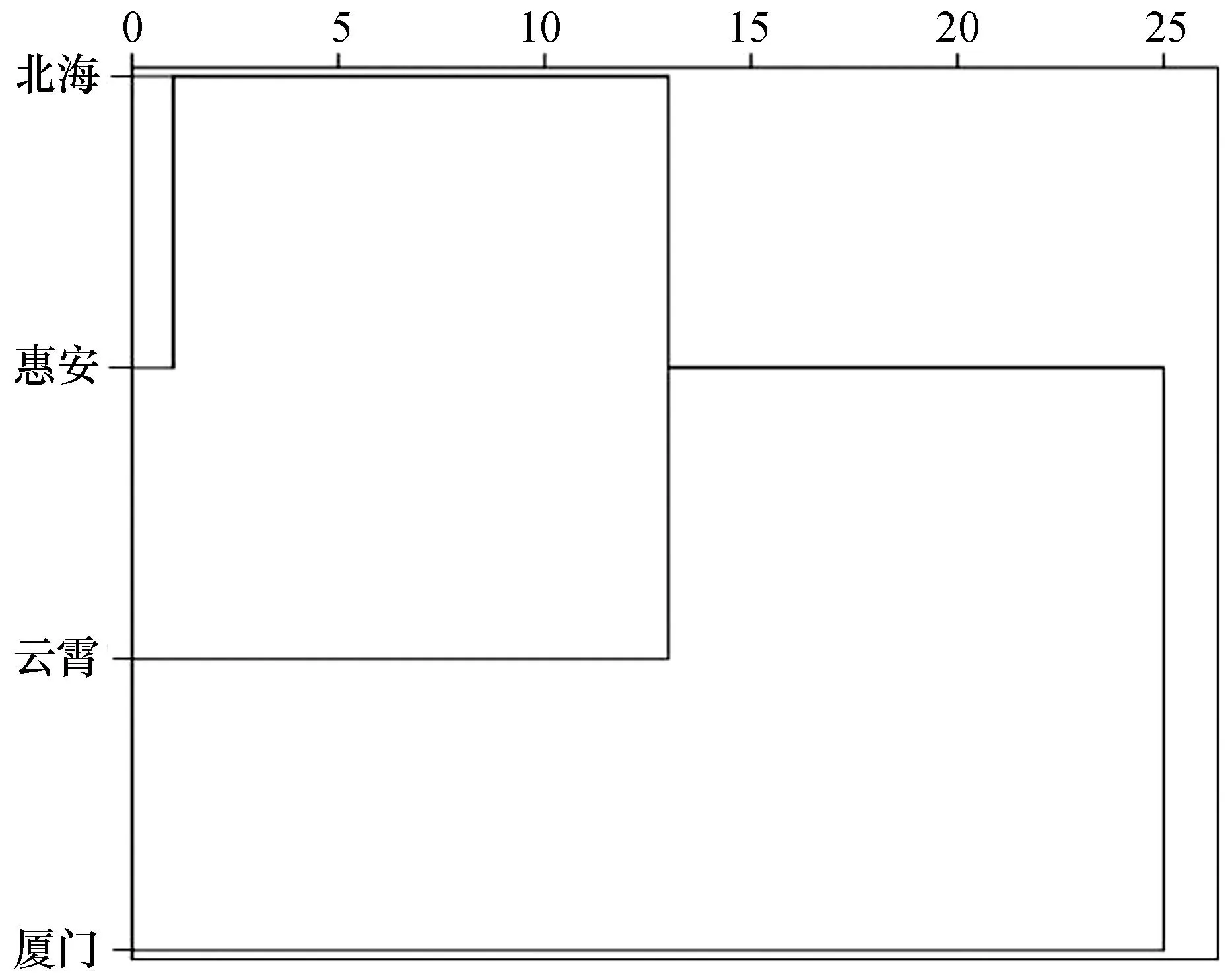
图3 波纹巴非蛤4个群体形态特征的聚类分析Fig. 3 Cluster analysis of four Paratapes undulatas populations
2.1.3 主成分分析
对4个群体样本的5个比例性状进行主成分分析,得到两个主成分PCI和PC2,由表3可知,:PC1的贡献率为36.134%,PC2的为21.443%,二者累积贡献率仅为57.577%。在PC1中,Ms/Ls的贡献最大,第二为Ws/Ls;在PC2中 ,Hs/Ls和Af/Ab贡献最大。这两个主成分的累计贡献率未达到85.0%,说明波纹巴非蛤不同群体间的形态差异难以用几个相互独立的因子来解释。
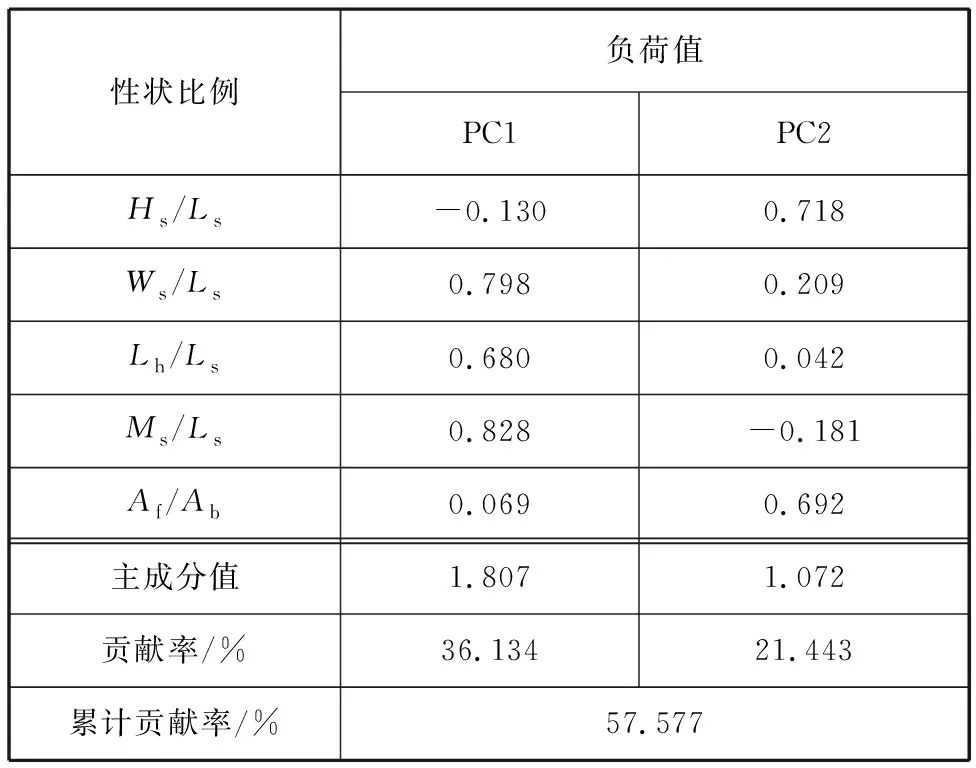
表3 波纹巴非蛤形态特征的主成分的负荷值和贡献率Tab. 3 Contribution and load of principal components on morphological characteristics of Paratapes undulatas
PC1与PC2的散点图如图4所示。由图4可见4个群体中北海与惠安群体有较多的重叠区域,说明二者在形态参数上较为接近,云霄群体与厦门群体均独立分布,较少与其余样品重叠,区分明显。该结果与聚类分析结果基本一致。
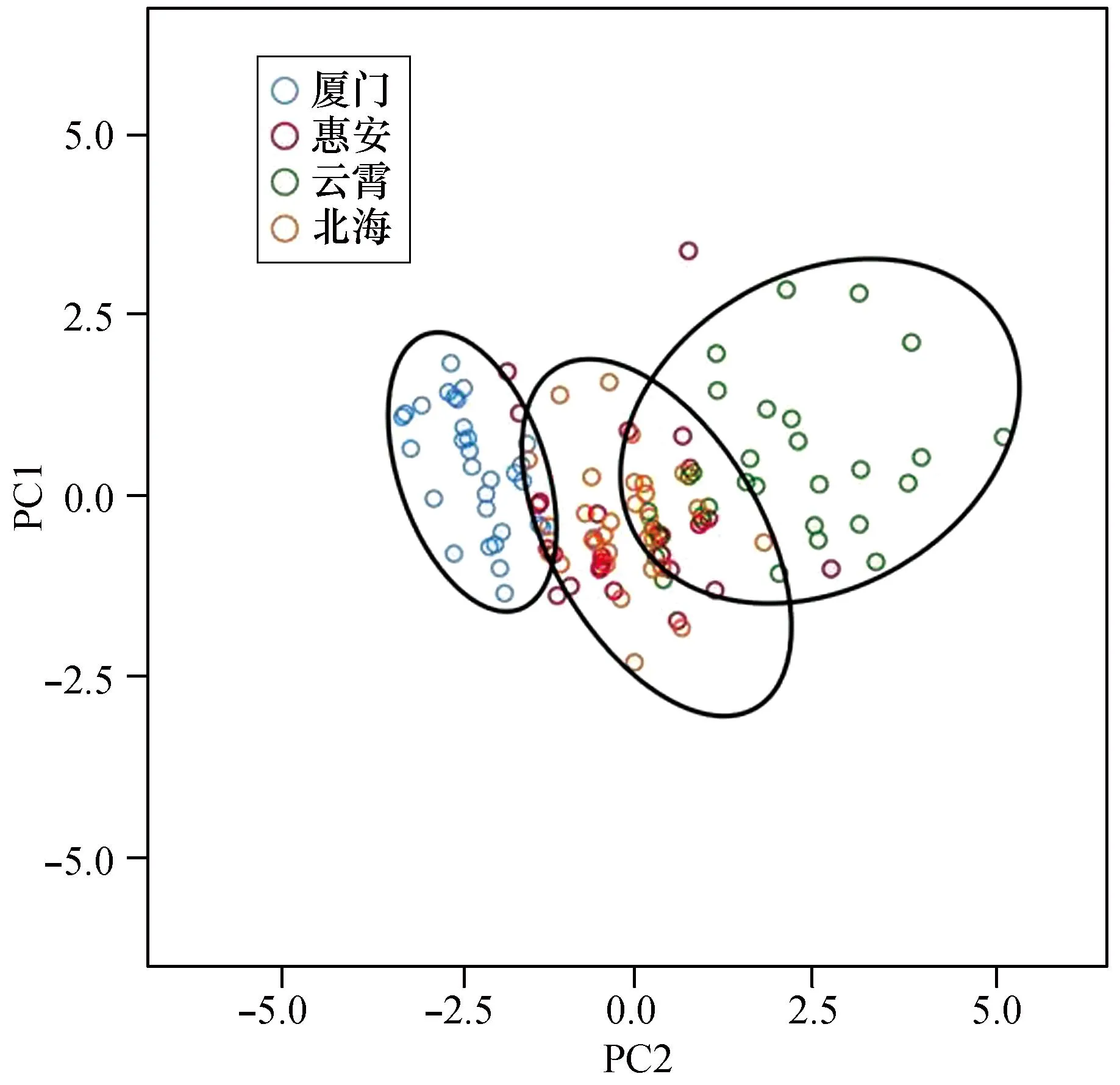
图4 4个波纹巴非蛤群体的第一、二主成分的散布图Fig. 4 Scatter diagram for PC1 and PC2 of four Paratapes undulatas populations
2.1.4 判别分析
利用逐步判别分析法筛选出5个性状比例的特征值,建立4个群体的判别函数,5个变量X1、X2、X3、X4、X5分别代表Hs/Ls、Ws/Ls、Lh/Ls、Ms/Ls与Af/Ab。表3为判别函数的各项系数及常数项。4个群体的判别公式如下:
Y厦门=1 134.881X1+899.281X2+373.775X3
-207.158X4+185.212X5-587.362
(1)
Y云霄=1 141.218X1+871.409X2+389.677X3
-27.383X4+165.181X5-585.651
(2)
Y惠安=1 119.267X1+880.459X2+393.719X3
+144.492X4+167.024X5-597.905
(3)
Y北海=1 148.237X1+882.468X2+393.311X3
-42.030X4+165.234X5-592.944
(4)
判别分析结果如表4所示:判别准确率P1为43.3%~93.5%;判别准确率P2为44.8%~88.0%;4个群体的综合判别率为 66.1%。
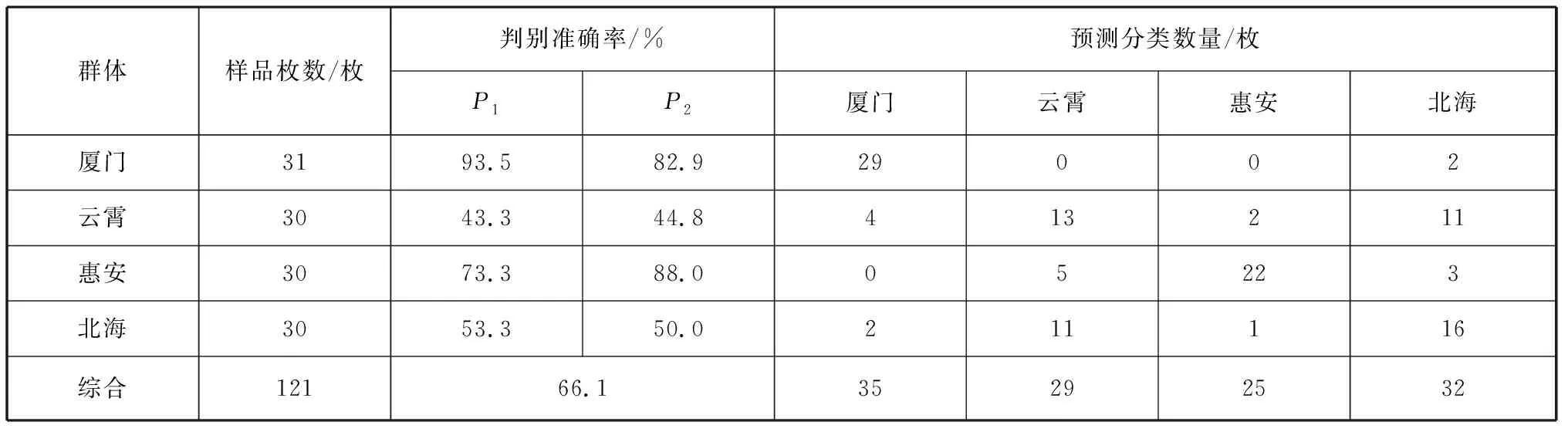
表4 波纹巴非蛤4个群体的判别分析结果Tab. 4 Discriminant results of four populations of Paratapes undulatas
2.2 分子遗传学研究结果
2.2.1 遗传多样性
本研究采用10对微卫星分子引物对4个波纹巴非蛤群体96个个体进行遗传多样性分析。由表5可以看出,4个群体的遗传差异较小,平均等位基因数范围为6.40~6.60对,平均有效等位基因数范围为3.18~3.51对,平均多态信息含量范围为1.29~1.40,平均观测杂合度范围为0.39~0.50,平均期望杂合度范围为0.62~0.70。
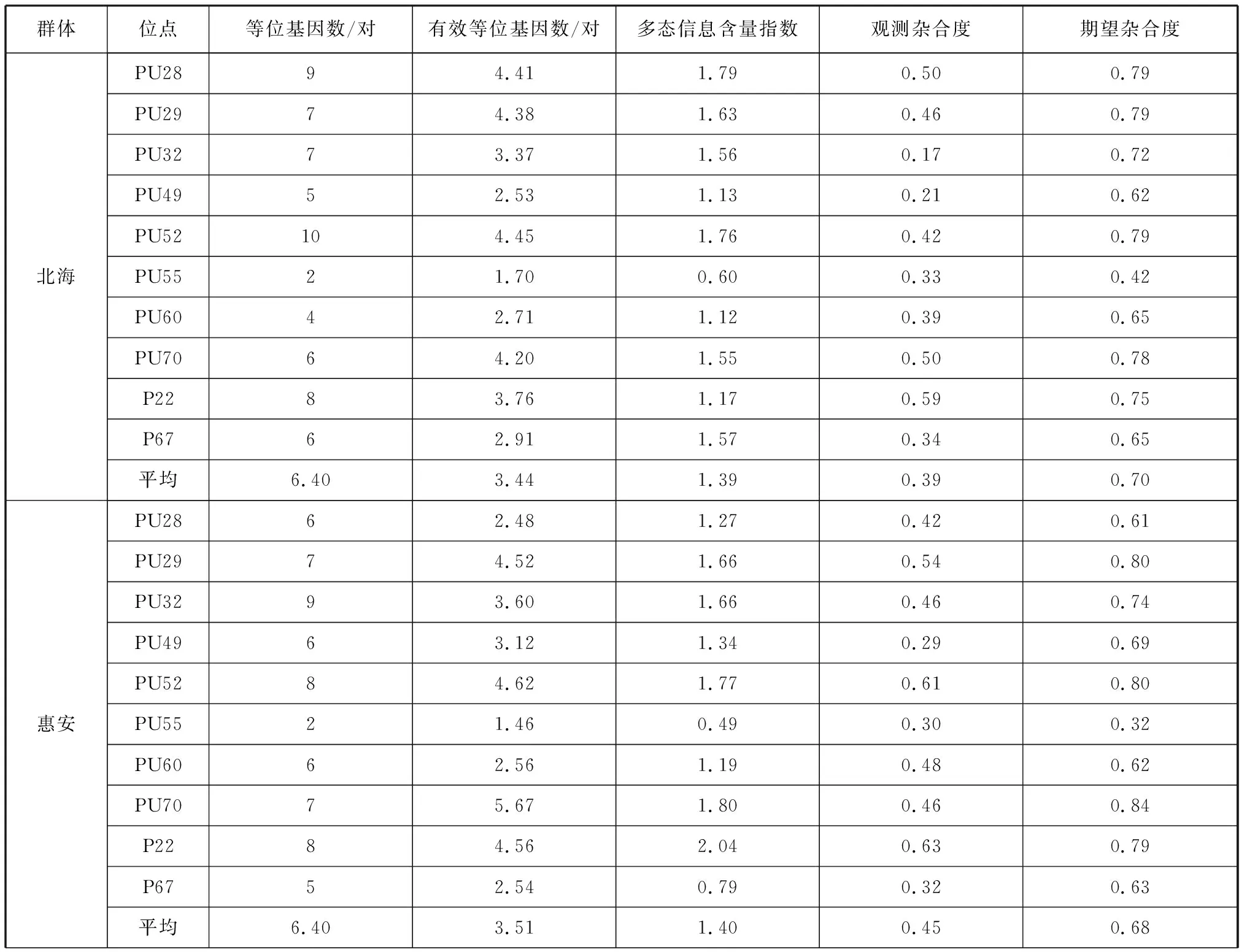
表5 波纹巴非蛤4个群体的遗传多样性参数Tab. 5 Genetic diversity parameters of four Paratapes undulatas populations
综合各遗传多样性指数来看,4个群体中惠安群体的遗传多样性最高(平均期望杂合度为0.68,平均多态信息含量指数为1.40),云霄群体的遗传多样性最低(平均期望杂合度为0.62,平均多态信息含量指数为1.29)。
2.2.2 遗传结构
4个波纹巴非蛤群体间的遗传分化指数范围为0.00~0.03,均呈现较低的状态,表明各群体的遗传分化水平较弱。群体间遗传距离范围为0.08~0.11,云霄群体与其余3个群体间的遗传距离均为最低(0.08),厦门群体与北海群体间的遗传距离最大,为0.11,厦门群体与惠安群体间的遗传距离为0.09(表6)。
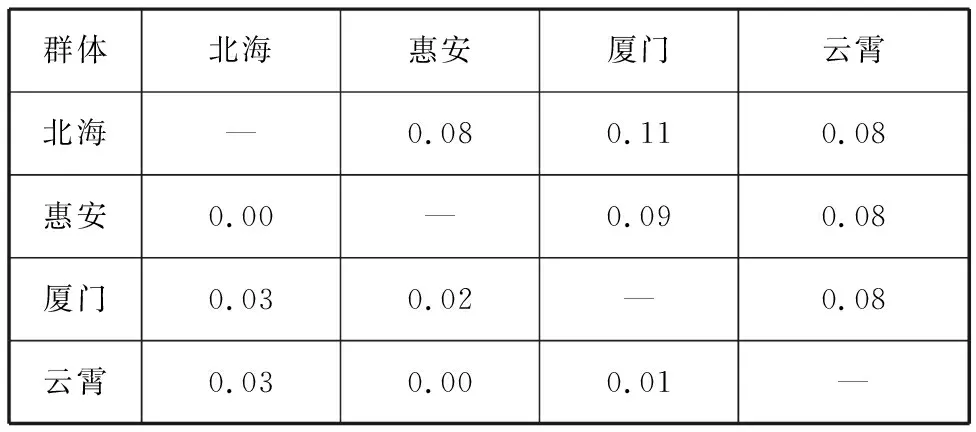
表6 波纹巴非蛤4个群体间遗传分化指数与遗传距离Tab. 6 Genetic differentiation index and genetic distance among four populations of Paratapes undulatas
基于4个波纹巴非蛤群体之间的遗传距离构建NJ系统发育树(图5),可知北海群体首先与惠安群体聚类,再与厦门和云霄群体聚类,未检测到与地理位置相对应的谱系结构。
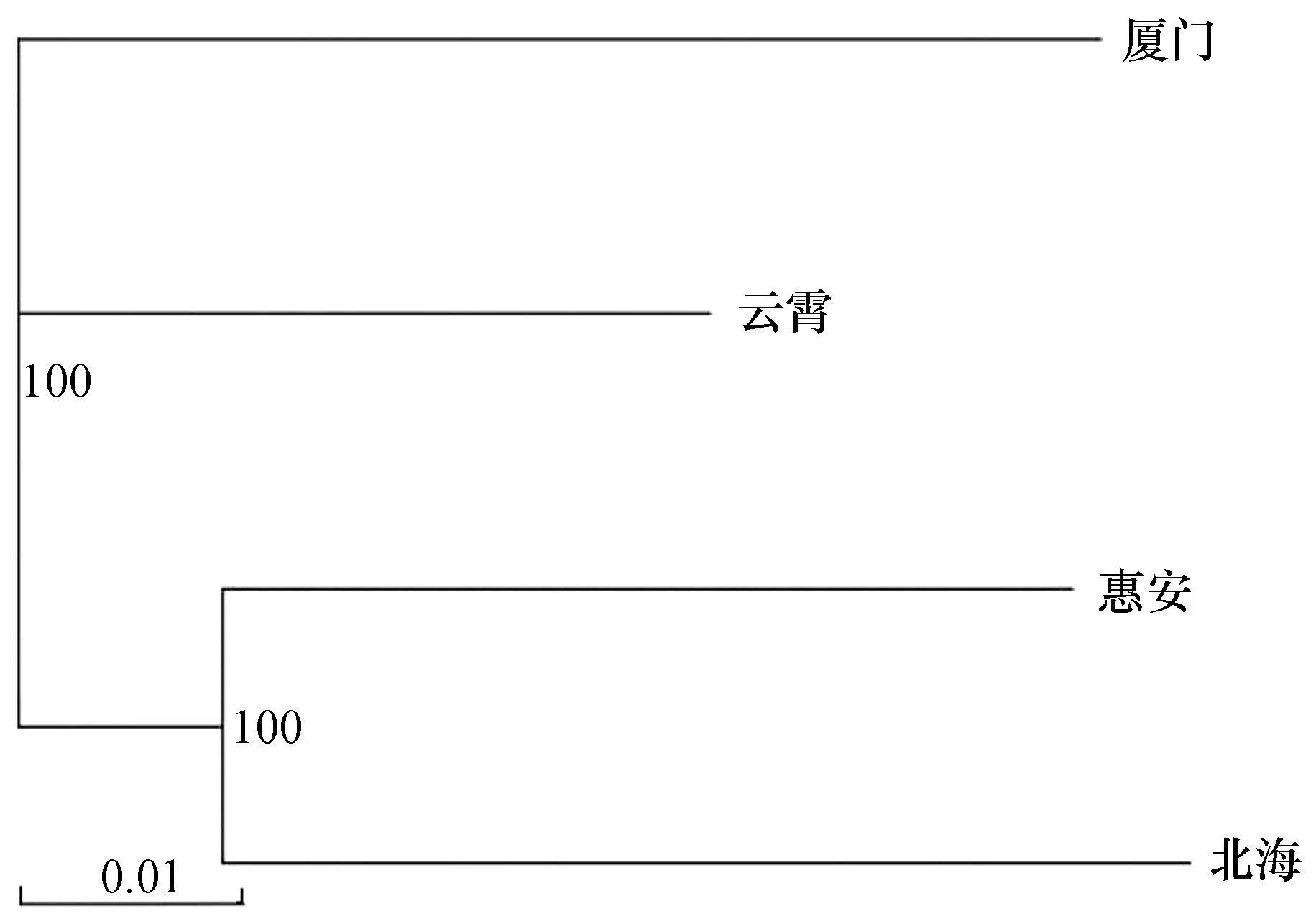
图5 基于遗传距离构建的四个波纹巴非蛤群体NJ系统发育树Fig. 5 NJ phylogenetic tree of four Paratapes undulatas populations based on genetic distance
2.2.3 自由交配估计
运用Structure软件,根据等位基因频率不相关假设(allele frequencies independent assumption)[26],进一步确认波纹巴非蛤群体遗传结构。首先假设自由交配组群(K)可能为1~9,对每个假设自由交配组群进行50次独立重复运算,最后使用最小二乘法估算出最佳自由交配组群(最小交叉熵值)为K= 1(图6)。
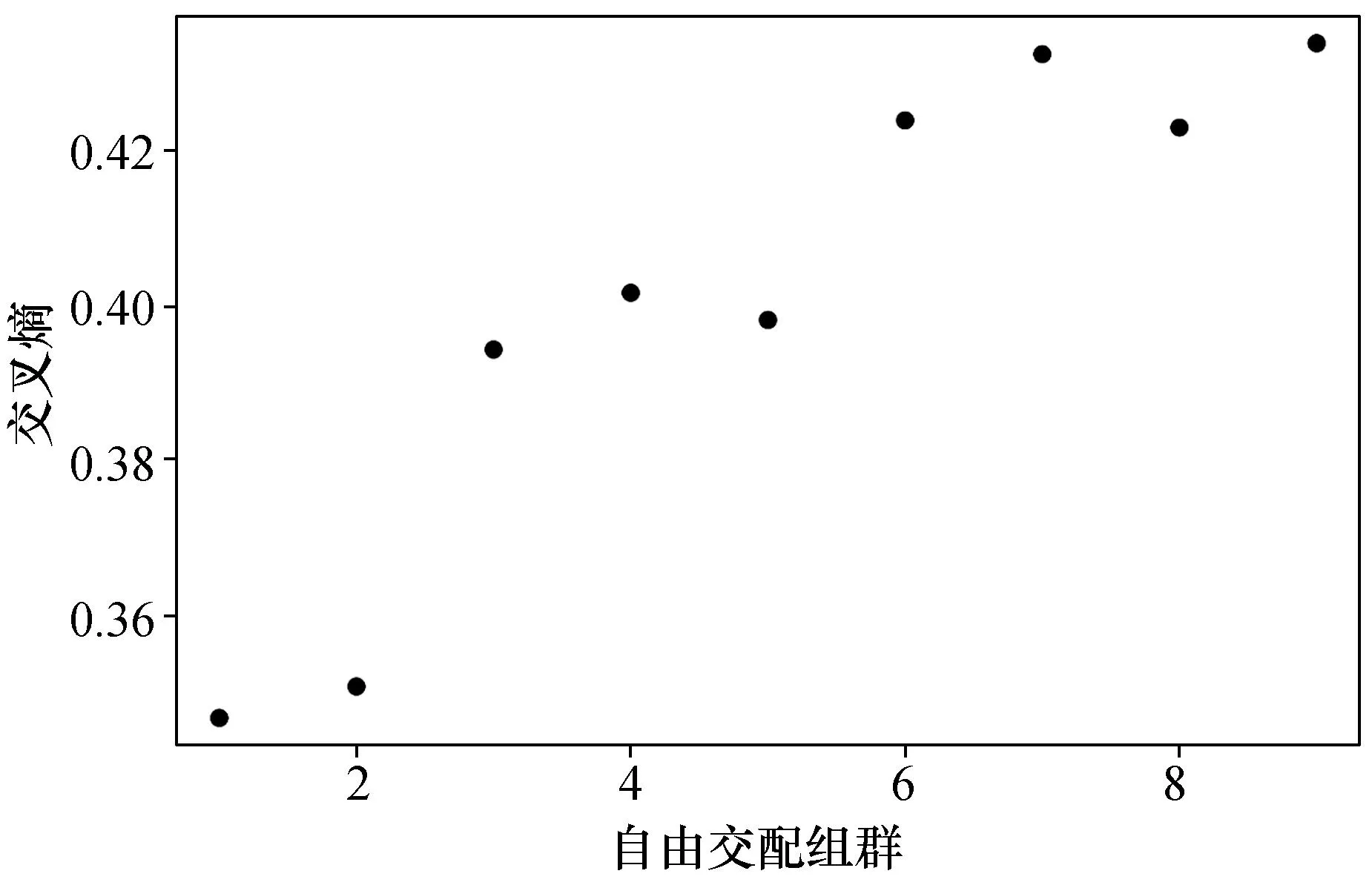
图6 波纹巴非蛤四个群体的交叉熵值Fig. 6 Cross entropy of four Paratapes undulatas population
2.2.4 瓶颈效应检测分析
利用BOTTLENECK软件检测波纹巴非蛤4个群体近期是否经历过瓶颈效应事件(表7)。首先基于IAM、TPM和SMM 3种不同模型进行Wilcoxon检验。结果显示在TPM模型下,所有波纹巴非蛤群体均未出现偏离突变-漂变平衡的现象,而在IAM模型下北海群体存在偏离现象,在SMM模型下厦门与云霄群体出现偏离现象。其次,Mode shift检验结果显示,4个群体的等位基因分布频率均呈正常的L型分布。TPM是介于IAM和SAM之间的模型,其突变大多为逐步突变,TPM和SMM模型描述的位点突变模式更接近微卫星的突变机理,因此TPM模型更适合微卫星数据的分析[27]。综上所述,4个波纹巴非蛤群体近期未经历过瓶颈效应事件。
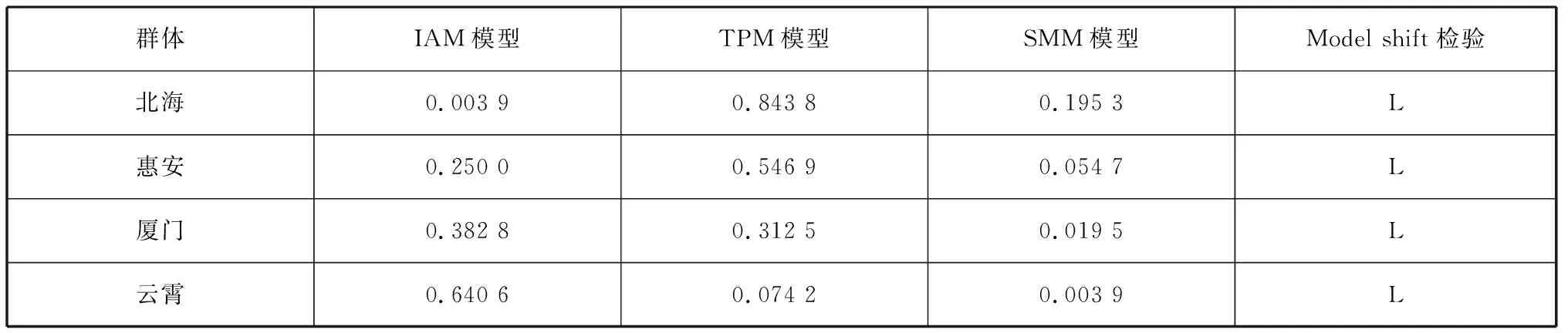
表7 基于3种模型的波纹巴非蛤4个群体的瓶颈效应检测Tab. 7 Bottleneck effect detection of four Paratapes undulates populations based on three models
3 讨论
3.1 4个自然种群波纹巴非蛤形态差异比较分析
贝类养殖已成为海洋经济产业中重要的一环,其遗传学相关研究也越来越受到生物学家的重视[28]。研究表明,在基因交流阻隔的情况下,同一种类生物的不同地理群体也会出现较明显的遗传差异[29]。我们分析了福建闽南地区及广西北海地区的波纹巴非蛤地理群体的多种形态学参数,可知惠安与北海群体的形态差异最小,且与云霄、厦门群体之间均存在着一定的形态差异。这种差异一方面可能是由地理因素、环境条件决定的,但聚类分析与主成分分析结果均表明惠安地区与北海地区的差异远比云霄和厦门小。造成这一结果的原因可能是惠安大港湾近岸海域近年来大量引进北海地区波纹巴非蛤种苗,从事水产增养增殖活动。这些人为商业活动可能使群体间种质交叉增多,向当地自然海区释放,从而大大增加了群体间基因杂交的概率[30]。另外,形态差异除了种质本身的特性外,与饵料种类、水文水质、底质类型等因素也有较大关系[31-32]。
3.2 4个自然种群波纹巴非蛤分子遗传学比较分析
传统的贝类遗传学研究集中在其形态指标上,但形态参数可能存在误差,并不能完全准确地反映物种的群体特性,因此需要结合分子生物学方法,获得更加真实稳定的遗传学研究结果[33]。我们用分子生物学手段调查了福建闽南地区及广西北海的波纹巴非蛤群体样本,其遗传多样性均呈现较高的水平(多态信息含量指数>0.5),与骆轩等的研究结果[10]基本持平,说明波纹巴非蛤群体仍具有较高的环境适应能力和进化潜力。波纹巴非蛤能维持如此高的遗传多样性,除了与其有大量的有效补充群体外,还与其未经历过瓶颈效应有关。3种突变模式对波纹巴非蛤的瓶颈效应检验均未发现杂合过剩现象,等位基因频率均呈正常的L型分布,在群体扩张的过程中等位基因丢失的概率较小,较多的遗传突变得以保存[34]。随着波纹巴非蛤资源的不断破坏和国内外需求的不断增加,养殖无法仅依靠海区捕捞的天然苗种得到满足,人工繁育大量兴起[35]。在后期养殖过程中,波纹巴非蛤作为主要的海水养殖贝类在不同养殖区间被频繁的人为引种、贸易及增殖放流。而在形态学和分子遗传学聚类分析时,发现厦门群体均独立形成一支,遗传分化指数和遗传距离厦门群体也略高于其他群体,这可能与厦门海域有不少国家级自然保护区,禁止捕捞和滩涂养殖,该群体可能保留了更多的原始遗传信息有关。
4 结论
本研究中4个波纹巴非蛤群体间遗传分化微弱,自由交配估计检验也显示所有群体为1个自由交配组群;NJ系统发育树聚类结果未检测到与地理位置相对应的谱系结构。相较于沿岸流输送波纹巴非蛤浮游期幼虫进行群体间的基因交流,不同地区间的人为引种、贸易及增殖放流等活动可能是造成波纹巴非蛤不同种群间基因频繁交流的主要原因,其中厦门群体则保留了更多的原始遗传信息。综合形态学和分子遗传学结果来看,在满足养殖需求的前提下,仍需要保护本地波纹巴非蛤的种质资源,防止外来群体的“生态入侵”。
猜你喜欢
收藏与投资(2023年12期)2024-01-03
当代作家(2023年3期)2023-04-23
北京航空航天大学学报(2021年6期)2021-07-20
阅读(低年级)(2021年10期)2021-03-08
少年漫画(艺术创想)(2019年10期)2019-04-20
红土地(2018年9期)2018-02-16
红土地(2018年9期)2018-02-16
中国军转民(2017年8期)2017-12-13
中国军转民(2017年8期)2017-12-13
学与玩(2017年5期)2017-02-16
