
miRNA-589-5p调控氧糖剥夺人脑微血管内皮细胞损伤的机制
2024-03-22 08:39王欣丽甄娜杨海龙
中国老年学杂志 2024年6期
王欣丽 甄娜 杨海龙
(衡水市人民医院,河北 衡水 053000)
脑小血管病发病隐匿,轻者可表现为认知功能障碍,重者可发展为腔隙性脑梗死、脑出血、脑卒中〔1,2〕,其发病机制与各种原因引起的内皮或血-脑屏障功能受损有关〔3~5〕。微小核糖核酸(miRNA)是一种广泛参与人类多种疾病的内源性稳定单链非编码小分子〔6〕,而miRNA-589-5p是新发现的参与内皮细胞损伤的miRNA,关于其对人脑微血管内皮细胞(HBMEC)功能的研究较少。本研究旨在探究miRNA-589-5p对氧糖剥夺诱导的HBMEC损伤的调控机制及其与组蛋白脱乙酰酶(HDAC)3、核因子E2相关因子(Nrf)2/血红素加氧酶(HO)-1信号通路之间的关系。
1 材料与方法
1.1材料 HBMEC购自中国上海科学研究院细胞库;改良Eagle培养基(DMEM)培养液购自美国Invitrogen;胎牛血清购自浙江天杭生物科技股份有限公司;Lipofectamin3000转染试剂购自赛默飞世尔科技(中国)公司;真核表达载体pcDNA3.1均购自Promega公司;RNA抽提试剂盒、实时荧光定量聚合酶链反应(RT-PCR)试剂盒均购自日本Takara公司;乳酸脱氢酶(LDH)检测试剂盒、RIPA裂解液购自上海碧云天生物技术研究所;膜联蛋白Ⅴ-异硫氰酸荧光素(Annexin Ⅴ-FITC)/碘化丙啶(PI)细胞凋亡检测试剂盒购自上海贝博生物科技公司;双荧光素酶报告基因检测试剂盒购自上海百赛生物技术股份有限公司;封闭专用脱脂奶粉购自北京普利莱基因技术有限公司;空白对照的miRNA序列(miRNA-con)、miRNA-589-5p模拟物的miRNA序列(miRNA-589-5p mimics)、沉默空白对照序列(si-con)、沉默HDAC3序列(si-HDAC3)、阴性对照组的miRNA抑制剂序列(anti-miRNA-con)、miRNA-589-5p抑制剂的miRNA序列(anti-miRNA-589-5p)及引物的设计合成均委托上海吉玛制药技术有限公司。鼠抗HDAC3单抗、兔抗裂解的半胱氨酸蛋白酶(Cleaved Caspase)-3 单抗、兔抗B细胞淋巴瘤(Bcl)-2相关X蛋白(Bax)多抗、兔抗Bcl-2多抗、兔抗NRF2多抗、兔抗HO-1多抗、HRP标记的二抗购自上海艾博抗贸易有限公司;Varioskan LUX型多功能酶标仪购自美国赛默飞世尔科技(中国)有限公司;Bio-Rad ZE5型智能流式分析系统购自美国伯乐生命医学产品有限公司。
1.2HBMEC的培养 10%胎牛血清+1%双抗混合的DMEM培养液,置于37 ℃、5%CO2饱和湿度恒温细胞培养箱中常规培养,2 d更换1次新鲜培养液。
1.3细胞转染与分组 将正常培养的HBMEC分为对照组(不做任何处理的HBMEC细胞)、氧糖剥夺组、氧糖剥夺+miRNA-con组、氧糖剥夺+miRNA-589-5p组、氧糖剥夺+si-con组、氧糖剥夺+si-HDAC3组、氧糖剥夺+miRNA-589-5p+pcDNA组、氧糖剥夺+miRNA-589-5p+pcDNA-HDAC3组、miRNA-con组、miRNA-589-5p组、anti-miRNA-con组、anti-miRNA-589-5p组。氧糖剥夺组细胞的处理〔7〕:将10%胎牛血清+1%双抗混合的DMEM培养液,置于37 ℃、5%CO2饱和湿度恒温细胞培养箱中培养的HBMEC更换为无糖DMEM,94%N2、1%O2、5%CO2的37 ℃恒温细胞培养箱中培养30 min。使用5倍DNA或质粒质量的Lipofectamin3000脂质体与DNA或质粒混合,将miRNA-con、miRNA-mimics、si-con、si-HDAC3、miRNA-589-5p+pcDNA、miRNA-589-5p+pcDNA-HDAC3依次转染至HBMEC,转染12 h,更换培养基进行氧糖剥夺处理。使用RT-PCR或Western印迹实验检验转染的效果。
1.4RT-PCR检测细胞中miRNA-589-5p、HDAC3表达 用RNA抽提试剂盒提取细胞总RNA,反转录试剂盒将其合成cDNA,做模板。用RT-PCR试剂盒,取2 μl cDNA与上游引物、下游引物、水及SYBR Premix Ex TaqTM在Mx3005P RT-PCR仪上进行扩增,记录循环数(Ct)值。以U6、GAPDH为内参,2-ΔΔCt法计算miRNA-589-5p、HDAC3相对表达量。ΔCt目的基因=Ct实验组目的基因-Ct对照组目的基因,ΔCt内参=Ct实验组内参-Ct对照组内参,-ΔΔCt=-(ΔCt目的基因-ΔCt内参)。反应程序为95 ℃,2 min;95 ℃,30 min;58 ℃,30 s;72 ℃,30 s;72 ℃,4 min,45个循环。miRNA-589-5p上游引物5′-CGAGGTCAGCGTGATTTCATGG-3′,下游5′-TGTGTCCAAGTCCCAGCCAGAG-3′;U6上游引物5′-CTCGCTTCGGCAGCACA-3′,下游5′-AACGCTTCA-CGAATTTGCGT-3′;HDAC3上游引物5′-GGAGCTGGACACCCTATGAA-3′,下游5′-TATTGGTGGGGCTGACTCTC-3′;GAPDH上游引物5′-GACAACAGCCTCAAGATCATCAG-3′,下游5′-GTGGCAGTGATGGCATGGA-3′。
1.5Western印迹检测细胞中HDAC3、Cleaved Caspase-3、Bax、Bcl-2、NRF2和HO-1蛋白表达 将细胞放在冰上用RIPA裂解液处理30 min,提取总蛋白。检测蛋白浓度,100 ℃沸水浴10 min变性,用上清做蛋白电泳样本。取50 μg上清至于上层分离胶梳子孔,低压温流电泳至分离胶低端,换成高压继续电泳直至蛋白到达浓缩胶底端。使用转膜仪将胶上的蛋白转移至聚偏氟乙烯(PVDF)膜,整个装置维持在0 ℃。结束后,封闭液(含2.5%脱脂奶粉)37 ℃,2 h封闭膜。电泳液充分洗膜,滴加稀释的一抗(鼠抗HDAC3单抗,1∶1 000;兔抗Cleaved Caspase-3 单抗,1∶2 000;兔抗Bax、Bcl-2、NRF2、HO-1多抗,1∶1 000),37 ℃,摇床孵育3 h,充分洗膜。滴加稀释的二抗溶液〔辣根过氧化物酶(HRP)标记的二抗,1∶500〕,37 ℃孵育2 h。洗膜后滴加ECL发光液,显影定影曝光。使用Image J软件分析蛋白的灰度值,目的蛋白灰度与内参蛋白(GAPDH)灰度之比表示目的蛋白相对表达水平。
1.6酶联免疫吸附试验(ELISA)检测细胞上清LDH 将细胞4 ℃低温12 000 r/min离心10 min,取上清。按照LDH试剂盒要求操作,检测细胞上清中LDH的相对表达情况。
1.7Annexin Ⅴ-FITC检测细胞凋亡 将细胞先用PBS洗涤5次,再用结合缓冲液悬浮细胞,待用。取500 μl的悬浮细胞,加入10 μl的Annexin Ⅴ-FITC避光孵育15 min,再加入10 μl的PI避光孵育20 min,结束后,将细胞用300目筛过滤,立即置流式细胞仪检测分析细胞的凋亡情况。细胞总凋亡率(%)=早期凋亡率(%)+晚期凋亡率(%)〔8〕。
1.8双荧光素酶报告基因检测实验 在线预测网站Starbase(https://starbase.sysu.edu.cn/)预测miRNA-589-5p的靶基因发现,HDAC3是miRNA-589-5p的一个靶基因。根据预测到的结合位点化学合成含有结合位点的片段(HDAC3-WT)和不含结合位点的片段(HDAC3-MUT),插入荧光载体,构建荧光报告基因。将荧光报告基因与miRNA-con、miRNA-589-5p共转染至细胞。按照双荧光素酶报告基因检测试剂盒说明书要求操作,检测分析细胞的荧光活性。用海参荧光素酶的活性与荧火虫荧光素酶的活性的比值表示细胞的荧光活性。
1.9统计学处理 采用SPSS22.0软件进行t检验、方差分析。
2 结 果
2.1miRNA-589-5p和HDAC3在HBMEC细胞中的表达 与对照组相比,氧糖剥夺组细胞中miRNA-589-5p表达水平显著降低,HDAC3 mRNA和蛋白表达均显著升高(均P<0.001)。见图1、表1。
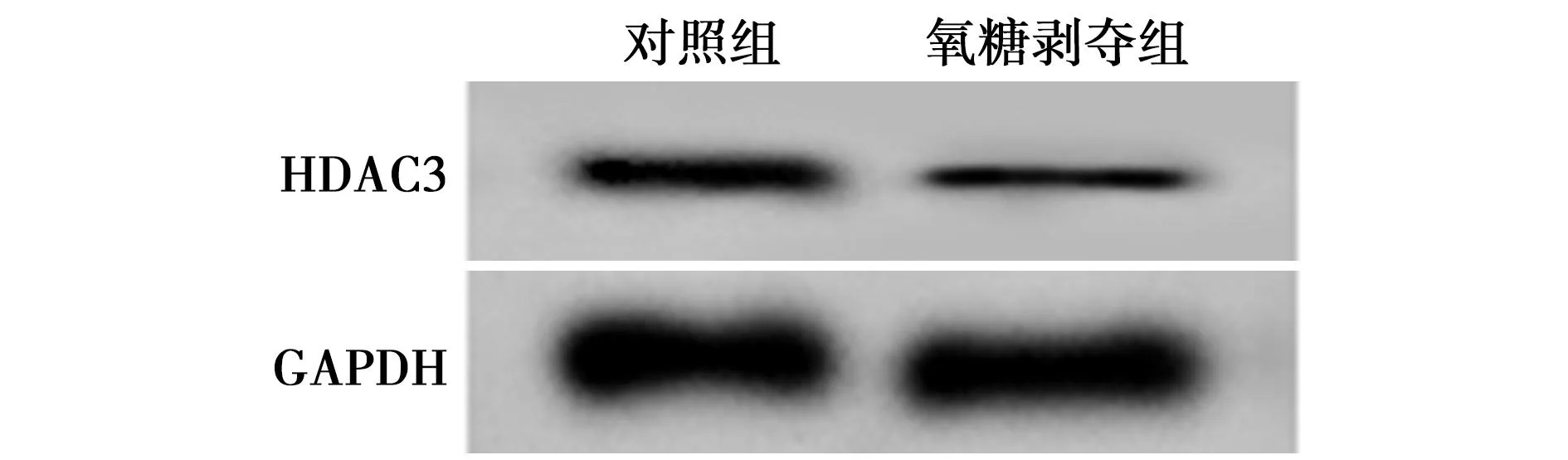
图1 HBMEC细胞中HDAC3的蛋白表达
表1 检测HBMEC细胞中miRNA-589-5p、HDAC3的mRNA和HDAC3蛋白的表达
2.2过表达miRNA-589-5p对氧糖剥夺诱导HBMEC细胞损伤、凋亡的影响 与对照组相比,氧糖剥夺组细胞中miRNA-589-5p表达、Bcl-2蛋白表达显著降低,LDH、Cleaved Caspas-3、Bax蛋白表达、细胞凋亡率显著升高(P<0.05);与氧糖剥夺+miRNA-con组相比,氧糖剥夺+miRNA-589-5p组细胞上述指标均反向变化。可见,过表达miRNA-589-5p减轻氧糖剥夺诱导HBMEC细胞的氧化应激和凋亡。见图2、图3、表2。
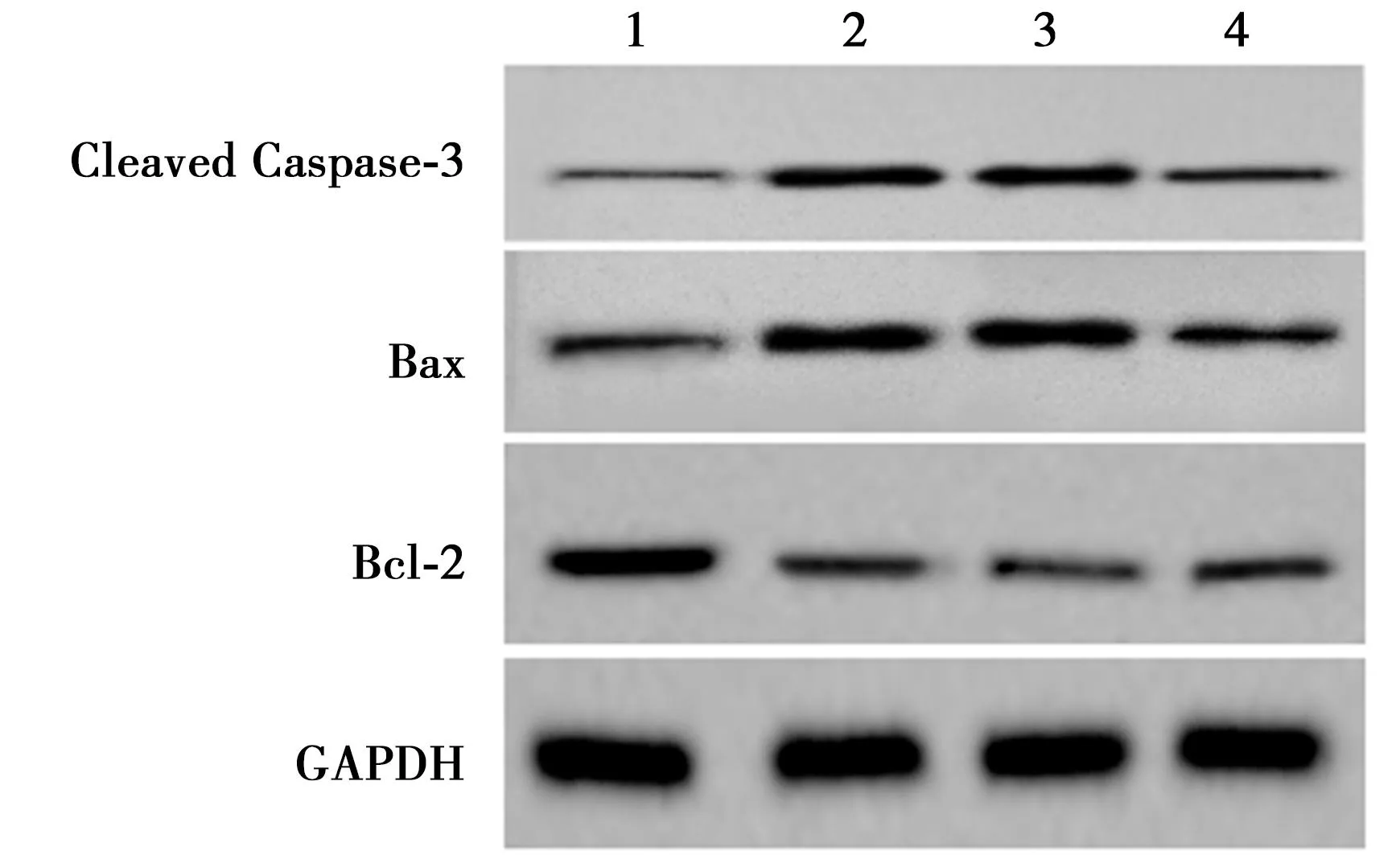
1~4:对照组、氧糖剥夺组、氧糖剥夺+miRNA-con组、氧糖剥夺+miRNA-589-5p组

图3 各组HBMEC细胞凋亡
表2 过表达miRNA-589-5p的HBMEC细胞中LDH表达及Cleaved Caspase-3、Bax、Bcl-2蛋白表达和凋亡率的影响
2.3miRNA-589-5p靶向HDAC3 通过靶基因在线预测软件预测到HDAC3的3′UTR存在与miRNA-589-5p互补的8个核苷酸序列(图4)。双荧光素酶报告基因检测实验结果如表3所示,miRNA-589-5p组HDAC3-WT细胞的荧光活性显著低于miRNA-con组(P<0.05)。见表3。与miRNA-con组(0.41±0.08)相比,miRNA-589-5p组细胞HDAC3蛋白表达显著降低(0.24±0.05,P<0.05),与anti-miRNA-con组(0.44±0.05)相比,anti-miRNA-589-5p组细胞HDAC3蛋白表达显著升高(0.68±0.07,P<0.05)。见图5,可见miRNA-589-5p靶向负调控HDAC3。

图4 miRNA-589-5p 与HDAC3互补的结合位点
表3 双荧光素酶报告实验

1~4:miRNA-con组、miRNA-589-5p组、anti-miRNA-con组、anti-miRNA-589-5p组
2.4沉默HDAC3对氧糖剥夺诱导HBMEC细胞损伤、凋亡的影响 与对照组相比,氧糖剥夺组细胞中HDAC3、LDH、Cleaved Caspase-3、Bcl-2蛋白表达、细胞凋亡率显著升高,Bax-2蛋白表达显著降低(P<0.05);与氧糖剥夺+si-con组相比,氧糖剥夺+si-HDAC3组细胞的上述指标发生显著的反向调控。见图6、图7、表4。

1~4:对照组、氧糖剥夺组、氧糖剥夺+si-con组、氧糖剥夺+si-HDAC3组

图7 沉默HDAC3对氧糖剥夺诱导HBMEC细胞凋亡的影响
表4 沉默HDAC3对氧糖剥夺诱导HBMEC细胞中LDH表达及Cleaved Caspase-3、Bax、Bcl-2蛋白表达和凋亡率的影响
2.5过表达HDAC3部分逆转过表达miRNA-589-5p对HBMEC细胞的保护作用 与氧糖剥夺+miRNA-589-5p+pcDNA组相比,氧糖剥夺+miRNA-589-5p+pcDNA-HDAC3组细胞miRNA-589-5p表达显著降低,Bcl-2蛋白表达、细胞凋亡率、HDAC3、LDH、Cleaved Caspase-3、Bax蛋白表达显著升高(P<0.05)。见图8、表5。

1,2:氧糖剥夺+miRNA-589-5p+pcDNA组、氧糖剥夺+miRNA-589-5p+pcDNA-HDAC3组
表5 沉默HDAC3部分逆转过表达miRNA-589-5p对HBMEC细胞LDH表达及Cleaved Caspase-3、Bax、Bcl-2蛋白表达和凋亡率的影响
2.6miRNA-589-5p和HDAC3表达对HBMEC细胞NRF2/HO-1信号通路的影响 与对照组相比,氧糖剥夺组细胞NRF2核蛋白/PCNA、HO-1蛋白表达均显著降低(P<0.05);与氧糖剥夺+miRNA-con组相比,氧糖剥夺+miRNA-589-5p组细胞NRF2核蛋白/PCNA、HO-1蛋白表达均显著升高(P<0.05);与氧糖剥夺+miRNA-589-5p+pcDNA组相比,氧糖剥夺+miRNA-589-5p+pcDNA-HDAC3组细胞NRF2核蛋白/PCNA、HO-1蛋白表达均显著降低(P<0.05)。见图9、表6。
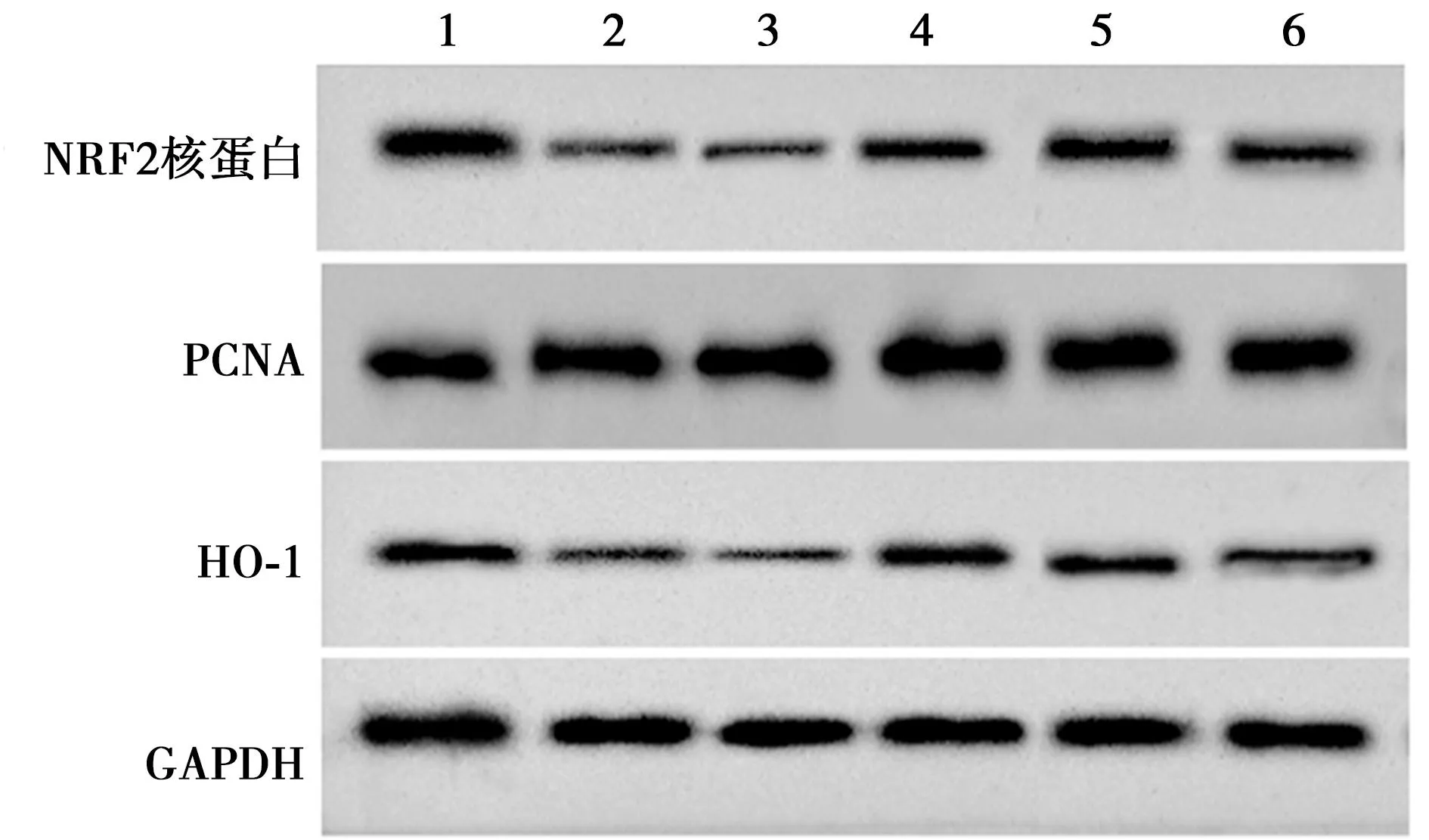
1~6:对照组、氧糖剥夺组、氧糖剥夺+miRNA-con组、氧糖剥夺+miRNA-589-5p组、氧糖剥夺+miRNA-589-5p+pcDNA组、氧糖剥夺+miRNA-589-5p+pcDNA-HDAC3组
表6 沉默HDAC3部分逆转过表达miRNA-589-5p对HBMEC细胞NRF2和HO-1蛋白表达的影响
3 讨 论
miRNA-589-5p在人类癌症中的功能研究比较丰富,其在肝癌、前列腺癌、喉癌、肾癌等实体瘤中均扮演抑癌基因的角色〔9〕。但是miRNA-589-5p在血管内皮细胞中的作用研究甚少。Yu等〔10〕发现,miRNA-589-5p在高糖诱导的视网膜微血管内皮细胞中低表达,作为环状RNA UBAP2的下游吸附因子和EGR1的上游靶基因调控高糖诱导的视网膜微血管内皮细胞的氧化应激、增殖和迁移。Yu等〔11〕通过氧化低密度脂蛋白(ox-LDL)诱导的HBMEC细胞构建脑血管细胞损伤模型发现,miRNA-589-5p的表达明显上调,抑制miRNA-589-5p能够抵抗ox-LDL对HBMEC细胞的损伤作用。miRNA-589-5p在血管内皮细胞中的功能尚处于研究初期,其具体的功能仍有待继续探究。本研究与Yu等〔11〕研究结果相悖,与Yu等〔10〕研究结果相吻合。进一步研究通过双荧光素酶报告基因检测实验发现,miRNA-589-5p能够靶向负调控HDAC3的表达,猜测这可能与miRNA-589-5p抵抗氧糖剥夺诱导的HBMEC细胞损伤有关系。
有研究报道,抑制HDAC3能够通过提高PPARγ的活性明显改善氧糖剥夺-复氧诱导的HBMEC细胞的跨内皮通透性增加〔12,13〕。Lin等〔14〕研究发现,HDAC2在发生卒中后表达上调,而环境富集(EE)有益于脑卒中的恢复,并抑制HDAC2的上调,过表达HDAC2则可部分逆转EE的恢复卒中功能,敲除HDAC2增强神经营养素和神经可塑性,说明HDAC2对于EE的功能恢复至关重要,精准靶向HDAC2可能成为中风恢复的新疗法。Liao等〔15〕报道,小胶质细胞中HDAC3的缺失可以减轻缺血再灌注(I/R)诱导的神经炎症和脑损伤,究其原因为HDAC3/p65/cGAS/STING信号通路参与I/R诱导的神经炎症的发展,为缺血性脑卒中的治疗提供新方向。本研究结果也显示,过表达HDAC3部分逆转了过表达miRNA-589-5p对氧糖剥夺诱导HBMEC细胞的保护作用。内皮细胞的损伤离不开氧化应激引起的细胞凋亡,而对氧化应激具有保护作用的药物多数是通过Nrf2/HO-1信号通路的激活抑制内皮细胞的氧化应激和凋亡〔16,17〕。Yang等〔18〕研究发现,高糖能够显著降低Nrf2和HO-1的表达,降低线粒体膜电位,增加细胞内和线粒体活性氧(ROS)的生成,诱导BMECs的细胞骨架损伤和凋亡,线粒体ROS清除剂处理显著上调HG诱导的BMEC中Nrf2和HO-1的表达,这伴随着线粒体膜电位的提高和ROS的减少,表明线粒体ROS清除剂通过激活Nrf2/HO-1信号通路对高糖诱导的BMECs损伤具有保护作用。本研究说明,NRF2/HO-1信号通路参与miRNA-589-5p的改善氧糖剥夺诱导HBMEC细胞损伤的过程。本研究结果仅在体外得到初步证实,缺乏体内的动物实验研究。
综上,miRNA-589-5p改善氧糖剥夺HBMEC的损伤,产生这种作用的机制可能与靶向HDAC3,激活NRF2/HO-1信号通路的活性有关,为脑小血管病的治疗提供新思路。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
中国眼镜科技杂志(2019年9期)2019-11-11
安徽医科大学学报(2016年12期)2017-01-15
中国药理学通报(2015年2期)2016-01-12
中国病理生理杂志(2015年8期)2015-12-21
热带农业科学(2015年9期)2015-10-14
中国医学科学院学报(2015年5期)2015-03-01
中国医药导报(2015年27期)2015-02-28
