
泥河湾盆地马圈沟遗址哺乳动物破碎长骨反映的古人类行为
2024-04-16 20:11王晓敏刘连强陈国鹏李锋谢飞高星
人类学学报 2024年1期
王晓敏 刘连强 陈国鹏 李锋 谢飞 高星

关键词:早更新世;泥河湾盆地;埋藏学;破碎长骨;人类行为
1引言
第一批由非洲向欧亚大陆扩散的古人类遭遇了各种各样的生态环境,与他们在非洲生存时所适应的环境有显著差别。因此,早更新世古人类如何适应本地环境并获取能量继以生存,一直是古人类学和考古学关注的热点。对于有动物化石出土的早更新世遗址来说,如何从动物遗骸中知晓古人类获得和使用动物资源的方式是研究者最关心的问题,而这一问题的本质就是识别猎物的首要消费者(primaryaccesstocarcasses,即优先获取并改造猎物的摄食者)与次要消费者(secondaryaccesstocarcasses,即在首要消费者之后获取并改造猎物的摄食者)。早更新世考古遗址中哺乳动物化石聚集的重要埋藏营力主要来自于食肉动物与古人类,当然,这一过程也受到了其他自然作用的影响。想要探究古人类获取动物资源的能力,就需要有效地区分古人类和食肉动物利用动物性资源的先后顺序,进而识别出古人类利用动物资源的方式。在20世纪60年代以前,研究者们普遍认为古人类是天生的狩猎者,考古遗址的动物遗骸都是古人类猎取并食用后留下的,他们甚至可以轻松地猎取体型巨大的动物,如象和犀[1]。Binford首次提出非洲奥杜威时期(Oldowan,距今2~0.7Ma)一些考古遗址出土的动物骨骼是食肉类动物消费的残余,早期人类只是捡食了残存的肢骨[2]。一些研究者通过分析这些遗址的动物骨骼各解剖部位的出现频率及骨骼表面痕迹,支持了Binford的观点,并提出非洲早更新世古人类是机会主义食腐者(opportunisticscavengers)[3]。近年来,随着研究材料的进一步丰富及实验考古相关数据的积累,大多数研究倾向于认为早更新世古人类应该是猎物的首要消费者,而骨骼所经受的食肉类改造大多发生在他们将动物遗骸抛弃之后[4,5]。直到现在,随着研究方法的不断修正以及研究数据的不断积累,研究者对早更新世古人类行为特点的认识也在不断地调整,有关狩猎还是食腐的问题仍有争议。
亚洲高纬度地区最早的、有确切人类活动的考古记录来自于中国的泥河湾盆地、上陈、西侯度以及格鲁吉亚的德马尼西等遗址(Dmanisi,44°20′N,距今1.78~1.85MaBP)[6-9]。动物考古学的研究显示,德马尼西遗址的古人类生活在相对暖湿的环境之中,栖居地周围的动物资源丰富[10]。在泥河湾盆地发现的大量早更新世考古遗址中,小长梁、大长梁、半山、马圈沟遗址的测年结果显示,至少在1.66MaBP以前,古人类已经在泥河湾盆地活动[6,11-13]。比较普遍的观点认为,这意味着早期人类能够在中国北方高纬度地区长期生存,他们适应环境,特别是在寒冷干燥气候下的适应能力很强,而这种适应性策略最重要的行为特征之一就是经常性地猎取和消费大型哺乳动物,尤其是适应寒冷气候的草原猛犸象[14,15]。然而,也有学者提出,泥河湾盆地并没有确切的早期人类主动获取大型动物的证据,当地古人类应对干冷气候的策略应该是向南迁徙;与之伴生的草原猛犸象在中国北方最初的记录年代处于相对温暖的气候阶段,这说明泥河湾盆地至少在秋冬季节并不适宜早期人类生活[16]。
在泥河湾盆地的一些早更新世遗址,研究人员虽然开展了一些动物骨骼的研究,但并不系统,所解决的科学问题也比较局限。这种情况一方面受制于研究手段的缺乏;另一方面也是由于这些遗址出土的动物骨骼保存状况欠佳,所能提取的信息有限。1990年中美合作发掘东谷坨遗址之后,美方研究者简单报道了该遗址动物化石的情况并做了初步鉴定,但認为该遗址的骨骼过于破碎而无法说明更多的问题[17]。Peterson等[18]通过对小长梁遗址的埋藏学研究发现,该遗址动物化石不大可能是古人类和食肉动物消费后的堆积;相反,由于接近50.4%的动物骨骼表面都有明显的水流磨蚀痕迹的出现,因此这一动物群化石很可能是因流水搬运而形成的次生堆积。Keates[19]报道了马圈沟遗址第3文化层出土动物化石的基本情况,但没有继续开展相关动物骨骼的研究。本文以马圈沟遗址第3文化层出土的部分动物化石为研究材料,基于骨骼保存的状况,从长骨的破碎方式入手,利用多变量的骨骼破碎信息,特别是断口破裂的角度,阐明提取碎骨信息的必要性,并讨论了这些信息所反映的古人类与食肉动物的互动关系。
2第3文化层
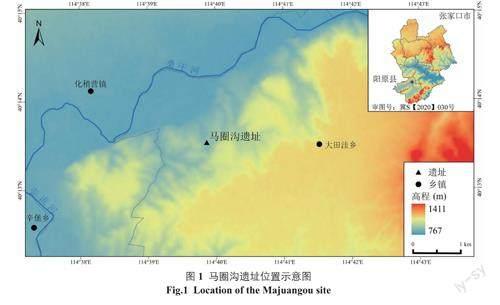
马圈沟遗址是泥河湾盆地年代最早的古人类遗址,是东亚地区年代最久远的旧石器时代遗址之一。该遗址位于河北省阳原县,地处泥河湾盆地东部,大田洼台地北部边缘,地理坐标为40°13′31.1″N、114°39′51.0″E,海拔约840m[15](图1)。
马圈沟遗址发现于1992年春,发掘者于同年清理了文化层以上的地层堆积。次年9~10月,谢飞、李珺、成胜泉等对该遗址进行首次发掘,发掘对象是第1文化层。2000年,河北省文物研究所(现河北省文物考古研究院)对该遗址进行了再次调查,于已发掘文化层下方发现第2、3文化层;后又于2001~2003年,对第2、3文化层分别进行了三次发掘[15]。2005年,考古队继续在第3文化层的底面向下发掘,找到了年代更早的第4、5、6三个文化层。2014~2019年,河北省文物研究所等单位在原马圈沟遗址核心区域以外开展了早期人类遗存的调查和发掘,不仅扩展了原先发掘的第3~6文化层的区域,并且先后发现并发掘了马圈沟遗址D区、E区、鱼咀沟1号地点、鱼咀沟2号地点及棋盘梁地点。至此,马圈沟遗址的材料越来越丰富,扩展为包括马圈沟遗址、马圈沟遗址D区、半山遗址、鱼咀沟1号地点、鱼咀沟2号地点、棋盘梁地点等8个地点的早更新世考古遗址群。
马圈沟是泥河湾湖滨由于水流作用而产生的冲沟。第3文化层厚约33cm,为灰黑色黏土层。朱日祥等根据古地磁年代学的研究推测第3文化层的年代为1.66MaBP[6];蔡保全等则通过生物地层学研究推断第3文化层年代应早于距今1.8MaBP[20]。随后,刘春茹团队使用电子自旋共振(ESR,ElectronSpinResonance)测年法,估测第3文化层年代约为1.7MaBP[21]。Duval指出该研究在实验方法和数据公开方面可能存在一些问题,但并不否认该测年方法的有效性[22]。在此年代范围内,孢粉数据显示泥河湾盆地和中国北方地区气候以暖温带、温带针叶和阔叶混交林气候为主[23]。
第3文化层出土的石制品原料以质地细密的灰黑色燧石为主,有较为成熟的锤击法打片技术[15]。
马圈沟遗址出土了大量哺乳动物化石,其中,第3文化层的出土数量最多,数次发掘积累了数千件化石,现已鉴别出20余种。超大型哺乳动物中大部分为草原猛犸象(Mammuthustrogontherii),也有披毛犀(Coelodontaantiquitatis)的遗存。其他典型的哺乳动物包括三门马(Equussanmeniensis)、鬣狗(Pachycrocutasp.)、鹿(Cervussp.)和羚羊(Gazellasp.)等[20]。该哺乳动物群与小长梁遗址的典型动物群较一致[24]。有研究者报道,在马圈沟遗址出土的一件草原猛犸象肋骨和数件可能属于鹿或马类动物的肢骨骨干上发现了被认为是切割、砍砸、刮削的人工痕迹[6,14,15]。据此,不少学者提出在泥河湾盆地,可能早在1.60MaBP以前,古人类便能够捕猎并消费体型很大的哺乳动物[14]。但也有研究者提出,大部分骨骼表面可能存在磨损,可能影响人工痕迹的识别[6]。Dennell[16]便指出,马圈沟遗址骨骼表面痕迹鉴定的结果并不成熟,即使人工痕迹确实存在,也无法将当地早更新世古人类的狩猎和食腐行为区分开来。
为了进一步寻找区分古人类狩猎及食腐行为的证据,中国社会科学院考古研究所与河北省文物考古研究院及中国科学院古脊椎动物与古人类研究所合作,对马圈沟遗址出土的动物化石进行了更系统、细致的观测与分析。本文的研究材料主要来自2001~2003年马圈沟遗址第3文化层发掘出土的长骨残段及残片。
3研究方法
骨髓是油脂的重要来源,其富含的脂溶性维生素A、D、E和K是人体生长发育的重要营养元素[25]。人类和食肉动物都会打碎动物的长骨来获取骨髓,这种行为既针对被他们捕获的猎物,也经常发生在那些被他们发现的因其他原因死亡的动物遗骸上。古人类敲碎长骨获取骨髓的行为最早可追溯至2.6MaBP之前[26]。考古遗址经常大量地出土长骨的碎块/片,为研究古人类开发骨髓资源以及古人类与食肉动物的互动关系提供了丰富的材料。
在针对古人类遗址出土动物遗存的埋藏学研究方法中,有四个重要的手段用来识别古人类和自然作用的埋藏特征,分别是动物死亡年龄分布(mortalitypattern)、骨骼单元分布(skeletalelementprofiles)、骨骼表面痕迹(bonesurfacemodification)以及骨骼破碎方式(bonebreakagepattern)[27]。其中,前三项指标的观测与分析被认为是分辨上/更新世动物群中人类与食肉动物行为的基础手段,然而无论研究者如何定义古人类的行为所造成的埋藏特征,都不能完全排除食肉动物或其他自然作用去造成相似的特征(即等效现象,equifinality)[28]。一段时间以来,国内外学者通过详细的实验与实证考古研究以及考古案例分析,利用骨骼表面痕迹分析的手段尝试区分等效现象,取得了不错的效果。长骨上的砍砸痕、切割痕及食肉动物啃咬痕迹的识别与统计分析为确认古人类的“狩猎”行为提供了力证[29,30]。但是,由于各种各样的原因,很多遗址的骨骼表面并没有发现确切的痕迹。尤其对于早更新世的旷野遗址,古人类和食肉动物低强度的活动及遗址保存环境对骨骼保存状况的影响,都可能导致骨骼表面缺乏痕迹或者痕迹很难被识别出来。除了骨骼表面痕迹的研究以外,敲击长骨时产生的碎片也引起了一些学者的关注,但受限于研究材料,相关研究成果并不多[31]。
大多数考古遗址中的动物遗存都以无法鉴定的骨骼碎片的形式出现,想要从这些碎骨上提取信息并加以分析是一项耗时费力的工作,而提取的这些信息究竟是否能够提供分析古人类行为的证据,也令研究者们对分析碎骨的价值感到困惑。即便如此,国内外学者仍进行了大量的尝试和积累。骨骼破碎方式的研究方法也从简单的观察描述发展、简单的测量和统计发展到大数据处理,研究体系日趋系统与完善[32,33]。目前,国内已有多位学者对骨骼破裂特征的相关理论方法进行过综述,并在河南许昌灵井遗址[34]、贵州马鞍山遗址[35]、宁夏鸽子山遗址[36]及泥河湾盆地的于家沟遗址[37]进行了运用。
总的来说,研究骨骼破碎方式的落腳点在于骨骼断裂面的性状,涉及两条逻辑线索,一是判断骨骼在破裂前是否在新鲜的状态,以推断是否存在取食骨髓的可能性;二是统计骨骼破裂面的角度,以辨识取食骨髓的主体是人类还是食肉类。采集和分析数据常使用分类变量[38-43]。第二条线索的情况更复杂一些,针对其因果对应性的实验及争论更多[33]。
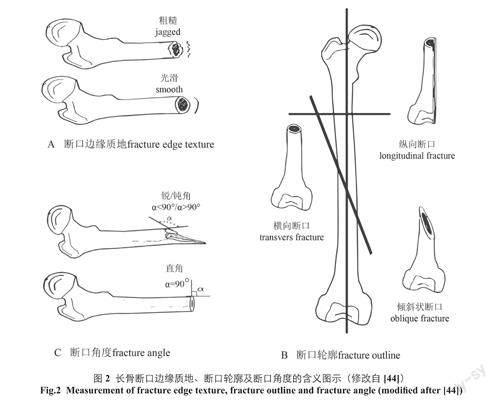
现阶段,研究中使用最广泛的方法体系是Villa和Mahieu[40]通过观察、测量及统计考古遗址出土的、不同状况下破碎的人类四肢骨骼而提出的三个变量,即断口边缘质地(fractureedgetexture)、断口轮廓(fractureoutline)和断口角度(fractureangle)。为了进一步明确这三个变量的用法,我们将它们的含义简述如下:断口边缘质地指长骨断口表面的纹理(图2:A)。通常有平滑和锯齿状两种:在新鲜状况下破裂的骨骼其断口常为平滑状,反之则常见锯齿状[40]。
断口轮廓指长骨断口与骨骼长轴的相对方向(图2:B)。通常有纵向(长骨断口与骨骼长轴平行)、横向(长骨断口与骨骼长轴垂直)和倾斜状(长骨断口与骨骼长轴呈角度倾斜)三种。倾斜断口的轮廓在空间上的形态更为多样,可能为平直且其延长线在平面上与长骨长轴相交,也可能为螺旋状且其在长骨纵剖面的投影与长骨长轴相交,非新鲜状况下破裂的骨骼常见横向断口[40,45,46]。过去,我们通常认为由人类利用工具砸击长骨导致的破裂呈螺旋状[47-49],但一些实验考古研究的结果表明,除了人类使用工具进行砍砸以外,其他埋藏因素(如食肉动物的牙齿)也会导致螺旋状的破裂[50]。
断口角度指骨骼破裂面与骨骼外表面的形成的角度(图2:C)。当这个角度近似直角时,一般表明骨骼是在非新鲜状况下破裂的[40]。
尽管Villa和Mahieu提出的几个变量确实能够可靠地反映骨骼最初破裂时的新鲜程度,并且能够与长骨骨髓被取食的行为联系起来[40],但取食骨髓的主角究竟是谁,是人类还是食肉类,其实并不明晰。于是,研究者们又着眼于三个变量中最复杂的一个变量,即断口角度进行了更细致的研究。
早在1985年,Johnson就提出人为造成的骨骼破裂是冲击性的,这种力量的传递在骨骼结构周围以不同的方式分配冲击能量,而食肉动物啃咬所爆发的力在骨骼表面的传导要均匀得多[51]。Capaldo和Blumenschine通过比较石锤砸击及食肉类啃咬的新鲜长骨碎片的断口角度,发现前者更接近钝角,而后者更接近直角[52]。他们认为,人类挥舞石锤敲击长骨的动作是一种动态冲击(dynamicloadingforce),力的传导更快速,爆发力更强,更容易破坏长骨骨密质的结构而形成倾斜的角度;而食肉类牙齿的啃咬是一种静态荷载(staticloadingforce),其施力的力矩和力臂都更小,更易形成没有倾斜角度的垂直状断口。Alcántara-García等人[53]通过对大型牛科动物的肢骨进行敲击实验对骨骼断口角度进行了区分,他们认为,在一般情况下,新鲜骨骼会沿着相对于其长轴倾斜和/或纵向的平面破裂,其破裂角度通常小于85°或大于95°。而干燥骨骼通常会以直角沿着横向和/或纵向平面断裂,并且具有不规则的破裂面,这与Villa和Mahieu的观察是基本一致的。另外,他们还在实验中发现,被人类锤击和食肉动物咀嚼破坏的新鲜骨骼样本中,沿着纵向平面断裂的断口角度,其范围有显著的重叠,而沿着横向平面断裂的断口角度也有很多重叠,尤其是在角度大于90°的大型动物骨骼标本上。与纵向和横向的破裂相比,人类和食肉动物造成的倾斜破裂面的破裂角度的重叠要小得多。进一步讲,在新鲜骨骼上由食肉动物造成的骨骼破裂角度通常在80°~110°(但通常不在85°~95°,这是干燥骨骼破裂的特征),而人类引起的骨骼破裂(尤其是倾斜状的断口)通常更尖锐(小于80°)或更钝(大于110°)。这些差异在统计学上是显著的,并且在倾斜状骨骼破裂上最为明显。而横向破裂面上的破裂角度的指示意义并不强。他们得到的实验结果几乎与Capaldo和Blumenschine的观察一致,后者也认为食肉动物造成的骨骼破裂角度比人类用石锤敲击的骨骼破裂角度更接近90°。
Pickering等人[32]和Domínguez-Rodrigo和Barba[5]依照这一结论对斯瓦特科兰斯(Swartkrans)和奥杜威峡谷(OlduvaiGorge)的FLKZinj(FLKZinjanthropusLevel)出土的长骨破裂面的角度进行了测量,并按骨骼破裂面的形态进行了分类统计和比较,推测长骨的破裂大部分是由于古人类的活动造成的,但这些骨骼同时也受到了食肉动物的侵扰。
DeJuana和Domínguez-Rodrigo利用马类动物骨骼进行了敲击实验,却观察到与Alcántara-García等人的部分数据相反的趋势[53-54]。在马骨上用石锤敲击产生的破裂角比在牛骨上更接近90°,并且与食肉动物造成的破裂角度更相近。然而,他们在实验中却没有将骨骼破裂的轮廓进行分类,而是把所有新鲜状况下破裂的骨骼作为一个整体进行观察,其结果并不利于开展比较研究。Coil等人[33]进一步细化了观察的对象,将解剖部位、动物年龄及骨骼是否冷冻这些参数加入变量之中。这些实验都表明,骨骼破裂角度的情况很复杂,受到很多变量的影响,这些变量可能是动物种属、骨骼部位、动物年龄及个体大小,等等。
4材料与方法
马圈沟遗址第3文化层出土的标本中,有部分草原猛犸象化石。较完整的标本以及之前报道有骨骼表面痕迹的标本都暂时陈列在泥河湾博物馆。加之超大型哺乳动物的骨骼破碎状况并不像其他更小体型的哺乳动物那样,有实验和考古研究结果进行对比,所以这些草原猛犸象的骨骼并不作为本文的材料。相应地,在破碎的长骨中,我们也排除了那些属于超大型哺乳动物的碎块,包括草原猛犸象及披毛犀。总体来说,本文关注的长骨残段及碎片共213件,既包括有可鉴定的长骨残段,也包含了不可以鉴定的长骨碎片。我们采集了这些标本的尺寸、风化等级、磨蚀状况、所属动物体型大小及骨骼破碎状况的信息。
标本的尺寸指长骨残最大长,直接用游标卡尺进行测量。长骨残段/片所属动物体型的大小主要依据该标本所属动物类别、解剖学部位及骨壁厚度進行划分。体型大小主要分为两类,为了便于与Alcántara-García的实验及斯瓦特科兰斯(Swartkrans)的研究数据进行对比,本研究参照Brain的研究[55],将那些体型在20~90kg范围内,包括与鹿及羚羊体型接近的动物,归类为中型动物;而体型在90~1000kg范围内并且与马和牛体型接近的动物,归类为大型动物。
在记录长骨的风化状况时,我们参考了Behrensmeyer[56]的六个风化阶段,分别是:0级,骨骼表面光滑,无风化裂痕;1级,与长轴平行的裂纹开始在骨表面出现;2级,骨骼表面开始出现片状剥离,剥离的骨皮仍与主体相连,边缘卷起;3级,片状剥离大面积出现,部分骨皮已经脱落;4级,骨骼表面呈粗糙的纤维状,风化作用已经影响到骨骼内部;5级,骨骼已风化破碎,原来骨骼的形态可能已较难辨认,往往暴露出海绵质。另外,我们也对长骨的磨圆状况进行了记录,主要针对有破碎断口的长骨,其中,把“标本的破裂面锋利”记录为未被磨圆。
基于前文所述的对骨骼破碎方式的研究方法的认识,我们首先识别出那些在新鲜状况下破裂的长骨。如果断裂面质地光滑,则视为新鲜状况下破裂的长骨。有一些断裂面可能受到埋藏作用的影响,其质地介于光滑与粗糙之间,对于这样的标本,如果其断裂的角度非常接近直角,则将其作用非新鲜状态下破裂的标本。识别出新鲜状况下破裂的标本之后,再将其断口分为横向、纵向以及倾斜三种类型,然后再测量这些断口的角度。然而,在实际的操作中我们发现,有些断口很难有效地分类,并且一些断口的角度并不总是均匀的。于是我们参考了斯瓦特科兰斯遗址出土的碎骨断口测量的方法[32],先选取中点进行测量,再沿中点整个断裂面至少取两点进行测量,最后取一个平均值。
5分析结果
5.1马圈沟第3文化层出土动物长骨化石的基本状况
长骨的残长是反映骨骼破碎状况的最基本数据信息。为了说明马圈沟遗址第3文化层长骨的破碎状况,并为将来与泥河湾盆地内其他早更新世遗址出土的动物骨骼进行基本信息的对比,我们以每20mm为一个范围,对100mm以内的骨骼尺寸进行了统计(图3)。结果显示,马圈沟遗址第3文化层残长大于100mm的长骨(n=59)最多,占总数的27.7%;其次为40~60mm的长骨(n=48),占22.5%左右;尺寸在20~40mm(n=41)以及60~80mm(n=42)的标本再次之,分别占19.2%及19.7%;尺寸小于20mm的长骨(n=2)最少,仅占0.93%。
我们对213件标本进行了观察,未发现5级风化等级的长骨标本(图4)。在这些长骨碎片标本中,有26.8%的标本(n=57)处于0级风化阶段(未风化),另有43.2%(n=92)处于1级,21.1%(n=45)处于2级。总体来说,马圈沟遗址第3文化层90%以上的长骨(n=194)处于风化作用早期的三个阶段,其骨骼表面并未发生显著的剥落;处于第3级和第4级的标本(n=15,n=4)非常少,这说明骨骼暴露在地表的时间并不长,几乎没有发生陆上风化。
在这些长骨标本中,骨骼边缘磨圆的状况并不多见,仅在9件标本上观察到了部分磨圆的现象。这9件标本均为不可鉴定的长骨骨干碎片。这些磨圆都是局部的,应该发生在骨骼破裂之后,很可能是短暂的水流冲刷造成的。所以即使有磨圆的存在,也并不影响我们观察这些标本的破碎方式。
在213件长骨标本中,仅有4件可以鉴定到属的标本,其中3件属于马类动物,均为胫骨远端的残片,1件属于鹿类动物。余下的209件标本以其所属动物体型的相对大小分为中型动物与大型动物,39.4%的标本(n=84)属于中型动物,60.6%的标本(n=129)属于大型动物。
5.2长骨破碎方式
本文所关注的马圈沟遗址出土的长骨标本虽然有可以鉴定的标本,但几乎没有完整的长骨。有一些标本保留了长骨近端或远端,但总是存在一个或数个断口。除此之外,其余的长骨标本均为长骨的碎片/块。也就是说,这些长骨受到了一种或多种埋藏作用的影响而破裂。经过观察,仅有15件标本具有非常粗糙的断口,占标本总量的7%。这样的断裂一般是在骨骼比较干燥時发生的。有50件标本的断口质地难以被认定是完全光滑的,而其中,有39件标本的断口角度非常接近90°;这些标本的断裂应该发生在标本的有机质完全流失以后。因此,总共有54件标本的在断裂之前已经有大部分有机质流失,约占标本总量的25.4%。图5展示了马圈沟遗址第3文化层出土长骨残段破裂面的基本情况。
在余下159件新鲜状况下破裂的标本中,有8件标本具有1个断口,12件标本具有2个断口,139件标本具有4个断口。这样,我们从159件新鲜破裂的长骨标本上共获得了588个骨骼破裂面的数据。将这些断口按其所属标本动物体型大小、轮廓及角度性质(锐角或钝角)进行分类统计,并计算其平均值、标准差及95%的置信范围(表1)。
从断口角度的平均值来看,倾斜状断口的锐角平均值显著低于其他断口类型的锐角,表明这些标本不仅是在新鲜状况下破裂的,而且受到的冲击力应该更接近动态冲击。而倾斜状断口测量值为锐角的标本数量也是最多的。大型动物的横向断口平均值最接近90°,这种断口的状态与非新鲜状况下破裂的断口性质更接近。置信范围的大小似乎与样本数量存在一定关系,样本量较大的断口类型,置信范围比较集中,数据的准确性也可能更高;特别是中型动物大于90°的横向断口,标本数量仅为2件,但其置信范围却非常大,并超过了我们预先定义的大于90°的范围。尽管如此,横向断口的置信范围普遍要大于其他类型的断口。
为了与实验结果以及斯瓦特科兰斯(Swartkrans)出土的长骨标本进行比较,我们将马圈沟遗址第3文化层出土长骨破裂面角度的平均值及95%的置信范围,与这二者进行了对比(图6)。
在横向断口上(图6:A,B),马圈沟遗址第3文化层中型动物长骨断裂角度的均值与实验样本中人类石锤砸击的角度最接近。除了大型动物中断裂角大于90度的标本之外,马圈沟遗址第3文化层长骨的置信范围与斯瓦特科兰斯的值非常接近。
在纵向断口上(图6:C,D),马圈沟遗址第3文化层中、大型动物长骨断裂的锐角角度平均值与实验中人类砸击以及斯瓦特科兰斯中、大型动物破裂角的平均值更接近,并且落入了95%的置信范围。而相应的钝角角度并不像斯瓦特科兰斯样本那样有更集中的置信范围,但其平均值似乎与实验中食肉类改造的标本更接近。
无论是实验数据、斯瓦特科兰斯的破裂角度数据,还是马圈沟遗址第3文化层的统计结果,在倾斜状断口上的角度置信范围都很集中(图6:E,F)。其中,中型和大型动物标本锐角的平均值在人类作用的实验数据范围,但钝角的平均值却与食肉动物的实验数据更接近。马圈沟第3文化层倾斜断口角度的置信范围都与斯瓦特科兰斯的非常接近。
6讨论与结语
通过对马圈沟遗址第3文化层出土的大中型哺乳动物长骨破碎方式进行观测,我们初步获得了如下几个方面的认识。
首先,该遗址第3文化层出土的动物长骨残段及碎片大小混杂,虽然以大于10cm的长骨为主,但其他小尺寸的长骨碎片也有很多。这些标本中仅有非常少量的标本上发现被磨蚀的痕迹。加之该层出土了许多人工制品,且保存状况较好,表明该遗址出土遗物大部分应该是由于古人类活动而聚集在此地的。
长骨风化状况的分析结果表明,绝大多数标本只在地表经历了短暂时间的暴露,风化作用对骨骼保存状况没有大的破坏。但遗憾的是,在本文所涉及的标本中,并没有发现骨骼表面保存有自然或人为的痕迹。
第二,马圈沟遗址第3文化层出土了不少草原猛犸象的化石,据发掘者观察,应有牙齿、肋骨、肢骨及脊椎骨等,可能来自于同一个体[15]。在遗址出土的牙齿化石中,我们也发现了属于犀牛的牙齿碎片。除了这些超大型哺乳动物之外,遗址还出土了不少马类、鹿类及羚羊类的标本。遗址出土的长骨残段及碎片中,几乎没有发现可以鉴定到种的标本。我们按以往鉴定的出土动物名单,以动物体型将本次研究的长骨分别归为与羚羊、鹿体型相当的中型动物及与马、牛体型相当的大型动物。结果表明,属于大型动物的标本数量超过60%。需要注意的是,本次研究的标本量并不足以进行个体数量的量化(如统计最小个体数或最小骨骼部位数),只能提供一个相对数量的对比。
第三,通过对这些長骨断口的质地和角度进行观察,我们发现约75%的标本符合新鲜状态下破碎的长骨骨骼特征,即断口质地较光滑或其断口角度为锐角或钝角。哺乳动物的长骨在新鲜状况下破碎一般与其骨髓被取食密切相关,因此马圈沟遗址第3文化层大量新鲜状况下破裂长骨的存在表明,这些哺乳动物因某种原因在本地死亡后,绝大多数被古人类或者食肉动物取食了骨髓。
第四,为了搞清究竟是食肉类还是古人类获取了骨髓,我们测量了新鲜状态下破裂的长骨的断口角度,并与实验数据及针对斯瓦特科兰斯的同类研究结果进行了比较。通过对比统计数据,我们发现马圈沟遗址第3文化层出土的长骨断裂角度与古人类及食肉动物造成的破裂角度均有重合,表明古人类和食肉动物都造成了新鲜骨骼的破裂。但我们测量的数据,其角度测量值的统计范围与实验数据中石锤造成的断口角度的重叠更多。虽然也发现有一些与实验数据中食肉动物造成的断裂角相吻合的数据,但总体上符合食肉动物啃咬角度特征的标本很少。由此我们认为,本文所涉及的大多数标本,都可能与古人类的取食骨髓的行为相关。另一方面,与斯瓦特科兰斯遗址的断口角度范围相比,其置信范围重合的部分更多。虽然在马圈沟遗址第3文化层并未发现确切的骨表面痕迹,但在斯瓦特科兰斯遗址发现的食肉类齿痕出现在了一些新鲜破裂的长骨内侧,表明食肉类取食骨髓的行为应晚于古人类。依据现有的证据我们只能推测,与斯瓦特科兰斯相似,马圈沟第3文化层摄取动物骨髓的主体是古人类,同时也受到食肉类不少的干扰,而古人类的取食行为很可能早于食肉类。虽然这一结论在讨论早更新世古人类狩猎还是食腐问题中的区分效力并不如骨骼表面痕迹理想,但显然提供了一条可以使用的证据,能够评估早期人类行为在考古遗存中的相对贡献,为复原早更新世人类生存状况的证据链提供支持。
第五,研究方法的适用性。早更新世旷野遗址出土动物骨骼的保存状况常常不佳,这限制了对骨骼表面痕迹的观察研究。然而,这种研究的窘迫促使研究者将目光转移到其他类型的数据积累,尝试去推断食肉动物和古人类的相对影响。另一方面,骨骼表面痕迹的分析可能无法自行解释骨骼破碎方式的问题[57],因此也需要使用其他方法来确定骨骼断裂是如何发生的。本文介绍了其中一种重要的方法,就是观测断口的状况和角度并进行分类变量的对比。虽然这个研究体系还存在许多问题,但在数据足够丰富的情况下可以用来区分古人类和食肉动物行为。现阶段,除了进行更丰富、完善的实验考古研究并详尽地报道数据之外,也应尽可能地采集考古遗址出土动物骨骼的破裂信息。显然,破碎骨骼的断口角度甚至断口轮廓及性质这样的基本信息,在以往的研究中也是不受重视的。而本文,也是这一方法在中国早更新世遗址研究中的首次运用。
最近,在标本及数据大量积累的前提下,许多研究者将目光投向了机器学习(machinelearning),该项技术可以分析多变量下的骨骼破损信息,并有效地区分不同营力造成的骨骼破损[58]。当然,这项技术的运用还仅限于实验标本,对考古材料的要求极高。这也促使我们需要进一步开展精细的考古发掘,并不断积累数据。在此基础之上,或许能在早更新世古人类狩猎还是食腐的问题讨论中提出更有力的证据。
