
甘薯PEBP基因家族鉴定以及影响甘薯块根发育候选PEBP基因的鉴定
2024-06-05 08:36黄哲瑞辛曙丽赵添刘永华朱国鹏
热带作物学报 2024年3期
黄哲瑞 辛曙丽 赵添 刘永华 朱国鹏

关键词:甘薯;PEBP 基因家族;生物信息学;组织表达特异性;块根发育
磷脂酰乙醇胺结合蛋白(phosphatidylethanolamine-binding protein, PEBP)具有1 个非常保守的PEBP 结构域,该结构域因能与磷脂酰乙醇胺结合而得名[1]。PEBP 是一种非常保守的蛋白,在生物界中广泛存在,无论是植物、动物还是古细菌中均有存在[2]。植物中的PEBP 最早在拟南芥中发现,可分为3 个亚家族:FT (FLOWERINGLOCUS T)-like、TFL1 (TERMINAL FLOWER 1)-like、MFT (MOTHER OF FT AND TFL1)- like[3]。随着研究的深入,在小麦、棉花等作物中发现PEBP 家族中还存在另一个亚家族,将其命名为PEBP-like[4-5]。
PEBP 家族中成员较多,功能多样[3]。在拟南芥中存在6 个PEBP 家族成员,其中2 个属于FT-like(AtFT、AtTSF)、3 个属于TFL1-like(AtTFL1、AtBFT、AtATC),1 个属于MFT-like(AtMFT)。拟南芥AtFT 基因与AtTFL1 基因在开花诱导方面显示出显著的拮抗功能[6]。在长日照条件下,拟南芥AtFT 的上游基因CONSTANS(CO)表达上升并与光敏色素作用因子PIF4 相互作用于AtFT的启动子区域,从而激活了叶片中FT 的表达[7],FT 蛋白在叶片中产生后,通过维管束运输到顶端分生组织的细胞质中[8],与bZIP 转录因子FD 蛋白结合形成FT/FD 蛋白复合物,从而诱导下游成花相关基因的表达,促进拟南芥开花[9]。在短日照条件下,AtTFL1 基因表达上升,而FD 既能和FT 蛋白结合,也能和TFL1 蛋白结合,因此TFL1蛋白通过与FT 蛋白竞争和FD 的结合,从而抑制拟南芥开花[10]。拟南芥FT-like 的另一成员TSF会协助FT 对FD的竞争。而TFL1-like 成员AtBFT、AtATC 也会协助TFL1 对FD 的竞争。因此拟南芥开花不仅取决于FT 和TFL1,还与FT-like、TFL1-like 其他成员的表达水平有关[11]。过表达研究表明,AtMFT 有促进拟南芥开花的作用,AtMFT 基因还可以通过调控ABA 和GA信号来调节种子的萌发[12-14]。与拟南芥FT-like 类似,水稻FT-like的RICE FLOWERING LOCUS T1(RFT1)基因在长日照时具有诱导开花的作用[15]。
除了调控植物开花和种子萌发外,PEBP 还可调控植物地下部储藏器官如鳞茎、块茎等的生长发育。如洋葱(Allium cepa L)FT-like 中的AcFT1和AcFT4 对洋葱磷茎的发育有拮抗作用。在長日照条件下,AcFT1 基因表达量升高,能够促进洋葱产生鳞茎,而在短日照条件下,AcFT4 表达量增加,抑制AcFT1 基因的转录,使洋葱提早开花,同时鳞茎的形成受到抑制[16]。在马铃薯中,过表达FT 同源基因StSP6A 会抑制蔗糖转运蛋白StSWEET11 的活性,使蔗糖的卸载途径从质外体途径转变为糖转运效率更高的共质体途径,从而提高蔗糖向块茎的转运和马铃薯块茎的形成[17]。而另一个FT 同源基因StSP5G 可以通过抑制StSP6A 的表达阻碍马铃薯块茎的形成[18]。马铃薯StCEN1 属于TFL1-like,与脱落酸和细胞分裂素的信号传导有关[19],StCEN1 的过表达也可以抑制马铃薯块茎的形成[20]。水稻FT 同源基因Hd3a的产物在短日照条件下会与细胞质的14-3-3 蛋白结合形成二级复合体,然后该复合体进入细胞核中与bZIP 转录因子FD 蛋白结合形成三级复合体FAC,诱导水稻提早开花[21]。此外,水稻Hd3a基因在马铃薯中过表达能够促进马铃薯块茎的形成[18]。还有研究表明PEBP 基因也在番茄[22]、大麦[23-24]、毛竹[25]、木薯[26]、苹果[27-28]、郁金香[29]等多种作物的生长发育中发挥着重要的调控作用。
甘薯(Ipomoea batatas)别名甜薯、地瓜、番薯,为旋花科草本植物。甘薯起源于中南美洲,在16 世纪末才传入中国[30]。由于甘薯环境适应能力强、产量较高且易于栽培管理,因此很快就在中国广泛种植[31]。甘薯块根、茎、叶均可食用,富含淀粉、可溶性糖、维生素C、维生素E、胡萝卜素、花青素等人体所需物质[32]。此外,甘薯还具有重要的经济价值,可作为饲料、淀粉加工原料和工业原材料[33]。甘薯已经成为全球第七大农作物,我国甘薯栽培面积和产量分别占全球的42%和68%,稳居世界第一(FAO,2020)。目前我国甘薯单产已处于平台期,近20 年来(1999—2020 年)一直徘徊在22 t/hm2 左右,没有进一步提升(FAO, 2020)。虽然和大多数发展中国家相比我国甘薯单产较高,但和发达国家相比,我国甘薯单产仍有一定差距,例如美国和澳大利亚平均单产分别高达25.4 t/hm2 和39 t/hm2(FAO,2020)。如何进一步提升甘薯产量是我国甘薯产业健康、可持续发展所亟需解决的重要问题之一。
与马铃薯块茎和洋葱鳞茎等类似,甘薯的主要食用器官块根也属于地下部储藏器官,因此对其PEBP 家族成员进行研究将有助于进一步提高甘薯产量。虽然已有研究报道甘薯PEBP 基因家族,但该研究仅从甘薯基因组中鉴定出13 个PEBP 家族成员,不包含PEBP-like 成员[34]。截至目前,尚未有报道系统鉴定和阐明与甘薯块根发育相关的候选PEBP 家族成员。鉴于此,本研究首先利用生物信息学方法对甘薯基因组中PEBP家族成员的数量和种类进行系统鉴定,然后通过生物信息学分析PEBP 基因表达的组织特异性(幼叶、成熟叶、茎、纤维根、柴根、薯皮、块根、花),初步鉴定出可能调控甘薯块根发育的候选PEBP 家族成员,最后通过对候选PEBP 基因在不同发育时期块根中表达水平的动态变化及其与SWEET 基因表达水平之间的相关性分析,进一步确认其在调控甘薯块根发育中的重要作用。本研究将为后续深入研究甘薯PEBP 基因家族的功能奠定基础,同时为进一步提高我国甘薯产量提供理论支撑。
1 材料与方法
1.1 材料
选取甘薯品种高系14 为实验材料,于海南大学(20°2?39.73?N,110°18?26.93?E)进行露地栽培,以史丹利复合肥(N∶P∶K=15∶15∶15)450 kg/hm2 和羊粪有机肥12 000 kg/hm2 作为基肥,在种植前先整地起垄,按照40 cm 间隔起垄,垄宽80 cm,每垄只种1 行甘薯,株距为25 cm。种植时间为2021 年9 月20 号至2021 年12 月3号,开花后(74 d)取样。为了研究不同组织的基因表达,选择高系14 的幼叶(茎尖周围的未展开叶)、成熟叶(从茎尖向下数的第5 片完全展开叶片)、茎、花(不包括花柄和花萼)、纤维根(白色细长呈纤维状的根)、柴根(直径0.2~2.0 cm 粗细均匀的红色根)、块根(直径约4 cm 的膨大根)的薯皮和薯肉8 个部位(图1),每个部位取0.2 g,3 个生物学重复。
2022 年1—5 月在海南大学海甸校区温室大棚盆栽种植高系14。花盆规格为:外径29.6 cm,内径25.4 cm,高19.7 cm,底部直径17.8 cm。基肥用羊粪有机肥和史丹利复合肥(N∶P∶K=15∶15∶15),田园土和羊粪有机肥按3∶1 的体积比加上每盆0.8 g 的史丹利复合肥(N∶P∶K=15∶15∶15)混合均匀,装入花盆。剪取25 cm 长带茎尖的甘薯茎段,以直插法进行扦插,深度为10 cm。在甘薯扦插后的30、60、90、120 d 取样,取0.2 g块根薯肉提取总RNA,每个取样时期均含4 个生物学重复。取样后液氮处理,置于–80 ℃超低温冰箱保存备用。
1.2 方法
1.2.1 甘薯PEBP 基因家族成员鉴定 从甘薯基因组数据库(https://ipomoea-genome.org/)下载甘薯的全基因组文件和全基因组注释文件。从NCBI 数据库(https://www.ncbi.nlm.nih.gov/)下载拟南芥PEBP 家族的蛋白序列[26]。从Phytozomev13 数据库(https://phytozome-next.jgi.doe.gov/)下载木薯PEBP 家族的蛋白序列[26]。从马铃薯的Spud DB 数据库(http://solanaceae.plantbiology.msu.edu)下载马铃薯PEBP 家族的蛋白序列[35]。
为了鉴定甘薯PEBP 候选基因,使用2 种方法搜索甘薯的蛋白质序列。第一种方法:准备拟南芥(6 个)、木薯(10 个)、马铃薯(15 个)的PEBP基因家族蛋白序列,通过TBtools[36](https://github.com/CJ-Chen/TBtools)在線软件将甘薯全蛋白序列进行本地Blastp 比对,参数阈值设置为E-value≤1e–5,其他参数为默认值。第二种方法:通过查找PEBP 家族蛋白的隐马尔可夫模型(HMM)的ID(PF01161)、下载甘薯全蛋白序列和Pfam-A.hmm 文件(ftp://ftp.ebi.ac.uk/pub/databasesfam/current_release/Pfam-A.hmm.gz)。将ID(PF01161)、甘薯全蛋白序列、Pfam-A.hmm 文件用TBtools 软件检索出可能的甘薯PEBP 基因。
将2 种方法得到的候选基因,提交给Pfam 数据库[37](http://pfam.xfam.org/)、NCBI 网站[38]( https://www.ncbi.nlm.nih.gov/Structu-re/cdd/cdd.html ) 和SMART 网站[39] ( http://smart.embl-Heidelberg.de/smart/set_mode.cgi?NORMAL=1)进一步确认是否含有保守的PEBP 结构域。
1.2.2 甘薯PEBP 家族成员染色体定位 使用TBtools 软件分析甘薯PEBP 家族成员基因的注释信息。
1.2.3 甘薯PEBP 家族蛋白系统发育分析 为了对甘薯的PEBP家族基因进行分类系统发育分析,使用MEGA-X(https://www.megasoftware.net/)在线软件的邻接法(Neighbor-joining method)将拟南芥、木薯、马铃薯、甘薯的PEBP 蛋白的全长氨基酸序列构建系统发育树,其中参数设定的自展值为1000,其他参数默认。使用Evolview( https://www.evolgenius.info/evolview/)在线软件对系统发育树进行美化。此外,按上述方法用MEGA-X 软件将甘薯PEBP 基因与其他物种已知功能的PEBP 基因(马铃薯的StSP6A、StSP5G、StCEN1;洋葱的AcFT1 和AcFT4;水稻的Hd3a)构建系统发育树,以此推断相关甘薯PEBP 基因的功能。
1.2.4 甘薯PEBP 家族成员理化性质分析 使用ExPASy( http://web.expasy.org/protparam/)在线软件预测PEBP 蛋白的理化性质(等电点、分子量、氨基酸数目)。使用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)在线软件进行亚细胞定位预测。
1.2.5 甘薯PEBP 基因家族成员表达分析 使用Premier 5.0 软件设计实时定量PCR 所需引物,并在NCBI 网站检验引物特异性。其中IbFT1、IbFT3与IbFT2 的CDS 序列相似度高且前2 个基因的CDS 序列比IbFT2 短,IbTFL1 与IbTFL2 相比也是如此,因此IbFT1、IbFT3、IbTFL1 无法设计特异性引物,后续不再对IbFT1、IbFT3、IbTFL1进行基因表达研究。具体引物信息如表1 所示,其中,5 个SWEET 基因(IbSWEET4、IbSWEET7、IbSWEET11、IbSWEET16 和IbSWEET19)的引物信息参照张文杰[40]的研究。
利用CWBIO 公司的RNA 提取试剂盒[OmniPlantRNA Kit(Dnase I)]提取总RNA,并通过琼脂糖凝胶电泳检查其完整性;逆转录使用诺唯赞HiScript III 1st Strand cDNA Synthesis Kit 逆转录试剂盒;使用诺唯赞ChamQ Universal SYBRqPCR Master Mix 试剂盒,以甘薯的β-Actin 基因为内参,用德国耶拿qTOWER3G 定量PCR 仪进行基因表达水平测定。具体反应条件如下:95 ℃预变性5 min;95 ℃变性5 s,58 ℃退火30 s,72 ℃延伸30 s,40 个循环。采用2–ΔΔCT 法计算基因的相对表达量。
1.3 数据处理
利用Excel 2020 软件和Graphpad prism 8.0 软件处理甘薯PEBP 基因相对表达数据,使用SPSS22.0 软件对数据进行单因素方差分析(ANOVA),多重比较采用Duncans 法分析。
2 结果与分析
2.1 甘薯PEBP 基因家族成员的鉴定
通过本地Blastp 比对和隐马尔可夫模型(HMM)分析,从甘薯基因组中找到16 个PEBP候选基因,然后将其提交到Pfam 数据库、NCBI网站和SMART 网站验证,发现编号为g46241.t1的蛋白无PEBP 结构域,将其剔除。因此,从甘薯基因组中总共鉴定出15 个PEBP 家族成员。用MEGA-X 软件将鉴定出的15 个甘薯PEBP 家族成员与已经公布的拟南芥、木薯、马铃薯的PEBP家族成员用邻接法构建系统发育树(图2)。结果显示,15 个甘薯PEBP 家族成员被分为4 个亚家族,其中包含5 个FT-like 成员、6 个TFL1-like成员、2 个MFT-like 成员和2 个PEBP-like 成员。根据每个亚家族成员在染色体上位置的先后顺序进行命名,分别命名为IbFT1~5、IbTFL1~6、IbMFT1~2、IbPEBP1~2(表2)。值得注意的是,现有研究仅在甘薯基因组中鉴定出13 个PEBP 家族成员,包括5 个FT-like 成员、6 个TFL1-like成员、2 个MFT-like 成员,但缺少2 个PEBP-like成员[34]。
2.2 甘薯PEBP 基因家族成员的功能预测
为了鉴定出可能与甘薯块根发育相关的PEBP家族成员,将其他农作物中已知会促进或抑制地下部储藏器官膨大的PEBP 基因(包括马铃薯的StSP6A、StSP5G 和StCEN1 基因[17-20],洋葱的AcFT1 和AcFT4 基因[16],以及水稻的Hd3a 基因[18])和甘薯的15 个PEBP 家族成员进行聚类分析(图3),通过亲缘关系的远近预测可能和块根发育相关的甘薯PEBP 基因。结果发现,在15 个甘薯PEBP 基因中,IbFT5 与洋葱基因AcFT1 和水稻基因Hd3a 的亲缘关系较近,而这2 个基因可分别促进洋葱鳞茎[16]和马铃薯块茎的发育[18]。因此,推测IbFT5 可能和甘薯块根发育密切相关。此外,与其他甘薯PEBP 基因相比,IbTFL3 和马铃薯中抑制块茎膨大的TFL1-like 基因StCEN1(StPEBP5)[20, 35]的亲缘关系较近,因此推测IbTFL3 可能会抑制甘薯块根的发育。
2.3 甘薯PEBP 基因的组织特异性表达分析
为进一步揭示甘薯PEBP 家族成员的功能,对甘薯PEBP 基因在不同组织(幼叶、成熟叶、茎、纤维根、柴根、薯皮、薯肉、花)中的表达特异性进行qRT-PCR 分析(图4)。结果表明,IbTFL5、IbMFT1 和IbPEBP2 在所有组织中均具有较高的表达水平,其可能在不同组织的发育中均发挥着重要作用,而与块根的膨大无必然联系。
IbFT2 只在幼叶和成熟叶中表达,为叶片特异表达基因。IbMFT2 在花中的表达水平远高于其他组织,为花特异性表达基因。IbFT4 仅在花和成熟叶中有表达,且在花中的表达水平显著高于成熟叶,为花和叶片特异表达基因。IbTFL3、IbTFL4和IbTFL6 不仅在花中高表达,也在块根的薯皮和薯肉中高表达。根系特异表达的基因为IbFT5、IbTFL2 和IbPEBP1,其中IbFT5 仅在非膨大根(纤维根和柴根)中高表达,而IbTFL2 和IbPEBP1不仅在非膨大根中高表达,也在膨大块根的薯皮和薯肉中高表达。上述结果表明,甘薯PEBP 基因家族成员的表达具有明显的组织特异性,推测甘薯PEBP 家族成员存在显著的功能分化,其在不同组织中发挥着不同的功能。
由图4 可知,在根系(膨大根和非膨大根)中特异表达基因为IbFT5、IbTFL2 和IbPEBP1。
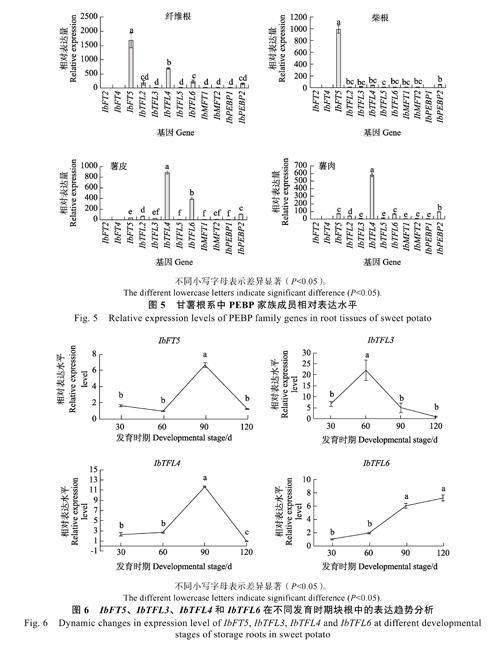
此外,IbTFL3、IbTFL4 和IbTFL6 也在根系中特別是薯皮和薯肉中高表达。推测上述6 个甘薯PEBP 基因可能与甘薯块根发育密切相关。为缩小PEBP 基因成员的选择范围,本课题组进一步对特定根系组织(纤维根、柴根、薯皮和薯肉)中不同PEBP 家族成员的相对表达水平进行测定(图5)。结果发现,在纤维根中,IbFT5 和IbTFL4为高表达基因,且IbFT5 的表达水平显著高于IbTFL4。在柴根中,IbFT5 为高表达基因,其余基因的表达水平极低。在膨大块根薯皮中,IbTFL4、IbTFL6 为高表达基因,其余基因低表达或不表达。在膨大块根薯肉中,IbTFL4 为高表达,其余基因为低表达或不表达。
综上所述,在根系中高表达的6 个PEBP 基因中,IbFT5、IbTFL4 和IbTFL6 不仅在根系中的表达水平高于在其他组织,而且也高于根系中其他PEBP 家族成员。因此推测这3 个PEBP 基因在块根膨大中发挥着重要作用。
2.4 甘薯PEBP 基因在不同发育时期块根中的表达及其与SWEET 基因表达的相关性
聚类分析揭示IbFT5 可能会促进甘薯块根的发育,而IbTFL3 则可能会抑制甘薯块根的发育(图3);此外,组织表达特异性分析则表明IbFT5、IbTFL4 和IbTFL6 与块根发育密切相关(图4、图5)。因此,本课题组进一步对上述4 个基因(IbFT5、IbTFL3、IbTFL4 和IbTFL6)在不同发育时期块根(定植后30、60、90、120 d)中的表达水平动态变化进行测定,观察上述基因表达水平和块根膨大之间的相关性。
结果表明,在块根发育的前期(30~60 d),IbFT5、IbTFL4 的表达水平无显著变化,而在块根发育中期(60~90 d),这2 个基因的表达水平快速大幅增加,但在块根发育后期(90~120 d),其表达水平均快速下降(图6)。与IbFT5、IbTFL4不同,IbTFL6 的表达水平在整个块根发育时期一直呈上升的趋势,在整个发育时期呈‘S形曲线,即在块根发育前期(30~60 d)和后期(90~120 d),其表达水平上升速度较慢,而在发育中期则上升速度较快。
在上述4 个基因(IbFT5、IbTFL3、IbTFL4和IbTFL6)中,只有IbTFL3 基因根据聚类分析(图3)被认为可能会抑制甘薯块根的发育。IbTFL3 基因表达水平的动态分析表明,其表达水平在块根膨大前期(30~60 d)呈上升趋势,而在块根发育的中后期(60~120d)其表达水平快速下降。总的来讲,伴随着IbFT5、IbTFL4 和IbTFL6表达水平的上升,IbTFL3 的表达水平则呈现下降趋势,表明这2 类基因之间可能存在拮抗作用,前者可能促进块根发育,而后者可能抑制块根发育。
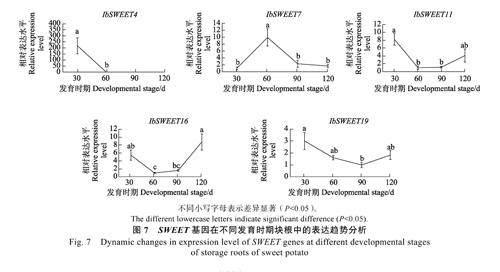
SWEET 蛋白为蔗糖转运蛋白,其主要功能是将蔗糖和己糖(葡萄糖和果糖)由细胞内向外转运,其表达量低往往代表糖分的转运途径以高效的共质体途径为主,而低效的质外体途径则属于次要途径。例如,在马铃薯中,过表达PEBP基因StSP6A 会抑制蔗糖转运蛋白StSWEET11 的表达,从而促进蔗糖由叶片通过共质体途径向块茎的高效转运,最终促进马铃薯块茎的形成[17]。研究表明,在甘薯块根中表达的SWEET 基因有5 个,分别为IbSWEET4、IbSWEET7、IbSWEET11、IbSWEET16 和IbSWEET19[40]。本课题组对不同发育时期块根中这5 个SWEET 基因的表达水平进行测定。结果表明,随着块根膨大和块根发育相关PEBP 基因(IbFT5、IbTFL4 和IbTFL6)表达水平的上升,IbSWEET4、IbSWEET11、IbSWEET16和IbSWEET19 的表达均呈现不断下降的趋势,在定植后60 d(IbSWEET19 是在定植后90 d)达到最低值,其表达水平在块根膨大后期(90~120 d)又呈现回升的趋势,而IbSWEET7 基因的表达在块根膨大前期(30~60 d)呈现上升的趋势(图7)。根据上述结果,可推测甘薯PEBP 基因很可能也是通过抑制SWEET 基因的表达来促进块根发育。
3 讨论
PEBP 家族在植物中广泛存在,近年来研究人员已经在多种植物中发现了PEBP 基因家族。PEBP 基因在植物生长发育中发挥着重要作用,不仅调控拟南芥和水稻等植物的开花[6, 12, 15, 21],还参与调控地下部储藏器官的形成如马铃薯块茎[17-20]和洋葱鳞茎等[16]。与马铃薯块茎和洋葱鳞茎等类似,甘薯的主要食用器官块根也属于地下部储藏器官,因此对其PEBP 家族成员进行研究将有助于进一步提高其块根产量。虽然近期有研究报道了甘薯PEBP 家族的生物信息学分析结果,但该研究从甘薯基因组中仅鉴定出13 个PEBP 家族成员,且只包含FT-like、TFL1-like 和MFT-like,并不包含PEBP-like[34]。此外,目前尚未有报道系统研究并筛选可能和甘薯块根发育相关的候选PEBP家族成员。
为了解决上述存在的问题,本研究利用生物信息学方法从甘薯基因组中共鉴定出16 个PEBP候选基因,然后将其提交到Pfam 数据库、NCBI网站和SMART 网站验证,发现除了编号g46241.t1的成员,其他15 个甘薯PEBP 成员和前人研究的拟南芥、木薯、马铃薯结果相同,均存在着保守的PEBP 结构域。将g46241.t1 剔除,得到15 个甘薯PEBP 家族成员。在植物进化过程中,许多植物都经历过1次或多次多倍体化从而导致基因家族成员扩张,拟南芥和木薯为二倍体,甘薯为六倍体,且拟南芥PEBP 家族成员(6 个)和木薯PEBP 家族成员(10 个)与甘薯成员相比,甘薯PEBP 家族成员数量更多,说明甘薯在进化过程中随着染色体倍数的增加,导致了甘薯PEBP 家族的扩张。此外,系统发育树结果显示与拟南芥和木薯相比,甘薯除了有FT-like、TFL1-like 和MFT-like 外,还多了PEBP-like,而在他人研究的马铃薯、小麦、棉花等多个物种中也鉴定出PEBP-like[4-5, 35]。甘薯有5 个FT-like 成员、6 个TFL1-like 成员、2 个MFT-like 成员和2 个PEBP-like成员,比较发现不管是在拟南芥、木薯还是馬铃薯中的TFL1-like 成员数量在亚家族中都是最多的,推测在进化过程中TFL1-like 受环境选择发生的基因复制和变异的频率更高。和已有的研究相比[34],本研究从甘薯基因组中多鉴定出2 个PEBP基因,且这2 个基因均为PEBP-like 成员。
在植物中PEBP 家族基因能够影响贮藏器官的形成。在马铃薯中,块茎为马铃薯的贮藏器官,过表达StSP6A 能够促进块茎的形成,而过表达StSP5G 和StCEN1 抑制了块茎的形成[17-20]。此外过表达水稻的Hd3a 基因也能够促进马铃薯块茎的形成[18]。在洋葱中,鳞茎为洋葱的贮藏器官,过表达AcFT1 能够促进鳞茎的形成,而过表达AcFT4 能够抑制鳞茎的形成[16]。甘薯、马铃薯、洋葱、水稻聚类分析发现,IbFT5 与洋葱基因AcFT1和水稻基因Hd3a 的亲缘关系较近,而这2 个基因可分别促进洋葱鳞茎[16]和马铃薯块茎的发育[18]。推测IbFT5 可能也会促进甘薯块根的发育。此外,和其他甘薯PEBP 基因相比,IbTFL3 和马铃薯StPEBP5(StCEN1)[20, 35]的亲缘关系较近,推测IbTFL3 可能会抑制甘薯块根的发育。
植物PEBP 基因家族具有组织特异性。木薯中MeFT1 和MeFT2 在成熟叶中高表达,推测成熟叶是产生花诱导信号的部位。MeMFT1 在幼叶和花芽的生长组织中高表达,而MeMFT2 在须根和茎中高表达[26]。本研究中,甘薯根系(膨大根和非膨大根)特异表达的基因为IbFT5、IbTFL2和IbPEBP1。此外,IbTFL3、IbTFL4 和IbTFL6除了在花中高表达,也在根系中特别是薯皮和薯肉中高表达,但在其他组织中为低表达或不表达。推测上述6 个PEBP 基因和甘薯块根发育密切相关。为进一步研究发现,上述6 个基因在甘薯根系中(纤维根、柴根和膨大块根薯皮和薯肉)中高表达的PEBP 基因只有IbFT5、IbTFL4、IbTFL6等3 個基因,其余基因为低表达。因此,在根系中高表达的6 个PEBP基因中,只有IbFT5、IbTFL4和IbTFL6 不仅在根系中的表达水平高于在其他组织中的表达水平,而且和根系中表达的其他PEBP 家族成员相比,这3 个基因的表达水平也是最高的。推测这3 个PEBP 基因(IbFT5、IbTFL4和IbTFL6)在根系膨大中发挥着重要的作用。
综合上述2 个方面的结果,推测IbFT5、IbTFL4 和IbTFL6 可能会促进块根的发育,而IbTFL3 则可能会抑制块根的发育。已有的研究表明,甘薯块根定植后30 d 开始逐渐膨大,定植后60~90 d 进入快速膨大期,随后膨大速度逐渐减慢[41]。对上述4 个基因在不同发育时期块根(定植后30、60、90、120 d)中的表达水平进行的测定结果表明,在块根发育的前期(30~60 d),IbFT5、IbTFL4 的表达水平无显著变化,而在块根快速膨大期(60~90 d),这2 个基因的表达水平快速大幅增加。与IbFT5、IbTFL4 不同,IbTFL6的表达水平在整个块根发育时期一直呈现上升的趋势,特别在块根快速膨大期(60~90 d)其上升速度较快。这与马铃薯中的StSP6A 表达相似。马铃薯的StSP6A 也在贮藏器官发育时期呈现上升趋势,当地下匍匐茎弯钩角度为150°时,StSP6A的表达量急剧增加,发育成小薯和大薯时,表达量也还保持在较高水平[42]。推测IbFT5、IbTFL4和IbTFL6 这3 个基因可能促进甘薯块根的发育,根据聚类分析推测IbTFL3 基因可能会抑制甘薯块根的发育。IbTFL3 基因表达水平的动态变化分析表明,其表达水平在块根快速膨大期(60~90 d)开始快速下降,表明其很可能对块根膨大起抑制作用。此外,IbTFL3 在柴根中表达水平显著高于其他根系(纤维根、块根的薯肉和薯皮),而柴根是生长停滞的块根,这进一步证明IbTFL3 很可能会抑制块根的膨大。
SWEET 蛋白为蔗糖转运蛋白,其主要功能是将蔗糖和己糖(葡萄糖和果糖)由细胞内向外转运,从而促进糖分经由低效率的质外体途径转运,而不是经由高效率的共质体途径转运。前人研究表明,在甘薯块根中高表达SWEET 基因有5 个,分别为IbSWEET4、IbSWEET7、IbSWEET11 、IbSWEET16 和IbSWEET19[40]。不同发育时期的块根中,随着块根膨大和PEBP 基因(IbFT5、IbTFL4和IbTFL6)表达水平的上升,4 个SWEET 基因( IbSWEET4 、IbSWEET11 、IbSWEET16 和IbSWEET19)的表达均呈现不断下降的趋势,在定植后60 d(IbSWEET19 是在定植后90 d)达到最低值。推测甘薯PEBP 基因很可能是通过抑制SWEET 基因的表达来促进块根发育。在马铃薯的研究中,过表达PEBP 基因,StSP6A 会抑制蔗糖转运蛋白StSWEET11 的表达,从而促进蔗糖通过共质体途径向块茎的高效转运,最终促进马铃薯块茎的形成[18]。此外,甘薯块根中SWEET 基因表达水平的下降也可以解释为何甘薯块根在定植后40 d 块根开始膨大时其蔗糖转运途径由质外体途径转变为共质体途径[41]。
本研究不仅确定了甘薯基因组中PEBP 基因家族成员的数量和种类,而且通过生物信息学分析和PEBP 基因表达的组织特异性分析初步鉴定出可能调控甘薯块根发育的候选PEBP家族成员。此外,通过对相关PEBP 基因在不同发育时期块根中表达水平的动态变化及其与SWEET 基因表达水平之间的相关性分析,初步推断PEBP 可能通过调控糖分转运来影响甘薯块根发育。本研究将为后续深入研究甘薯PEBP 基因家族的功能提供理论依据。
猜你喜欢
保鲜与加工(2021年1期)2021-02-06
今日农业(2019年16期)2019-01-03
中国校外教育(下旬)(2016年11期)2016-12-27
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
电脑知识与技术(2016年27期)2016-12-15
中国科技纵横(2016年17期)2016-11-30
科技视界(2016年23期)2016-11-04
西南农业学报(2016年5期)2016-05-17
当代畜禽养殖业(2014年7期)2014-02-27
