
板栗胚胎及胚乳发育过程的形态解剖观察
2020-05-15 00:17杜兵帅曹庆芹房克凤
北京农学院学报 2020年2期
杜兵帅,张 卿,曹庆芹,邢 宇,秦 岭,房克凤*
(北京农学院a. 园林学院;b. 植物科学技术学院;c. 农业应用新技术北京市重点实验室;d. 北京农学院林木分子设计育种高精尖创新中心,北京102206)
在被子植物的有性生殖过程中,花粉管释放的两个精细胞分别与胚囊中的卵细胞和中央细胞结合进行双受精作用,进而形成合子和初生胚乳细胞[1]。合子遵循定向且高度有序的分裂模式,在基因有序的选择性表达调控下,经过早期原胚,球形胚,心形胚,鱼雷形胚和子叶形胚,最终发育为成熟的胚胎[2]。胚乳作为双受精过程中精子与极核结合形成的滋养组织,主要分为沼生目型胚乳、核型胚乳和细胞型胚乳[3]。沼生目型胚乳多见于单子叶植物中;核型胚乳先进行核分裂,形成多个游离核,与周围的细胞质充满整个胚囊后形成细胞壁,再完成细胞化,进而为胚胎的发育提供营养;而细胞型胚乳则是初生胚乳细胞进行细胞分裂,有规则的形成细胞壁,不存在游离核时期[4]。胚胎发育至种子的过程与碳水化合物的代谢密切相关,胚乳又是胚胎生长发育所需碳水化合物的主要供体[5-8],所以两者的发育紧密相关。
板栗(CastaneamollissimaBlume.)为雌雄同株经济林树种,雄花为直立葇荑花序,雌花一般单独或数朵生于总苞内[9]。板栗雌苞内通常有3枚子房,每子房有6~9心室,每心室有两枚倒生胚珠,共12~18枚胚珠,胚珠双珠被,且靠子房一侧生长较快[10]。在这12~18枚胚珠中一般仅有1个胚珠可以发育为种子,其余的胚珠后期均败育。在可以完成受精作用的正常胚珠内,具备蓼型胚囊的典型结构[11]。花粉落至柱头上,萌发长出花粉管进入花柱后,精细胞与胚囊中卵细胞结合完成受精作用形成合子,开始胚胎基本组织的形成和形态的改变[12],而这些变化离不开胚乳的支撑,因此在这个过程中同时伴随着胚乳的发育与消耗[13]。已有研究表明成熟的板栗种子不具胚乳,食用部分为膨大的子叶[14]。因此胚乳的发育相对来说十分短暂,但过程多样且复杂。目前国内关于板栗生殖方面的研究虽然很多,但大多停留在形态描述、胚胎发生等方面,在胚胎发育过程中的4个关键时期(球形胚、心形胚、鱼雷形胚、子叶形胚),胚乳以怎样的形态存在和变化却并不十分清楚。
同时,板栗作为世界著名树种,种子具有丰富的营养价值,淀粉、蛋白质含量丰富,有“木本粮食”和“干果之王”的美誉[15],并且在山区绿化、促进山区经济发展方面起很大作用。就中国板栗栽培形势而言,种植面积广泛,但产量和效益相对较低,这与板栗种子在发育过程中出现的败育、空苞有很大的关系[16, 17]。因此本试验对板栗种子成熟过程中胚胎及胚乳的变化进行研究,以期进一步掌握板栗的生殖发育特征,并为探明板栗结实率低的内在原因提供解剖学依据。
1 材料与方法
1.1 试验材料
材料采自北京市怀柔板栗试验站,从7月1日至7月25日,每隔5 d采集雌苞,采回后将胚珠从子房中剥出。
1.2 试验方法
1.2.1 板栗胚珠石蜡切片制作 将胚珠标注好日期,放至FAA固定液中固定24 h,用梯度酒精(50%,70%,80%,95%,100%)和二甲苯(50%,100%)进行脱水,在最后1次处理后加入切碎的石蜡碎屑至饱和,在42 ℃恒温箱中过夜;次日在60 ℃恒温箱中将原液换为纯蜡(4.5 h换纯蜡2次),将浸好蜡的胚珠与蜡液一起倒至叠好的纸盒中,用镊子迅速调整胚珠的位置,使其冷却;次日用石蜡切片机对包埋好的胚珠进行纵切,切片厚度为5~7 μm;在48 ℃恒温箱中烘片1晚,次日用梯度二甲苯和梯度酒精进行脱蜡。
1.2.2 数据测量及分析 将制好的切片用BX51显微镜(Olympus)进行观察,用软件DP2-BSW对胚乳细胞径值进行测量,并用SPSS软件对数据进行记录分析,计算平均值及标准差,最后将结果输入Prism软件,绘制成波形图。
2 结果与分析
2.1 板栗胚胎发育过程形态解剖观察
‘怀黄’板栗的盛花期通常在六月中旬,从盛花期到子叶胚的形成需要35~45 d,到果实完全成熟需要90 d左右。当原胚形成后,在之后的发育过程中胚胎的形态及组成结构都发生了明显的改变:盛花期后15~20 d,胚胎的基本组织(原表皮层、维管组织等)开始形成,胚胎发育到达球形期(图1-a,b;图a为早球形胚,图b为晚球形胚);盛花期后20~25 d,胚体细胞快速分裂和分化,子叶原基开始形成,胚胎发育到达心形期(图1-c,d);盛花期后25~30 d,子叶增大,胚胎发育至鱼雷形(图1-e);盛花期后35 d左右,胚胎发育至子叶形(图1-f,g),而后,子叶形胚细胞开始快速扩张,子叶形胚膨大增厚,逐渐充满整个胚囊(图1-h)。
2.2 畸形胚胎
在试验过程中发现存在胚胎形态畸形的现象,2片子叶在发育过程中连接在了一起,形成环形,胚乳细胞未见明显横化趋势(图2-a,b),同时,还存在胚胎数量畸形的现象,在一枚胚珠内存在2个子叶胚,中间有环形珠被相隔(图2-c)。
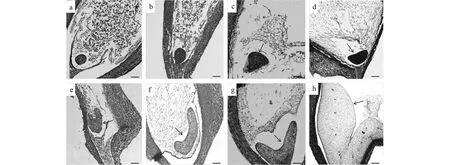
a.早球形胚;b. 晚球形胚;c, d. 心形胚;e. 鱼雷形胚;f, g, h. 子叶形胚;箭头指向胚胎,Bar=100 μm图1 板栗胚胎发育过程a, b. Globular embryos;c, d. Heart shaped embryos;e. Torpedo shaped embryos;f, g, h. Cotyledonary embryos;The arrow points to embryos, Bar=100 μmFig.1 Process of chestnut embryonic development
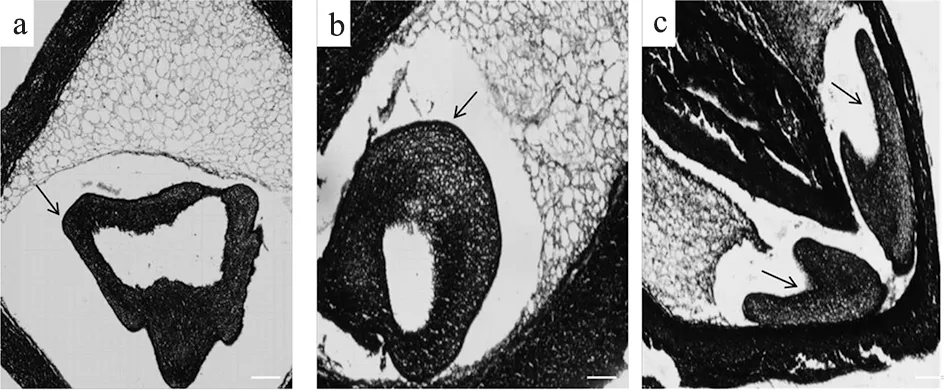
a, b.胚胎形状畸形;c. 胚胎数量畸形;箭头指向胚胎,Bar=100 μm图2 畸形胚胎 a, b. Abnormal embryos;c. Two embryos in one ovule;The arrow points to embryos, Bar=100 μmFig.2 Malformed embryos
2.3 板栗胚乳发育过程的形态解剖观察
板栗胚乳属于核型胚乳,在发育过程中存在游离核细胞化这一转变过程。盛花期后15~20 d,胚胎刚发育至早球形胚时,胚乳仍处于游离核时期,无细胞壁包被,同周围的细胞质共同构成胚乳囊(图3-a)。随着胚胎的发育,游离核开始从珠孔端进行细胞化,形成胚乳细胞(图3-b),合点端方向仍为游离核状态;盛花期后20~30 d,胚胎从球形向心形和鱼雷形发育的过程中,游离核胚乳的细胞化沿胚囊壁向合点端进行,直至胚乳完全细胞化,并且充满整个胚囊。但同时由于胚体本身的发育膨大,会逐渐水解一部分胚乳,形成胚乳腔(图3-c,d);盛花期后35 d左右,胚胎发育至子叶形,胚体会从胚乳细胞中大量吸收营养,进而导致胚乳细胞横向扩张,部分已出现解体、细胞不完整的现象(图3-e);伴随着子叶胚的膨大,胚乳被继续消耗、水解,细胞壁不完整,细胞质减少,进入脱水期,直至几乎被完全吸收(图3-f)。
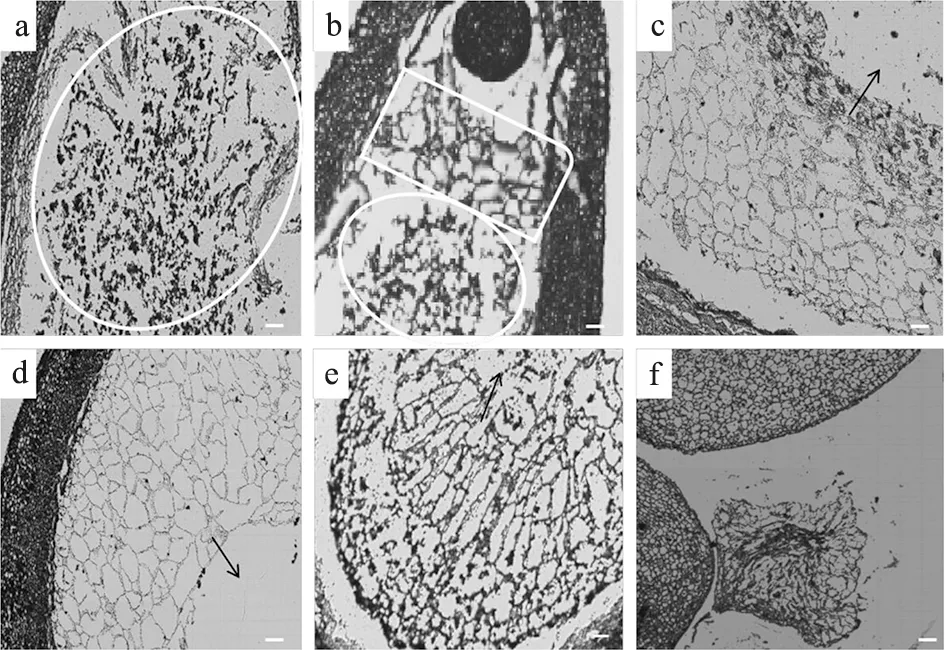
a. 盛花期后15 d的游离核胚乳,白色圆圈内是胚乳游离核;b. 盛花期后20 d后始细胞化后的胚乳,白色圆圈内是胚乳游离核,白色方框内是珠孔端细胞化后的胚乳;c. 盛花期后25 d的胚乳;d. 盛花期后30 d的胚乳;e. 盛花期后35 d的胚乳;f. 盛花期后45 d的胚乳;箭头指向胚乳腔,Bar=50 μm图3 胚乳的发育过程 a. The free nuclear endosperm of 15 d after anthesis, the free nuclear endospermisenclosed in white circle;b. Cell wall began to form in the endosperm of 20 d after anthesis, the free nuclear endosperm isenclosed in white circle, and endosperm cells of the micropylar end are enclosed in white box;c. Endosperm cells of 25 d after anthesis;d. Endosperm cells of 30 d after anthesis;e. Endosperm cells of 35 d after anthesis;f. Endosperm cells of 45 d after anthesis;The arrow points to endosperm chamber, Bar = 50 μmFig.3 The process of chestnut endosperm development
2.4 不同发育时期胚乳细胞的形态变化
为了进一步探究胚乳细胞的发育变化,对不同时期胚乳细胞的形态进行了测量,包括细胞的横径和纵径。结果表明,在盛花期后20 d左右,球形胚时期,游离核状态的胚乳正在从珠孔端向合点端进行细胞化,此时胚乳细胞横径和纵径相差不大,径值均较小(图4-a,f)。伴随着胚胎的发育,胚乳细胞开始扩张,淀粉粒增多,细胞化的胚乳进入成熟期,横径和纵径均有缓慢增加,但两者仍相差不大(图4-b,c,f)。在盛花期后35 d左右,胚胎发育至子叶胚时,胚乳细胞出现明显的横向扩张趋势,细胞的横径和纵径存在明显的差异(图4-d,g)。盛花期后40 d左右,随着子叶胚的膨大,胚乳细胞继续横向扩张,且纵径值大幅度降低(图4-e,g),在为胚胎提供营养的同时,更有利于子叶胚的膨大。
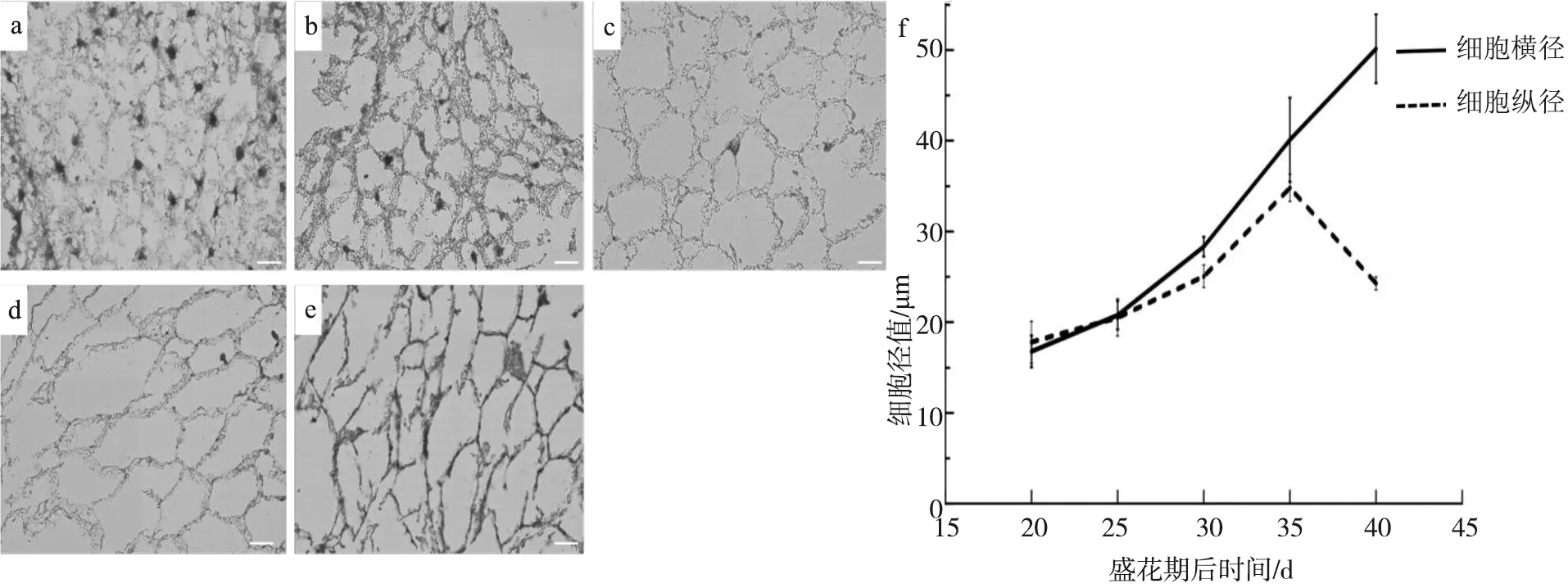
a.盛花期后20 d的胚乳细胞;b.盛花期后25 d的胚乳细胞;c. 盛花期后30 d的胚乳细胞;d. 盛花期后35 d的胚乳细胞;e. 盛花期后40 d的胚乳细胞;f. 细胞化的胚乳不同发育时期的细胞径值变化;Bar=20 μm图4 胚乳细胞的形态观察及径值变化a. Endosperm cells of 20 d after anthesis;b. Endosperm cells of 25 d after anthesis;c. Endosperm cells of 30 d after anthesis;d. Endosperm cells of 35 d after anthesis;e. Endosperm cells of 40 d after anthesis; f. Diameters of endosperm cells;Bar=20 μmFig.4 Morphological observation and diameter changes of endosperm cells
3 讨 论
在被子植物中,一对精细胞分别与卵细胞和中央细胞结合,在胚囊内完成双受精作用形成合子和初生胚乳核,两者分别以不同的方式进行分裂和分化[18, 19]。合子第一次分裂形成二细胞原胚,而后经过四细胞原胚、八细胞原胚、三十二细胞原胚[20, 21],随后依次经过球形胚、心形胚、鱼雷形胚及子叶形胚,最终发育为成熟的胚胎[22]。而初生胚乳核的分裂早于合子分裂,第一次分裂形成两个游离核,且核型胚乳以后进行的多次核分裂都不伴有细胞壁的形成[23, 24]。对于板栗而言,胚胎作为成熟果实的主要成分,它的发育与膨大是板栗果实体积和重量增加的主要原因[14,16]。与很多植物不同的是,成熟的板栗种子中并没有胚乳的存在。在本研究中通过对球形胚、心形胚、鱼雷形胚、子叶形胚的观察,确定了盛花期后胚胎发育的关键时间节点;同时观察了胚乳的形成、发育到解体的过程。依照Olsen[25]和旷仁平等人[26]对核型胚乳发育过程的时期分类,在本研究中观察到了胚乳发育的4个关键时期:游离核期、细胞化期、分化期和死亡期,进一步证实了板栗胚乳具备核型胚乳的典型特征。将胚胎与胚乳的发育相结合,研究结果表明:在球形胚时期(盛花期后15~20 d),胚胎细胞快速进行生命活动,处于高强度的分化及分裂状态,维管组织、表皮等结构开始形成[27, 28]。与此同步的是胚乳游离核向细胞化的转变(时期的转变:游离核期向细胞化期),且细胞化具有方向性,沿胚囊壁从珠孔端向合点端进行,其细胞与周围的胚胎细胞、珠被细胞也有明显的不同。在心形胚和鱼雷形胚时期(盛花期后20~30 d),球形胚顶端的两侧细胞加速分裂形成子叶原基,游离核胚乳完全细胞化,全部形成细胞;淀粉粒作为主要的营养储藏物质,数量明显增多。胚乳细胞迅速扩张,横纵径值均增加,体积增大;为了让胚胎更好的发育,胚乳腔形成(胚乳分化期)。子叶胚及子叶胚膨大时期(盛花期后35 d),胚乳细胞横向扩张明显,细胞横径值持续增加,但纵径值减小,两者相差较大,并伴随着胚胎的膨大开始逐渐被水解,此时期有逐渐被胚消化的胚乳细胞残余物的存在(胚乳死亡期)。有研究表明,有的植物胚胎在早期胚胎发育过程中,胚乳并不是主要的营养物质来源[29]。从本试验的研究结果可以表明,板栗胚的发育滞后于胚乳的发育,且在胚胎发育过程中时,胚乳细胞富含营养物质,到达成熟胚时,胚乳仅剩细胞残余物(即死亡期)。因此可以推断板栗胚胎发育所需的营养主要依赖于胚乳细胞所提供的营养物质。
板栗的种子存在一定的空瘪率,造成种子空瘪的原因较为复杂,在板栗胚胎和胚乳的发育过程中,所观察到的形成环形结构的形态畸形的胚胎,其胚乳细胞并未发生明显的横化,也并未观察到胚乳腔的存在,畸形胚胎是否能正常从胚乳细胞吸收营养物质还有待研究,但很可能是造成种子空瘪的一个重要原因[30-32]。而对于存在的2枚胚胎的情况,推测是由于在板栗的12~18枚胚珠中有2枚胚珠完成了双受精作用,因这2枚胚珠在同一心室,珠柄端相隔较近,导致2枚胚珠的珠柄粘合,继而可能发育成为常见的“双胞胎”。
本研究通过对胚胎发生及胚乳发育的过程的研究,揭示了两者在发育过程中的动态联系,并通过畸形胚胎对板栗种子败育的影响作了相关探讨,进一步掌握了板栗的生殖发育特征。
猜你喜欢
气象水文海洋仪器(2022年2期)2022-07-08
农产品市场周刊(2022年4期)2022-05-21
农产品市场周刊(2022年4期)2022-03-23
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
湖北农业科学(2017年6期)2017-04-26
小学阅读指南·高年级版(2016年9期)2016-10-31
江苏农业科学(2015年11期)2016-01-27
长江蔬菜·学术版(2014年7期)2015-01-21
中学生数理化·高二版(2008年11期)2008-06-17
