
基于gE蛋白的羊伪狂犬病病毒间接ELISA的建立与应用
2024-03-18 07:09刘广阔吴发兴于皓同王凯茸张锐铮许信刚
动物医学进展 2024年3期
刘广阔,吴发兴,于皓同,王凯茸,张锐铮,张 琪*,许信刚*
(1.西北农林科技大学动物医学院,陕西杨凌 712100;2.中国动物卫生与流行病学中心 国家动物血清库,山东青岛 266114)
伪狂犬病(Pseudorabies,PR)又称为奥叶基氏病(Aujeszky disease,AD),是由伪狂犬病病毒(Pseudorabies virus,PRV)引起的具有传染性的病毒性疾病。自该病毒报道以来[1],距今已有210年的历史。虽然猪是此病毒的唯一自然宿主,但其在人与其他哺乳动物中仍具有广泛的宿主,如山羊感染PRV后常以死亡告终[2],给养羊场(户)造成很大损失。
近年已有血清学技术及分子生物学方法用于伪狂犬病的诊断,但大部分血清学方法无法用于羊伪狂犬病的诊断,而病毒分离和分子生物学方法中的PCR、qPCR技术所需要的试验条件在中小型养殖场难以达成。因此,开发羊伪狂犬病诊断的血清学方法非常必要。近年采用的血清学检测方法中[3-9],间接ELISA方法因操作简单,所依赖的试验条件较低,并且具有较高的特异性和敏感性,在病原及抗体检测中应用广泛。本研究在克隆表达羊PRV gE蛋白的基础上,获得重组蛋白包被酶标板,建立检测羊源PRV抗体的间接ELISA检测方法(gE indirect ELISA,gE-iELISA),国内部分羊场用gE基因缺失疫苗对羊群进行免疫,该方法可以区分野毒感染和免疫接种动物,gE-iELISA方法填补了羊伪狂犬病血清学检测方法的空白,为规模化羊场提供了一种可供选择的检测方法。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒 羊伪狂犬病病毒(SDPD-14)由中国动物卫生与流行病学中心国家动物血清库(以下简称血清库)鉴定、分离与保存。pET-32a(+)载体购于博迈德基因技术有限公司,DH5α感受态细胞、BL21(DE3)细胞购于天根生物科技有限公司。
1.1.2 血清样品 羊伪狂犬病病毒标准阳性血清、羊伪狂犬病病毒抗体阴性血清、羊口疮病毒阳性血清、羊痘病毒阳性血清由中国动物卫生与流行病学中心国家动物血清库鉴定并保存;羊口蹄疫A型及O型病毒抗体阳性血清为北京金诺百泰生物技术有限公司试剂盒中的标准品,检测中使用的376份山羊临床血清样品采自云南、陕西、山东部分地区山羊场。
1.1.3 主要试剂 DNA/RNA提取试剂盒,西安天隆科技有限公司产品;琼脂糖凝胶DNA回收试剂盒、质粒快速小提试剂盒、T4 DNA连接酶,天根生物科技有限公司产品;PrimeSTAR®HS、BamHⅠ和XhoⅠ内切酶,Takara公司产品;TMB显色液,北京索莱宝科技有限公司产品;Ni-NTA His Bind Resin镍柱,七海复泰生物科技有限公司产品。
1.1.4 主要仪器 NP968核酸提取仪、基因扩增热循环仪,西安天隆科技有限公司产品;OSE-470P TGel蓝光凝胶成像仪,北京天根生物科技有限公司产品;精骐恒温振荡器,捷美电子有限公司产品;Flash Max 850型光吸收酶标仪,闪谱生物科技有限公司产品。
1.2 方法
1.2.1 引物设计 参考此前本实验室对SDPD-14株的测序结果(KY398797.1),用Primer 5.0进行特异性引物的设计(表1)。
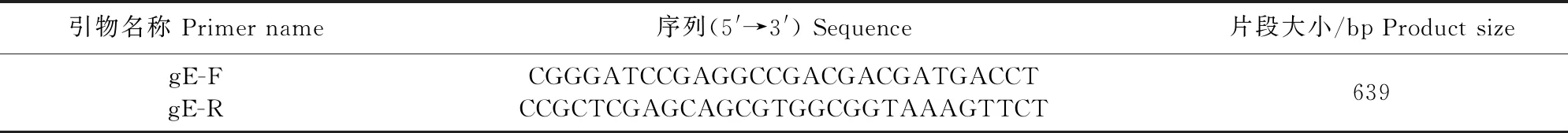
表1 gE基因引物序列
1.2.2 羊PRVgE基因扩增与重组表达载体构建 用DNA/RNA提取试剂盒与NP968核酸提取仪提取PRV SDPD-14株的基因组,以得到的DNA为模板,用1.2.1中的特异性引物进行gE基因扩增。PCR反应体系为:PrimeSTAR HS 10 μL,gE-F/gE-R各1 μL,DNA模板2 μL,用RNase-free ddH2O补足至20 μL。PCR反应程序为:95℃ 5 min;95℃ 30 s,58℃ 45 s,72℃ 1 min,30次循环;72℃延伸10 min。反应结束后经电泳判定是否得到目的条带。胶回收目的片段后,经BamHⅠ和XhoⅠ双酶切并与相同酶双酶切的pET-32a(+)载体连接构建重组质粒。对重组质粒分别进行双酶切鉴定和测序鉴定,将鉴定正确的重组质粒命名为pET-32a-gE。
1.2.3 重组gE蛋白诱导表达条件的优化 取1.2.2中的pET-32a-gE重组质粒10 μL转化至BL21(DE3)感受态细胞中,通过菌液PCR筛选出阳性表达菌株,表达前先将菌株活化,按照1∶100的比例转接于含有氨苄青霉素的液体LB培养基中,置于摇床37℃、220 r/min培养120 min测定其OD600nm值,当OD600nm值为0.6~0.8时,加入不同浓度的IPTG(使IPTG的最终浓度分别为0.2、0.4、0.6、0.8、1.0 mmol/L)对表达菌株进行诱导,37℃继续培养8 h。离心收集菌体沉淀,处理后经SDS-PAGE凝胶电泳以确定最佳诱导浓度。以最佳诱导浓度在37℃下对菌液进行诱导,在2~8 h内每间隔1 h离心后收集沉淀。通过SDS-PAGE凝胶电泳结果确定最佳诱导时间。
1.2.4 重组gE蛋白的表达形式的确定及纯化 参照1.2.3结果,大量诱导表达重组蛋白,诱导结束后离心并收集沉淀。将沉淀重悬后进行超声破碎,分别将离心得到的上清液、超声裂解后的上清液与沉淀各取30 μL,用2×蛋白上样缓冲液进行制样,通过SDS-PAGE凝胶电泳结果确定目的蛋白所在区间以明确重组gE蛋白的表达形式。用镍柱对重组gE蛋白进行纯化,用不同浓度尿素透析复性重组蛋白,用BCA蛋白定量方法测定重组蛋白浓度。
1.2.5 重组gE蛋白的Western blot分析 将1.2.4中复性得到的重组gE蛋白制样后进行SDS-PAGE,将目标区域切下,在半干转膜仪上进行转膜。将膜取下后用5%脱脂奶粉液封闭2 h,随后将其置1∶200稀释的羊伪狂犬病毒阳性血清中在4℃孵育12 h,次日洗膜后用1∶10 000稀释的HRP标记兔抗山羊IgG在室温下孵育1.5 h,洗膜后滴加ECL发光液至膜上,弃去多余液体后进行曝光。
1.2.6 gE-iELISA检测方法的建立、优化与应用
1.2.6.1 抗原包被量、一抗及酶标二抗稀释度的确定 用三因素正交试验确定最优条件,分别用1、2、4、8 μg/mL的gE蛋白包被酶标板;羊伪狂犬病毒阳性及阴性血清分别采用为1∶50、1∶100、1∶200稀释度;HRP标记兔抗山羊IgG的稀释比例分别为1∶2 500、1∶5 000、1∶10 000,进行间接ELISA,操作步骤参照文献[10],根据测定结果计算P/N,将P/N最大组合确定为抗原包被量、血清和酶标抗体的最佳工作条件。
1.2.6.2 其他试验条件的优化 在1.2.6.1中确定的最佳工作条件下,37℃分别以60、90、120 min的孵育时间对脱脂奶粉封闭时间、一抗及二抗孵育时间进行优化;以15、30、45 min进行TMB显色时间的优化,将P/N最大组确定为最适封闭时间、孵育时间和显色时间。
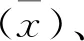
1.2.6.4 特异性试验 在1.2.6.1及1.2.6.2中确定的最适条件下,对羊痘病毒阳性血清、羊口蹄疫病毒A型及O型阳性血清、羊口疮病毒阳性血清进行检测,同时以羊伪狂犬病病毒阳性血清为阳性对照,用实验室保存的SPF山羊阴性血清作为对照,评价建立方法的特异性。
1.2.6.5 灵敏性试验 对羊伪狂犬病病毒阳性血清分别进行1∶32、1∶64、1∶128、1∶256、1∶512、1∶1 024、1∶2 048、1∶4 096的稀释,同时设置阴、阳性对照,用本研究建立的方法进行灵敏性试验。
1.2.6.6 重复性试验 用同批次表达纯化复性的重组gE蛋白包被酶标板,对3组不同的羊伪狂犬病病毒阴、阳性血清重复检测3次,根据结果评价批内重复性;用3组不同批次纯化复性重组gE蛋白包被酶标板,分别对3组不同的羊伪狂犬病毒阴、阳性血清重复检测3次,根据结果评价批间重复性。
1.2.6.7 临床样本检测 以本研究建立的gE-iELISA方法,对血清库保存的云南、陕西、山东部分地区羊场的376份山羊血清进行检测,计算羊伪狂犬病病毒的阳性率,并对此检测方法的临床应用效果做出评价。
2 结果
2.1 gE基因的扩增及重组质粒构建
以PRV SDPD-14株的DNA为模板,通过PCR扩增gE基因,得到639 bp的基因片段(图1),其大小与预期试验结果一致。对构建得到的pET-32a-gE重组质粒使用BamHⅠ和XhoⅠ双酶切后,得到639 bp的目的条带与约5 000 bp的载体条带(图2),结果与预期一致,说明重组表达载体pET-32a-gE构建成功。
M.DNA标准DL 2 000;1.gE基因扩增产物;2.阴性对照

M.DNA标准DL 5 000;1.重组载体pET-32a-gE经BamHⅠ和XhoⅠ双酶切

M.蛋白分子质量标准;1.诱导的pET-32a空载体;2~6.37℃下0.2,0.4,0.6,0.8,1.0 mmol/L的IPTG终浓度条件下诱导8 h的重组pET-32a-gE;7.重组pET-32a-gE未诱导

M.蛋白分子质量标准;1.诱导pET-32a空载体;2-8.pET-32a-gE在BL21中诱导2、3、4、5、6、7、8 h的表达产物

M.蛋白分子质量标准;1.菌液上清;2.菌体超声裂解液上清;3.菌体超声裂解沉淀;4.纯化gE蛋白
2.2 重组gE蛋白表达条件优化
以不同浓度的IPTG诱导重组菌株表达目的蛋白。结果显示,当IPTG终浓度为0.6 mmol/L时,目的蛋白达到最高产量(图3);37℃、0.6 mmol/L的IPTG终浓度下进行诱导表达,并在不同的时间收集菌体,结果显示该目的蛋白于8 h时达到最高的表达量(图4)。
2.3 重组gE蛋白的表达及纯化
在上述最适条件下诱导表达重组蛋白,收集菌体处理后进行SDS-PAGE。结果显示,重组gE蛋白主要在超声裂解后沉淀中以包涵体形式大量存在,用镍柱纯化得到目的蛋白大小为42 ku(图5)。
2.4 重组gE蛋白的Western blot鉴定
用羊PRV阳性血清作为一抗对纯化后的重组gE蛋白进行Western blot分析,发现在42 ku处出现一条清晰的特异性条带。表明重组gE蛋白可以被羊PRV阳性血清特异性识别,即具有良好的反应原性(图6)。

M.蛋白分子质量标准;1.重组gE蛋白;2.阴性对照
2.5 最佳工作条件确定
经过三因素正交试验可知,当重组gE蛋白的抗原包被浓度为1 μg/mL,血清以1∶100比例稀释,酶标二抗以1∶10 000比例稀释时P/N最高,将其确定为最佳工作条件(表2)。
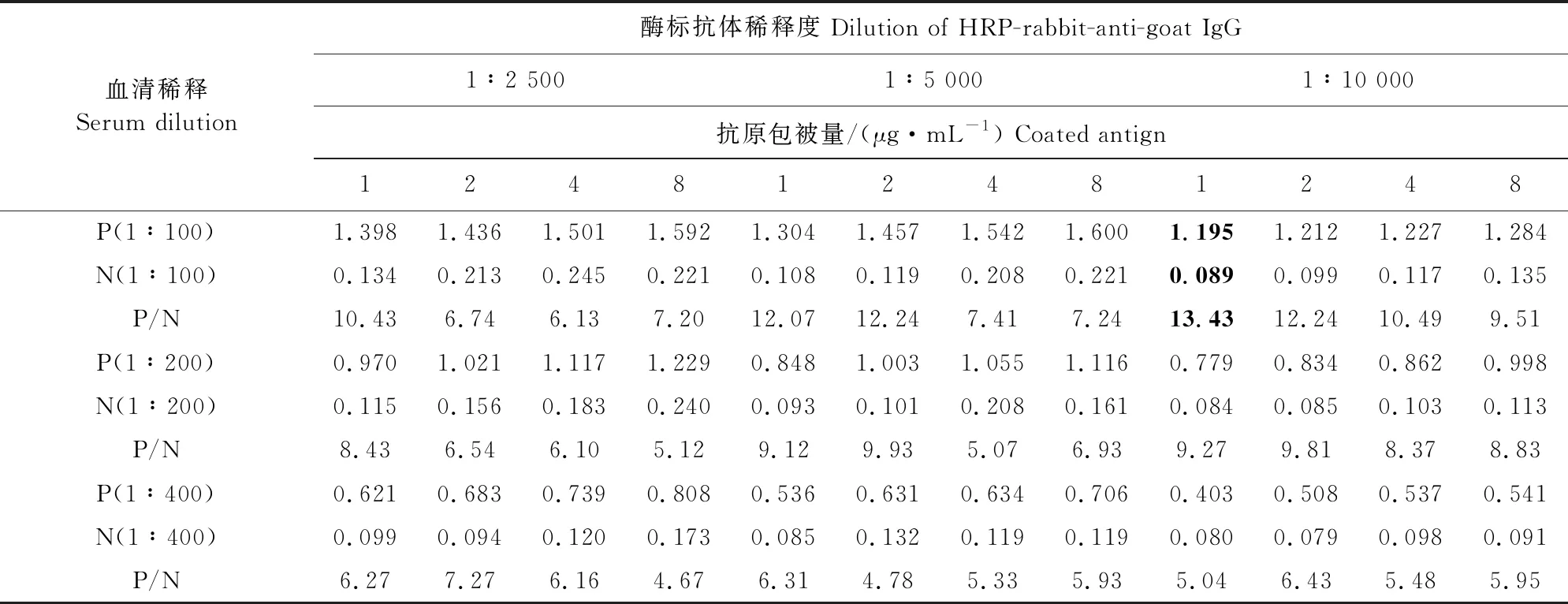
表2 抗原包被浓度、血清及酶标抗体稀释比例的确定
2.6 其他优化条件结果
在确定的最佳工作条件下进行试验,在37℃下进行其他条件的优化,结果显示封闭效果最优为5%脱脂奶粉封闭60 min,样品最佳孵育时间为60 min,HRP标记兔抗山羊酶标抗体最适孵育时间为60 min,TMB最佳显色时间为30 min。
2.7 阴阳性临界值的确定

2.8 特异性试验结果
用确定的最适条件,对羊痘病毒、羊口蹄疫A型及O型病毒、羊口疮病毒阳性血清进行检测,并以羊伪狂犬病毒抗体阳性血清为阳性对照,羊伪狂犬病毒抗体阴性血清为阴性对照。结果显示,阴阳性对照成立,其余血清OD450 nm值均小于0.284,为PRV抗体阴性,证明所建立的PRV gE-iELISA方法具有良好的特异性。
2.9 灵敏性试验结果
对羊伪狂犬病毒阳性血清分别以1∶32、1∶64、1∶128、1∶256、1∶512、1∶1 024、1∶2 048、1∶4 096比例稀释,同时设置阴阳性对照进行检测。结果显示,在阴阳性对照成立的前提下,当羊伪狂犬病毒阳性血清稀释至1∶2 048时,OD450 nm值为0.293,仍大于临界值,表明建立的方法具有较好的灵敏性。
2.10 重复性试验结果
用确定的最适条件,分别进行批内试验(变异系数为2.45%~5.00%)和批间试验(变异系数为2.55%~7.41%),变异系数均不超过10%,表明此方法重复性良好。
2.11 临床样本检测结果
用本试验建立的方法,对采自云南、陕西、山东部分地区羊场的376份山羊血清样本进行检测,其中40份血清呈羊伪狂犬病毒阳性,阳性率为10.6%。
3 讨论
2011年新出现的PRV变异株[11],使PR在Bartha-K61株弱毒疫苗免疫过的动物中重新暴发[12],PRV变异株对易感动物和人类的毒力比经典株更强[13],本研究建立的gE-iELISA方法对于目前缺乏相应检测手段的小型羊场有重要意义。伪狂犬病毒gE蛋白的主要抗原区集中在52-238位氨基酸[14]。本研究以此为依据,设计构建原核表达重组载体的引物,对gE蛋白的主要抗原区域进行表达,以保证其在用于包被抗原时具有较高的准确性。用镍柱对表达的重组gE蛋白进行纯化,并进行Western blot试验,结果表明重组gE蛋白具有良好的反应原性,可以作为诊断抗原用于酶标板的包被。
此前已在病死山羊的脑组织及肺脏组织中分离到伪狂犬病毒[15],虽然接种了伪狂犬病病毒活疫苗(Bartha-K61),但疫苗的安全性无法得到保障,有关于注射疫苗后导致羊死亡的报道[18],说明该病在羊群中的防控仍以病毒的及时检出为主。人也可感染伪狂犬病病毒,可引起神经症状,如眼内炎和脑炎,人类对本病的感染多是由于体表存在伤口。
本研究建立的gE-iELISA方法对云南、陕西、山东部分羊场收集到的376份山羊血清检测,PRV阳性率为10.6%,此数值比2008年青海省的4.65%有所升高[16],说明伪狂犬病病毒在羊群中的感染率呈上升趋势,应予以足够重视。
猜你喜欢
数学物理学报(2021年4期)2021-08-30
新世纪智能(数学备考)(2020年10期)2021-01-04
动漫星空(兴趣百科)(2020年3期)2020-03-24
英美文学研究论丛(2018年2期)2018-08-27
中成药(2017年12期)2018-01-19
中国交通信息化(2017年8期)2017-06-06
老年医学与保健(2017年6期)2017-02-06
湖南畜牧兽医(2016年3期)2016-06-05
兽医导刊(2016年12期)2016-05-17
中国继续医学教育(2015年1期)2016-01-06
